2 тарау
Жердегі өмірдің әртүрлілігі
2.1. Классификация
2.1.1. Ол не үшін керек?
Сізді кішкентай балалар өзінің іс – әрекеттерімен таңқалдырғаны рас шығар, мысалы әртүсті мұзды кәмпиттер немесе маркаларды сұрыптау, ол жинаған футболға билеттер немесе өзге де нәрселерге, онда сіз, сезікті іс – әрекеттердің куәгерлерінің бірі болдыңыз – кішкене сөрелерді жайып қою тілегі. Міне, бұл жерде классификация бар. Классификация – құрылымы бойынша ұқсас заттарды бір топқа біріктіру. Таксономия –классификацияның қағидасы, әдісі және ережесі бойынша тағайындалған ғылымның бөлігі. Біз неге классификациялауға ынтызар болып тұрамыз? Бұл сұрақтың жауабына кейбір биологтар былай жауап береді: біз өмір сүруді жеңілдету үшін біз бәрін топтастырып қоямыз. Біздің санамыз үлкен көлемде қоздырғыштарды жинақтап қалғанда, біз оларды жаймен жүйелеп, оларды классификациялаймыз. Біздің классификациялауда жасйтын алғашқы қадамдарымыз қате болып та жатады, мысалы, кішкентай бала кейде кез келген 4 аяқтыларды ит деп атап қалуы мүмкін. Бірақ жаймен адамда қоршаған ортаны тану туралы бір жүйе қалыптасады.
Жер шарында жартылай миллиондаған тірі ағзалардың түрі табылды, бірақ, өткізілген бағаға сәйкес бұның саны 10 – 100млнға жету керек. Және бұл ағзаларды топтастыру да таңқаларлық жай емес екені белгілі. Бұл классификациялар бір – бірінен олардың қандай мақсатпен жаратылатыны туралы ерекшеленеді. Мысалы, ерте заманғы Қытайда жануарлар патшалығы топтар тізбегіне бөлінді, қазіргі күнгі біздің тілімізбен жұмсағырақ айтқанда, өте біртүрлі көрінеді. бұл, соның ішінде, мификалық жануарлар, панасыз жануарлар, гүл сауытын сындыратын хайуандар немесе алыстан еске алатын шыбындар. Топтастыруды мысал ретінде былай жасаған дұрыс, мысалы, өсімдіктерді : улы және жеуге жарамды немесе жануарларды ұшатын немесе ұшпайтын. Заманауи топтастыруда , өзіміз көріп жүргеніміздей, әдетте ағзалардың арасындағы эволюциялық байланыстарды ерекше акцентепен қарастырады.
Біздің жануарларымызға деген көзқарасымыз кең болған сайын , классификация да өзгеріске түседі. Бірақ, бір затты да ұмытпау керек, мейлі бір ғана тірі ағзаның классификациялық жүйесі болсын, ол қанша біздің қалауымызбен болса да, ешқашан шүбәсіз, шын бола алмайды.
2.1.2. Таксономия
Таксоносия 2 бутаққа бөлшектенеді: 1 – ші бұтақ – ағзаның атауы бойынша иеленуі – бұл номенклатура деп аталса, екіншісі – ағзаның топ бойынша бөлінуі, бұл – систематика деп аталады. Систематикада ағзалардың ұқсастығымен қатар айырмашылылғы да жатады. Биологиялық номенклатураны шведтің табиғат зерттеушісі Карл Линней (1707 –1778ж) ашқан, биологиялық номенклатура биномальды жүйеге негізделген. Биномальды жүйесінде әрбір ағзаға екі – екіден латындық атау беріледі: рулық және түрлік. Рулық атаулар бас әріппен жазылады, ал түрлік атаулар кіші әріппен жазылады. Мысалы, адам мынадай атқа ие делік: Homo sapiens; бұл жерде Homo рулық атау және sapiens – түрлік атау. Латындық ру және түр атаулары жазба әріп тәрізді көлбеу қаріппен жазылады. Және оларды кәдімгі қаріппен де жаза беруге болады, бірақ астын қара сызықпен сызып қою керек. Осы айтылғандар туралы латындық атауларды жазып жатқанда ұмытпауымыз керек. Рулық атауларды қысқартып жазуға да болады, мысалы: H. sapiens , бірақ бұны осылай оның алдында толық атауы қолданғаннан кейін жазу керек. Ағзалардың латындық әріптері бүкін Дүниежүзінде қабылданған. Бұл жалпы бірге қабылданған жергілікті мекеннің тіршілігінің атауын айтқанда , шатастырмау үшін мүмкіндік береді. Сондай – ақ, Ficus caria өсімдігі көп тараған бірнеше атауы бар сөз: інжір (инжир), інжір ағашы (фиговое дерево), інжір ағашы (смоковница), інжір ағашы (фига). Uncia uncia деген қар барысын ілбіс (ирбис) деп атаса, ал қар қойын Ovis canadensis 2 атаумен атайды: шылым түтігі (чубук) және толсторог. Белгілі бір атау 1 түрден көп сөзге түсіндірілсе, үлкен емес шатастырылу болады. Мысалы, зимовник (әлекшөп), оны безвременник (Colchicum, әлекшөп) деп мелантийлік тұқымдасқа байланысты айтылса, морозник (Helleborus, әлекшөп) сарғалдақтар тұқымдасына жатады.
2.1.3. Таксономиялық иерархия
Линней өмірінің соңғы күндері рулық және түрлік атауларлдан басқа да көптеген заттар қосып, биномальды жүйені күшейтіп кетті. Ол иерархия тобын құрды, ең ірі көлемдегі топты және иерархияда бірінші тұратынды –патшалық деп санады. Бұл істелінген жұмыс осы күні де қолданылады.
2.1.1. Ол не үшін керек?
Сізді кішкентай балалар өзінің іс – әрекеттерімен таңқалдырғаны рас шығар, мысалы әртүсті мұзды кәмпиттер немесе маркаларды сұрыптау, ол жинаған футболға билеттер немесе өзге де нәрселерге, онда сіз, сезікті іс – әрекеттердің куәгерлерінің бірі болдыңыз – кішкене сөрелерді жайып қою тілегі. Міне, бұл жерде классификация бар. Классификация – құрылымы бойынша ұқсас заттарды бір топқа біріктіру. Таксономия –классификацияның қағидасы, әдісі және ережесі бойынша тағайындалған ғылымның бөлігі. Біз неге классификациялауға ынтызар болып тұрамыз? Бұл сұрақтың жауабына кейбір биологтар былай жауап береді: біз өмір сүруді жеңілдету үшін біз бәрін топтастырып қоямыз. Біздің санамыз үлкен көлемде қоздырғыштарды жинақтап қалғанда, біз оларды жаймен жүйелеп, оларды классификациялаймыз. Біздің классификациялауда жасйтын алғашқы қадамдарымыз қате болып та жатады, мысалы, кішкентай бала кейде кез келген 4 аяқтыларды ит деп атап қалуы мүмкін. Бірақ жаймен адамда қоршаған ортаны тану туралы бір жүйе қалыптасады.
Жер шарында жартылай миллиондаған тірі ағзалардың түрі табылды, бірақ, өткізілген бағаға сәйкес бұның саны 10 – 100млнға жету керек. Және бұл ағзаларды топтастыру да таңқаларлық жай емес екені белгілі. Бұл классификациялар бір – бірінен олардың қандай мақсатпен жаратылатыны туралы ерекшеленеді. Мысалы, ерте заманғы Қытайда жануарлар патшалығы топтар тізбегіне бөлінді, қазіргі күнгі біздің тілімізбен жұмсағырақ айтқанда, өте біртүрлі көрінеді. бұл, соның ішінде, мификалық жануарлар, панасыз жануарлар, гүл сауытын сындыратын хайуандар немесе алыстан еске алатын шыбындар. Топтастыруды мысал ретінде былай жасаған дұрыс, мысалы, өсімдіктерді : улы және жеуге жарамды немесе жануарларды ұшатын немесе ұшпайтын. Заманауи топтастыруда , өзіміз көріп жүргеніміздей, әдетте ағзалардың арасындағы эволюциялық байланыстарды ерекше акцентепен қарастырады.
Біздің жануарларымызға деген көзқарасымыз кең болған сайын , классификация да өзгеріске түседі. Бірақ, бір затты да ұмытпау керек, мейлі бір ғана тірі ағзаның классификациялық жүйесі болсын, ол қанша біздің қалауымызбен болса да, ешқашан шүбәсіз, шын бола алмайды.
2.1.2. Таксономия
Таксоносия 2 бутаққа бөлшектенеді: 1 – ші бұтақ – ағзаның атауы бойынша иеленуі – бұл номенклатура деп аталса, екіншісі – ағзаның топ бойынша бөлінуі, бұл – систематика деп аталады. Систематикада ағзалардың ұқсастығымен қатар айырмашылылғы да жатады. Биологиялық номенклатураны шведтің табиғат зерттеушісі Карл Линней (1707 –1778ж) ашқан, биологиялық номенклатура биномальды жүйеге негізделген. Биномальды жүйесінде әрбір ағзаға екі – екіден латындық атау беріледі: рулық және түрлік. Рулық атаулар бас әріппен жазылады, ал түрлік атаулар кіші әріппен жазылады. Мысалы, адам мынадай атқа ие делік: Homo sapiens; бұл жерде Homo рулық атау және sapiens – түрлік атау. Латындық ру және түр атаулары жазба әріп тәрізді көлбеу қаріппен жазылады. Және оларды кәдімгі қаріппен де жаза беруге болады, бірақ астын қара сызықпен сызып қою керек. Осы айтылғандар туралы латындық атауларды жазып жатқанда ұмытпауымыз керек. Рулық атауларды қысқартып жазуға да болады, мысалы: H. sapiens , бірақ бұны осылай оның алдында толық атауы қолданғаннан кейін жазу керек. Ағзалардың латындық әріптері бүкін Дүниежүзінде қабылданған. Бұл жалпы бірге қабылданған жергілікті мекеннің тіршілігінің атауын айтқанда , шатастырмау үшін мүмкіндік береді. Сондай – ақ, Ficus caria өсімдігі көп тараған бірнеше атауы бар сөз: інжір (инжир), інжір ағашы (фиговое дерево), інжір ағашы (смоковница), інжір ағашы (фига). Uncia uncia деген қар барысын ілбіс (ирбис) деп атаса, ал қар қойын Ovis canadensis 2 атаумен атайды: шылым түтігі (чубук) және толсторог. Белгілі бір атау 1 түрден көп сөзге түсіндірілсе, үлкен емес шатастырылу болады. Мысалы, зимовник (әлекшөп), оны безвременник (Colchicum, әлекшөп) деп мелантийлік тұқымдасқа байланысты айтылса, морозник (Helleborus, әлекшөп) сарғалдақтар тұқымдасына жатады.
2.1.3. Таксономиялық иерархия
Линней өмірінің соңғы күндері рулық және түрлік атауларлдан басқа да көптеген заттар қосып, биномальды жүйені күшейтіп кетті. Ол иерархия тобын құрды, ең ірі көлемдегі топты және иерархияда бірінші тұратынды –патшалық деп санады. Бұл істелінген жұмыс осы күні де қолданылады.
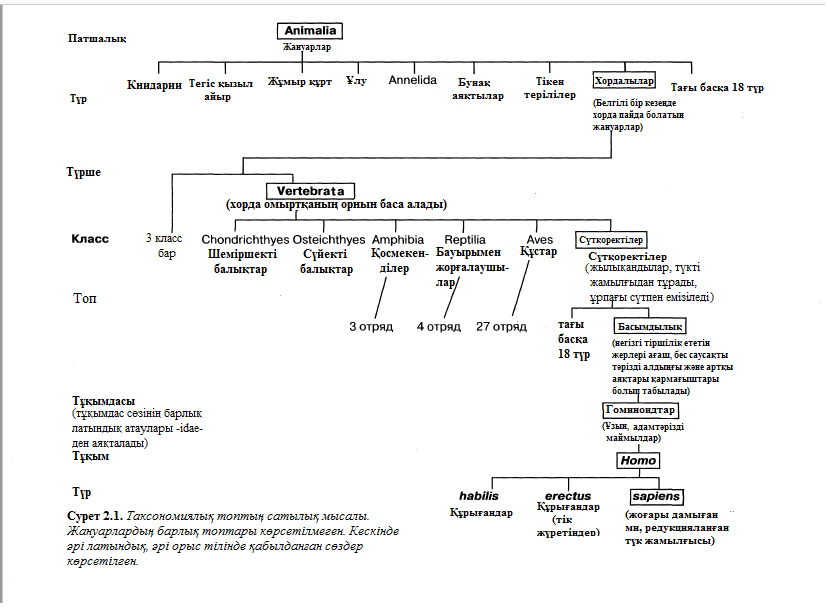
Оның қатарына келесі иерархиялық бірліктер кіреді:
Патшалық*
Тұқымдас (өсімдіктер бөлімі) ХІХ ғасырдың соңында Геккелдікімен жүргізілген
Класс*
Отряд*
Тұқымдас*
Тұқымдас*
Түр*
*Линнейдікімен жүргізілген
Классификация патшалығы туралы айқын мысалы 2.1. суретінде көрсетілген. Көріп тұрғанымыздай, суретте әрбір қатар немесе таксон, өзінің кезегінде басқа топтардың төмен дәрежедегі қатарын қосып алып тұр. Мысалы, Vertebrata (омыртқалылар) түршесі 6 классқа кіріп кетеді, ал Homo (адам) руы 3 түрден тұрады, оны 2 – і құрып кетті. Әрбір топтың өзіне тән белгілері болады, бұл топ өкілдері үшін өте сиректік тән болып табылады. Мұндай белгілер диагностикалықтар деп аталады. Мысалы, түк жамылғылары тек сүтқоректліреде кездеседі (Mammalia класы). Демек, түк жамылғысы – сүтқректілердің диагностикалық белгісі. Бірақ сүтқоректілер – құстар, бауырымен жорғалаушылар, қосмекенділер және балықтар секілді өткен иерархия тобындағы диагностикалық белгілерді иемдене алады, нақтырақ омыртқалылар.
2.1.4. Түрлер
Иерархияның барлық дәрежедегі нақты анықтамасы «түр» термині. Құнарлы ұрпақ бере алатын бір бірімен будандастырыла алатын жақын туысты ағзалардың тобын түр анықтап бере алады. Кейбір жағдайларда екі жақын туысты будандастырғанда ұрпақсыздыөөа әкеліп соқтырады. Мысалы, гибридті айтсақ болады, ол жылқымен (бие) есектің (аталық) шағылыстыруынан пайда болған ұрпақсыз болып қалды. Демек, есек пен жылқы әр түрге жатады.
Қашыр (мул, әңгі мен биенің буданы) ата – анасынан үлкен төзімділігімен ерекшеленеді, ата – анадан сау белгілері қамтамасыз етілген мұралану (қоспалы қуат).
Құнарлы ұрпаққа байланысты ережеден тыс ерекшеліктер бар. Мысалы, арыстандар мен тигрлер іртүрлі түрлерге жатады. Бірақ, егер арыстан мен тигрді (аталығын) шағылыстырсақ, онда құнарлы ұрпақ алынады, ал арыстан мен тигрдің аналық түрін шағылыстырсақ, онда ұрпақсыздыққа әкеліп соқтырады. Бірақ, тигрлердің тіршілігі табиғат жағдайымен байланысты, орманда тіршілік етеді, ал арыстандар шалғынды аймақта күнін көреді, екеуінің арасындағы шағылыстырылу еріксіздік болып тобылады.
Әрбір түр өзінің жеке морфологиялық, мінез – құлықтық және экологиялық белгілерін құрайды. Егер біз ойша таксономиялық иерархияның жоғарғы баспалдағына қарайтын болсақ, біз белгілердің саны бір топтан екінші топқа құлдырайтынын көреміз. Мысалы, бір рудың бірдей өкілдері ұқсас белгілерді тұқымдасқа немесе отрядқа қарағанда, көптеп құрайды.
Айтылғандарға қарағандай, түрлерге нақты анықтама беру практика жүзінде мүмкін емес. Уақыт өте берген сайын олар өзгеріске ұшырап отыра береді және бұл таңғаларлық жайт емес. Ең бейімделгіш даралар түрлерінің өзгеру үдерісі тіршілік етумен қамтамасыз етіледі, яғни ол даралар айқын қоршаған ортаға бейімделгіш даралар, бұл табиғи іріктеудің теориямен жарасымы болып табылады. Қоршаған ортада қандай да бір өзгеріс пайда болған жағдайда, іріктеу нәтижесінде және ақырындап көптеген ұрпақтар дамуына түр өзгерісін әкелетін дараларға жағдай туғызады. Бір түрдің бірақ әртүрлі популяция болған кезде бір – бірінен бөлек (жеке) тұрады, мысалы, экологиялық немесе физикалық бөгеттерде, сондай – ақ мұхит немесе таулы тізбектерде, бұл популяциялардың әрі қарайғы дамуы әр түрлі жолмен кетуі мүмкін және соңында олардың арасындағы шағылытырылу мүлдем мүмкін емес болады. Олар бөлек түр болып қалады.
Кейбір жағдайларда әртүрлі түрдің арасындағы шағылыстырылуда қатты генетикалық ұқсастық болмауы да мүмкін. Сондай – ақ, сыңқылдаған шағала мен тауық әр түрге жатады, сондықтан да олар морфологиялық және мінез – құлықтарымен ерекшеленеді дәне әдетте шағылыстырылмайды. Бірақ, кейбір жағдайларда олар бір жерге ұя салады және кейде тұқымдастық жұп болып саналады.
Патшалық*
Тұқымдас (өсімдіктер бөлімі) ХІХ ғасырдың соңында Геккелдікімен жүргізілген
Класс*
Отряд*
Тұқымдас*
Тұқымдас*
Түр*
*Линнейдікімен жүргізілген
Классификация патшалығы туралы айқын мысалы 2.1. суретінде көрсетілген. Көріп тұрғанымыздай, суретте әрбір қатар немесе таксон, өзінің кезегінде басқа топтардың төмен дәрежедегі қатарын қосып алып тұр. Мысалы, Vertebrata (омыртқалылар) түршесі 6 классқа кіріп кетеді, ал Homo (адам) руы 3 түрден тұрады, оны 2 – і құрып кетті. Әрбір топтың өзіне тән белгілері болады, бұл топ өкілдері үшін өте сиректік тән болып табылады. Мұндай белгілер диагностикалықтар деп аталады. Мысалы, түк жамылғылары тек сүтқоректліреде кездеседі (Mammalia класы). Демек, түк жамылғысы – сүтқректілердің диагностикалық белгісі. Бірақ сүтқоректілер – құстар, бауырымен жорғалаушылар, қосмекенділер және балықтар секілді өткен иерархия тобындағы диагностикалық белгілерді иемдене алады, нақтырақ омыртқалылар.
2.1.4. Түрлер
Иерархияның барлық дәрежедегі нақты анықтамасы «түр» термині. Құнарлы ұрпақ бере алатын бір бірімен будандастырыла алатын жақын туысты ағзалардың тобын түр анықтап бере алады. Кейбір жағдайларда екі жақын туысты будандастырғанда ұрпақсыздыөөа әкеліп соқтырады. Мысалы, гибридті айтсақ болады, ол жылқымен (бие) есектің (аталық) шағылыстыруынан пайда болған ұрпақсыз болып қалды. Демек, есек пен жылқы әр түрге жатады.
Қашыр (мул, әңгі мен биенің буданы) ата – анасынан үлкен төзімділігімен ерекшеленеді, ата – анадан сау белгілері қамтамасыз етілген мұралану (қоспалы қуат).
Құнарлы ұрпаққа байланысты ережеден тыс ерекшеліктер бар. Мысалы, арыстандар мен тигрлер іртүрлі түрлерге жатады. Бірақ, егер арыстан мен тигрді (аталығын) шағылыстырсақ, онда құнарлы ұрпақ алынады, ал арыстан мен тигрдің аналық түрін шағылыстырсақ, онда ұрпақсыздыққа әкеліп соқтырады. Бірақ, тигрлердің тіршілігі табиғат жағдайымен байланысты, орманда тіршілік етеді, ал арыстандар шалғынды аймақта күнін көреді, екеуінің арасындағы шағылыстырылу еріксіздік болып тобылады.
Әрбір түр өзінің жеке морфологиялық, мінез – құлықтық және экологиялық белгілерін құрайды. Егер біз ойша таксономиялық иерархияның жоғарғы баспалдағына қарайтын болсақ, біз белгілердің саны бір топтан екінші топқа құлдырайтынын көреміз. Мысалы, бір рудың бірдей өкілдері ұқсас белгілерді тұқымдасқа немесе отрядқа қарағанда, көптеп құрайды.
Айтылғандарға қарағандай, түрлерге нақты анықтама беру практика жүзінде мүмкін емес. Уақыт өте берген сайын олар өзгеріске ұшырап отыра береді және бұл таңғаларлық жайт емес. Ең бейімделгіш даралар түрлерінің өзгеру үдерісі тіршілік етумен қамтамасыз етіледі, яғни ол даралар айқын қоршаған ортаға бейімделгіш даралар, бұл табиғи іріктеудің теориямен жарасымы болып табылады. Қоршаған ортада қандай да бір өзгеріс пайда болған жағдайда, іріктеу нәтижесінде және ақырындап көптеген ұрпақтар дамуына түр өзгерісін әкелетін дараларға жағдай туғызады. Бір түрдің бірақ әртүрлі популяция болған кезде бір – бірінен бөлек (жеке) тұрады, мысалы, экологиялық немесе физикалық бөгеттерде, сондай – ақ мұхит немесе таулы тізбектерде, бұл популяциялардың әрі қарайғы дамуы әр түрлі жолмен кетуі мүмкін және соңында олардың арасындағы шағылытырылу мүлдем мүмкін емес болады. Олар бөлек түр болып қалады.
Кейбір жағдайларда әртүрлі түрдің арасындағы шағылыстырылуда қатты генетикалық ұқсастық болмауы да мүмкін. Сондай – ақ, сыңқылдаған шағала мен тауық әр түрге жатады, сондықтан да олар морфологиялық және мінез – құлықтарымен ерекшеленеді дәне әдетте шағылыстырылмайды. Бірақ, кейбір жағдайларда олар бір жерге ұя салады және кейде тұқымдастық жұп болып саналады.

2.1.5. Жасанды және табиғи классификация
Классификацияның екі түрі бар: жасанды және табиғи. Жасанды классификацияда негізге бір немесе бірнеше айырмалы белгілер алынады. Ол қолдануда ыңғайлылық және қарапайымдылық болса, практикалық тапсырмаларды қолданылады және жасап шығарылады. Жасанды классификация ерте ғасырлардағы Өытауда классификацияның бір жүйесі болып саналған. Линней барлық құрт тәрізді ағзаларды Vermes деген бір ғана топқа қосып, байланыстырды. Бұл топқа тіпті әртүрлі жануарлар да кірді, олар қарапайым жұмырдан(жұмыр құрт) және жауын құртынан, жыланға дейін барады. Линнейдің жасаған классификация тізбегі жасандыға да байланысты болып келеді, сондықтан да онда маңызды табиғи қарым – қатынас ескерілмеген, сондай – ақ, мысалы, жылан омыртқалы болса, ал жауын құртында ол жоқ. Шын мәнінде, жыландар құрттарға қарағанда, омыртқалылармен ұқсастықтары көп. Балықтар және ащылау су қоймасындағы балықтар олардың тұщы суда, теңізде бөлінуіне қызмет етеді – бұл жасанды классификацияның мысалы бола алады. Бұл классификация қоршаған ортаның айқын жағдайында бұл жануарлардың артықшылығымен негізделеді. Мұндай бөліну осморегулияцияның (судың мөлшерін реттейтін, және жануарлардың және ең қарапайым жануарлардың ағзасындағы тұздың концентрациясын реттейтін үрдіс) механизмін үйренуге қолайлы болып келеді. Сол сияқты, барлық ағзалар, микроскоптың көмегімен ғана көретінді микроағзалар деп атаймыз, оларды бір топқа байланыстыра отырып, оңай үйренуге болады, бірақ ол табиғи өзара шағылыстырылу бола алмайды.
Табиғи классификация – ағзалардың арасындағы табиғи өзара қарым – қатынастарды қолдану әрекеті. Бұл жағдайда жасанды классификацияға қарағанда қазіргілерден көп шамалар ескеріледі және бұл жерде тек сыртқы белгілер ғана емес ішкі белгілер де қарастырылады. Эмбриогенезбен, морфологиямен, анатомиямен, физиологиямен, биохимиямен, жасушалық құрылымымен және мінез құлқымен ұқсастықтары ескеріледі. Қазіргі біздің күндерімізде табиғи және шежірелік(шыққан тек)классификация басқаларға қарағанда көптеп қолданылады. Шежіре классификациясы эволюциялық өзара байланысқа негізделген. Бұл жүйеге шын көріністерге сәйкес бір топқа арғы аталары бір ағзаларды біріктіреміз. Филогения (эволюциялық тарих) осы немесе өзге топқа шыққан тек ағашымен көрініс беруі мүмкін, мысалы 2.3. суреттегідей.
Классификацияның екі түрі бар: жасанды және табиғи. Жасанды классификацияда негізге бір немесе бірнеше айырмалы белгілер алынады. Ол қолдануда ыңғайлылық және қарапайымдылық болса, практикалық тапсырмаларды қолданылады және жасап шығарылады. Жасанды классификация ерте ғасырлардағы Өытауда классификацияның бір жүйесі болып саналған. Линней барлық құрт тәрізді ағзаларды Vermes деген бір ғана топқа қосып, байланыстырды. Бұл топқа тіпті әртүрлі жануарлар да кірді, олар қарапайым жұмырдан(жұмыр құрт) және жауын құртынан, жыланға дейін барады. Линнейдің жасаған классификация тізбегі жасандыға да байланысты болып келеді, сондықтан да онда маңызды табиғи қарым – қатынас ескерілмеген, сондай – ақ, мысалы, жылан омыртқалы болса, ал жауын құртында ол жоқ. Шын мәнінде, жыландар құрттарға қарағанда, омыртқалылармен ұқсастықтары көп. Балықтар және ащылау су қоймасындағы балықтар олардың тұщы суда, теңізде бөлінуіне қызмет етеді – бұл жасанды классификацияның мысалы бола алады. Бұл классификация қоршаған ортаның айқын жағдайында бұл жануарлардың артықшылығымен негізделеді. Мұндай бөліну осморегулияцияның (судың мөлшерін реттейтін, және жануарлардың және ең қарапайым жануарлардың ағзасындағы тұздың концентрациясын реттейтін үрдіс) механизмін үйренуге қолайлы болып келеді. Сол сияқты, барлық ағзалар, микроскоптың көмегімен ғана көретінді микроағзалар деп атаймыз, оларды бір топқа байланыстыра отырып, оңай үйренуге болады, бірақ ол табиғи өзара шағылыстырылу бола алмайды.
Табиғи классификация – ағзалардың арасындағы табиғи өзара қарым – қатынастарды қолдану әрекеті. Бұл жағдайда жасанды классификацияға қарағанда қазіргілерден көп шамалар ескеріледі және бұл жерде тек сыртқы белгілер ғана емес ішкі белгілер де қарастырылады. Эмбриогенезбен, морфологиямен, анатомиямен, физиологиямен, биохимиямен, жасушалық құрылымымен және мінез құлқымен ұқсастықтары ескеріледі. Қазіргі біздің күндерімізде табиғи және шежірелік(шыққан тек)классификация басқаларға қарағанда көптеп қолданылады. Шежіре классификациясы эволюциялық өзара байланысқа негізделген. Бұл жүйеге шын көріністерге сәйкес бір топқа арғы аталары бір ағзаларды біріктіреміз. Филогения (эволюциялық тарих) осы немесе өзге топқа шыққан тек ағашымен көрініс беруі мүмкін, мысалы 2.3. суреттегідей.
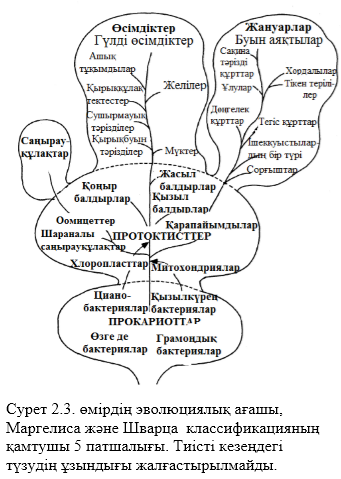
Қарастырылған классификациялардың қатарына фенотипиялық классификация да қосылады. Эволюциялық туысқандық орнатылу мәселелер әрекетінен қашуға мұндай классификация жағдай жасайды, кейде қарама – қарсылық болып және өте қиын болып кететін кездер де болады, әсіресе, қазба байлықтарының саны аз немесе мүлде жоқ болған кезде осындай жағдайға тап болады. «Фенотипикалық» сөзі грек сөзінен аударғанда, phainomenon, яғни, «біздің көріп тұрғанымыз». Бұл классификация сыртқы келбетке, яғни, көрініп тұрған белгілеріне(фенотипикалық сәйкестік) негізделген. Айта кететіні , барлық есептескіш белгілер бірдей маңызды рөл атқарады. Ағзаның әртүрлілік болатын белгілері қағида бойынша көп саналса, соншалықты жақсы болып саналады. Және олардың эволюциялық байланыстарды қамтып көрсетуі міндетті емес. Нақты деректердің саны жиналған кезде, ағзалардың арасындағы сәйкестікті негізге ала отырып, санайды; әдетте мұндайды компьютердің көмегіне жүгініп жасайды, себебі ол өте қиын болып келеді. Мұндай мақсатпен қолданылған компьютерлер сандық таксономия атына ие болды. Фенотипикалық классификация жиі филогенетикалықты еске салады, бірақ олардың шығу тарихтарында мұндай мақсат қарастырылмаған.
2.1.6. Организм анықтамасы және кілттер
Анықтамалық (диагностикалық) кестелер ағзаларды сәйкестендіру арқылы биологияда үлкен рөл атқаралы. Бұл үшін ең әуелі ағзаның берілген белгісінің тізімін біріктіреді, сосын бөлек таксономиялық диагностикалық белгілермен салыстырады. Анықтау үшін ережеге сәйкес айырмалы белгілер оңай қолданылады, мысалы, бояу формасы, аяқ – қолдар саны, сегменттер саны және т.б. Демек, анықтау жасанды немесе фенотипикалық болады, сондықтан мұндайда тек қана ағзаның сыртқы келбеті маңызды рөл ойнайды. Бұған қарамастан, диагностикалық кестелер барлық дерлік берілген таксонға иерархиялық классификацияның табиғи филогенетикалық бөлігі болып табылатын ағзаның керек заттарын анықтап бере алады.
2.1.6. Организм анықтамасы және кілттер
Анықтамалық (диагностикалық) кестелер ағзаларды сәйкестендіру арқылы биологияда үлкен рөл атқаралы. Бұл үшін ең әуелі ағзаның берілген белгісінің тізімін біріктіреді, сосын бөлек таксономиялық диагностикалық белгілермен салыстырады. Анықтау үшін ережеге сәйкес айырмалы белгілер оңай қолданылады, мысалы, бояу формасы, аяқ – қолдар саны, сегменттер саны және т.б. Демек, анықтау жасанды немесе фенотипикалық болады, сондықтан мұндайда тек қана ағзаның сыртқы келбеті маңызды рөл ойнайды. Бұған қарамастан, диагностикалық кестелер барлық дерлік берілген таксонға иерархиялық классификацияның табиғи филогенетикалық бөлігі болып табылатын ағзаның керек заттарын анықтап бере алады.
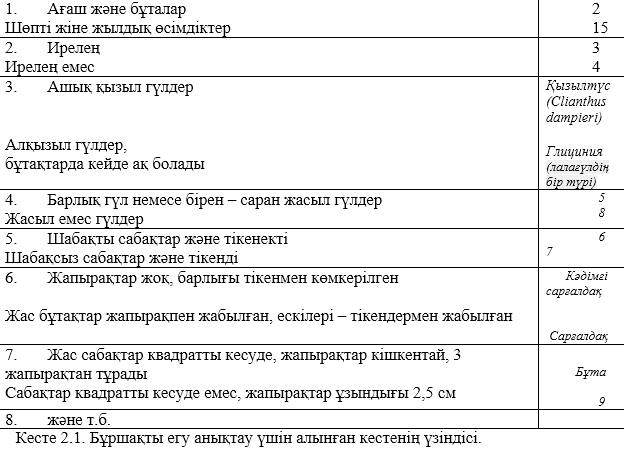
Диагностикалық кестенің әртүрлісі бар, соның ішінде ең қарапайымдысы дихотомикалық кесте. Берілген сатылардан бұл кесте жұп нөмірлі қабылдаудан құралады. Әрбір саты өзіндік анықтауын құрап отырады. Пікір бір жұпта қарама қарсы және бір жұпта бірін бірі жоққа шығаратын болуы керек. Ағзаның таксономиялық керек құрал – жабдықтарды анықтау үшін пікір жұптарын рет ретімен қарастырады. Айналымына қарай ағзалардың үлкен топтары 1 сатыдан басқасына қарай біртіндеп ыдырауға барлық кіші емес топтар түседі, сөйтіп қайта орнатылмайынша тұра береді, таксономиялық топқа берілген ағза байланысады.
Анықтамалық кестелерде қолданылатын белгілер морфологиялық және оңай айырмалы болу керек. Олар сапалы болуы мүмкін, мысалы қарынның және бояудың формасы, және санды болуы да мүмкін, мысалы шаштың саны сабақтың биіктігі. Анықтау үшін кез келген белгілерді қолдануға болады, бірақ олар үздіксіз әрі қоршаған оратының әсерінен өзгеріп кетпеу керек. Бұл жағдайда, өлшем мен бояуды қолдануға келмейді, себебі олар қоршаған ортаның әсерінен өзгеріп отырады, айталық, оның кезеңдері, жасы, немесе анықтау кезіндегі ағзаның мүшкілі. Анықтау үшін алынған белгілер бір немесе бірнеше нұсқалы болуы тиіс. Мысалы, өсімдің сабағының кескіні, оны дөңгелек қима нұсқасында, немесе тік бұрышты қима нұсқасында кездеседі.
Әрбір пікір сайын бізді тиісті сатыға жіберетін сан тұрады; берілген сатыдағы пікір, егер ағзаның сыртқы келбетімен сәйкессе, онда одан кейінгі тұрған сан келесі сатының нөмірін келесісін қарау үшін көрсетеді. Мысалы, асбұршақ пен түйебұршақты (2.1. кесте) қоса отырып, бұршақ егу анықтамасында (Leguminosae), сіз 5 – ші сатыға келдіңіз және өсімдік сабағында шабақ пен жапырағында әуелден тікен жоқ екенін көрдіңіз, онда әрі қарай 6ны өткізіп, 7 - ге көшу керек және т.с.с. (2.1. кесте).
2.2. Бес патшалық
Тағы салыстырмалы түрде жақында жалпылама мойындаумен барлық ағзалар 2 патшалыққа бөлінді, олар: жануарлар патшалығымен өсімдіктер патшалығы. Өсімдік пен жануарлардың ең негізгі айырмашылығы: олардың қоректенуінде. Кімде кім дайын қоректі жесе, соны жануарлар (гетеротрофты өнім әдісі)деп санады, ал өсімдіктер деп, өздері бейорганикалық заттан органикалық затты синтездеп
Анықтамалық кестелерде қолданылатын белгілер морфологиялық және оңай айырмалы болу керек. Олар сапалы болуы мүмкін, мысалы қарынның және бояудың формасы, және санды болуы да мүмкін, мысалы шаштың саны сабақтың биіктігі. Анықтау үшін кез келген белгілерді қолдануға болады, бірақ олар үздіксіз әрі қоршаған оратының әсерінен өзгеріп кетпеу керек. Бұл жағдайда, өлшем мен бояуды қолдануға келмейді, себебі олар қоршаған ортаның әсерінен өзгеріп отырады, айталық, оның кезеңдері, жасы, немесе анықтау кезіндегі ағзаның мүшкілі. Анықтау үшін алынған белгілер бір немесе бірнеше нұсқалы болуы тиіс. Мысалы, өсімдің сабағының кескіні, оны дөңгелек қима нұсқасында, немесе тік бұрышты қима нұсқасында кездеседі.
Әрбір пікір сайын бізді тиісті сатыға жіберетін сан тұрады; берілген сатыдағы пікір, егер ағзаның сыртқы келбетімен сәйкессе, онда одан кейінгі тұрған сан келесі сатының нөмірін келесісін қарау үшін көрсетеді. Мысалы, асбұршақ пен түйебұршақты (2.1. кесте) қоса отырып, бұршақ егу анықтамасында (Leguminosae), сіз 5 – ші сатыға келдіңіз және өсімдік сабағында шабақ пен жапырағында әуелден тікен жоқ екенін көрдіңіз, онда әрі қарай 6ны өткізіп, 7 - ге көшу керек және т.с.с. (2.1. кесте).
2.2. Бес патшалық
Тағы салыстырмалы түрде жақында жалпылама мойындаумен барлық ағзалар 2 патшалыққа бөлінді, олар: жануарлар патшалығымен өсімдіктер патшалығы. Өсімдік пен жануарлардың ең негізгі айырмашылығы: олардың қоректенуінде. Кімде кім дайын қоректі жесе, соны жануарлар (гетеротрофты өнім әдісі)деп санады, ал өсімдіктер деп, өздері бейорганикалық заттан органикалық затты синтездеп
алатындар (автотрофты өнім әдісі). Нақтырақ айтатын болсақ, гетеротрофты ағзалар, бұлар – органикалық байланыстан көміртекті бөліп алатындар, ал автотрофты ағзалар бейорганикалық формадан көміртекті қолданады, нақтырақ айтқанда, көміртек диоксиді. Әдетте, жануарларға қоректі өздері тауып іздеу керек болады , сондықтан да олар қозғалуға қабілетті болу керек. Ал бұл жерде жүйке жүйенің бар болу керектігі сезіледі, ол үлкенағзалы жануарларда үйлесімділік табады, ал өсімдік қозғалмайтын болып тіршілік етеді, олар қозғалуға қабілетсіз және оларға жүйке жүйесі керек емес.
Бірақ, бұл классификацияның түрінен бір факт айырылады, барлық жасушалы ағзалар 2 табиғи топқа ыдырайды, олар енді былай аталады: прокариоттар және эукариоттар.
Бұл екі топтың арасында үлкен айырмашылық жатыр. «Прокариот» және «эукариот» терминдері жасушадағы ДНҚ-ның қозғалысына байланысты ажыратылып көрсетіледі. Прокароитта ДНҚ ядролық мембранамен қапталмаған және цитоплазмада еркін қозғалады. Басқа сһзбен айтқанда, бұл жасушаларда тап таза ядро жоқ. Ал эукароит жасушаларында нағыз ядро бар. Эукариоттар прокариоттардан эволюциялық жолмен бірте – бірте дамыды.
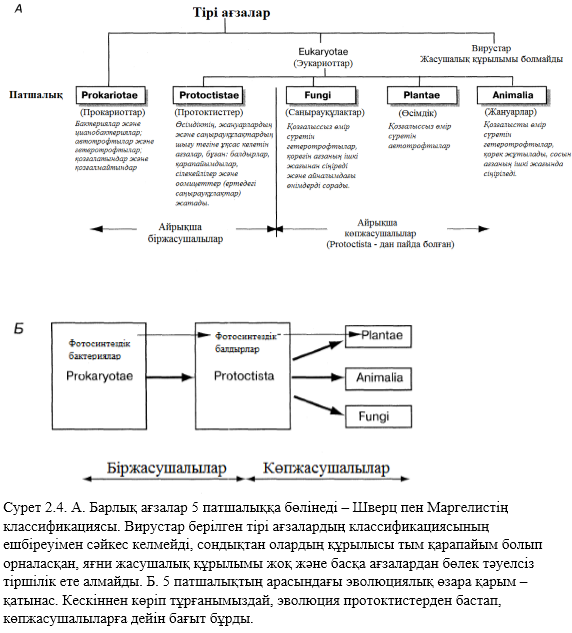
Барлық ағзалардың жануарларға және өсімдіктерге бөлінуі белгілі бір қиындықтарға ұшырайды. Мысалы, саңырауқұлақтар – гетеротрофтылар, бірақ олар қозғалмайды. Сонда оларды қай жаққа жатқызамыз? Бұл жағдайдан шығу үшін 2 патшалықтан да артық көп патшалық көп ашу керек деген шешім қабылданды. 1982 жылы Маргелис және Шварц (Margulis, Schwartz) патшылқтың жүйесін ұсынды, олар прокариот патшалығы және 4 эукариот патшалығы(2.4. сурет). Маргелис пен Шварцтың жасаған жүйесі мағызды танылымға ие болды және қолданысқа келтіре бастады. Эукариотттар Eukaryotae патшалығына жатады. Ең қарама қарсы топ – бұл протоктисттер , мүмкін олардың жасанды болу себебі де сол шығар. Бұл туралы бөлімнің 2.6. бөлігінде талқыланады.
«Ағзалардың» басқа бір тобына, классификацияның бір тобына да жатпайтын топ вирустар болып табылады. Вирустар – бұл өте – өте ұсақ бөліктер, тек қана генетикалық материалдардан (ДНҚ және РНҚ) тұрады, нәруыздық қорғаныш қабатымен қоршалады. Басқа барлық ағзалардан айырмашылығы: жасушалық құрылысы жоқ, тірі жасушаларға өтіп кетіп, көбеюге қабілетті. Вирустар табиғаты туралы 2.4. бөлігінде, ал 2.4. А. Суретінде қосымша топтар туралы белгіленеді.
Барлық ұсақ ағзалар таксономиялық табиғи бірлікке жатпаса да, олар жиі микроорганизмдер немесе микробтар деген атауға ұшырасып жатады. Бұл топ өзінің құрамына: бактерияларды, вирустарды, саңырауқұлақтарды, протоктисттерді қосады. Мұндай байланыс практикалық байланыстарды жеңілдетеді, сондықтан бұл ағзаларды үйренудегі әдістер, қағида бойынша, ұқсас болып келеді. Сонымен қатар, оларды көзбен қарау үшін микроскоп керек, ал оларды егу үшін асептикалық шараларда өткізу керек. Микроорганизмдерді үйренетін ғылым биологияның бір бұтағы микробиология деп аталады. Микроағзалар биохимияда, генетикада, агробиологияда, медицинада өте маңызды рөлге ие. Одан басқа, ол өндіріс бағытындағы биотехнологияда маңызды рөл атқарады. Бұл сұрақ 12 бөлімде талқыланады. Бактерия мен саңырауқұлақтар сияқты микроағзалар экологияда редуценттер ретінде өте маңызды рөл ойнайды.
2.3. Прокариоттар
Әдетте бактерия деп аталатын ағзалар прокариоттар патшалығына жатады. Бұл 3,5 млрд дыл бұрын пайда болған ең көне топ және жасушалық құрылымды құрайтын ұсақ ағзалар да. 2.2 кестесінде прокариоттар құрылымы туралы жинақталған. Негізі, қағида бойынша, прокариоттар біржасушалыларға жатады, біраққ көк – жасыл балдырлар жасуша тізбегін жасай алады, оны желілер деп атаймыз.
Кейбір бактериялар бір жоқ жүщім секілді ерекше жинақ жасап, бір біріне жабысып қалады, бірақ, бірімен бірі байланысып тұрған жасушалар бір – біріне тәуелсіз болып қалады. Жеке бактериялық жасушаны көру үшін тек қана микроскоптың көмегіне жүгіну арқылы көреміх, міне сондықтан да оларды микроағзалар деп атайды. Бактерияны зерттейтін ғылым – бактериология, ол микробиологияның маңызды бөлігін құрайды.
Бактериялар олардың көлемдерімен ерекшеленеді, олардың өлшемдері 0,1 мкм – ден 10 мкм – ге дейін теңселеді, ал диаметрі орташа алғанда 1 мкм болады. Орташа алғандағы ( диаметрі 5 нм) глобулярлы нәруыздың оның көлденеңіне сыйып кететіндей орын бакериялық жасушада жеткілікті. Сондықтан мұндай молекулалар 1 секундқа арақашықтыққа 60 мкм – ға тарауға қабілетті болып келеді, бұл ағзаға ешқандай арнайы транспорт механизмдері қажет еткізбейді.
Бактерия барлық жерді қамти алады: жер қыртысында, шаңда, суда жіне ауада, жануарлардың және өсімдіктердің ішінде және денесінде тіршілік етеді. Кейбір бактериялар немесе одан да үлкен болатын ыстық дерлерге қоныстанады. Ал басқалары өе тқмен температуралы тіпті мұздарда тіршілік етуге бейім болып келеді. Мұхиттың түбінде терең жарықтандыруда, қан қысымы өте жоғары және 360 өмірде сүретін бактериялар да кездеседі. Бұл мұхиттар аумағында олардан сирек қоректік тізбектер басталады. Бактериялар саны адам айтқысыз көп болып табылады. 1 грамм құнарлы жерде 2,5 млрд бактерия ұсталынады деп есептелінген; 1 см3 жаңа сүтте 3 млрд – қа артуы мүмкін. Бактерия саңылаулары басқа ағзаларға өмірлік маңызды рөл атқарады., сондықтан, нәтижесінде өзінің органикалық заттарының тіршілік әрекеттерін бұза отырып, олар табиғаттағы биогенді элементтердің айналымын қамтамасыз етеді. Одан басқа, олар адам өміріндегі маңызды заттарға ие, ол тек адам ағзасындағы ауруларды қоздырғышы емес, оның бтохимиялық реакцияларына қолданылуы мүмкін. Бұл туралы толығырақ 12 бөлімде айтылады.
Бірақ, бұл классификацияның түрінен бір факт айырылады, барлық жасушалы ағзалар 2 табиғи топқа ыдырайды, олар енді былай аталады: прокариоттар және эукариоттар.
Бұл екі топтың арасында үлкен айырмашылық жатыр. «Прокариот» және «эукариот» терминдері жасушадағы ДНҚ-ның қозғалысына байланысты ажыратылып көрсетіледі. Прокароитта ДНҚ ядролық мембранамен қапталмаған және цитоплазмада еркін қозғалады. Басқа сһзбен айтқанда, бұл жасушаларда тап таза ядро жоқ. Ал эукароит жасушаларында нағыз ядро бар. Эукариоттар прокариоттардан эволюциялық жолмен бірте – бірте дамыды.
Барлық ағзалардың жануарларға және өсімдіктерге бөлінуі белгілі бір қиындықтарға ұшырайды. Мысалы, саңырауқұлақтар – гетеротрофтылар, бірақ олар қозғалмайды. Сонда оларды қай жаққа жатқызамыз? Бұл жағдайдан шығу үшін 2 патшалықтан да артық көп патшалық көп ашу керек деген шешім қабылданды. 1982 жылы Маргелис және Шварц (Margulis, Schwartz) патшылқтың жүйесін ұсынды, олар прокариот патшалығы және 4 эукариот патшалығы(2.4. сурет). Маргелис пен Шварцтың жасаған жүйесі мағызды танылымға ие болды және қолданысқа келтіре бастады. Эукариотттар Eukaryotae патшалығына жатады. Ең қарама қарсы топ – бұл протоктисттер , мүмкін олардың жасанды болу себебі де сол шығар. Бұл туралы бөлімнің 2.6. бөлігінде талқыланады.
«Ағзалардың» басқа бір тобына, классификацияның бір тобына да жатпайтын топ вирустар болып табылады. Вирустар – бұл өте – өте ұсақ бөліктер, тек қана генетикалық материалдардан (ДНҚ және РНҚ) тұрады, нәруыздық қорғаныш қабатымен қоршалады. Басқа барлық ағзалардан айырмашылығы: жасушалық құрылысы жоқ, тірі жасушаларға өтіп кетіп, көбеюге қабілетті. Вирустар табиғаты туралы 2.4. бөлігінде, ал 2.4. А. Суретінде қосымша топтар туралы белгіленеді.
Барлық ұсақ ағзалар таксономиялық табиғи бірлікке жатпаса да, олар жиі микроорганизмдер немесе микробтар деген атауға ұшырасып жатады. Бұл топ өзінің құрамына: бактерияларды, вирустарды, саңырауқұлақтарды, протоктисттерді қосады. Мұндай байланыс практикалық байланыстарды жеңілдетеді, сондықтан бұл ағзаларды үйренудегі әдістер, қағида бойынша, ұқсас болып келеді. Сонымен қатар, оларды көзбен қарау үшін микроскоп керек, ал оларды егу үшін асептикалық шараларда өткізу керек. Микроорганизмдерді үйренетін ғылым биологияның бір бұтағы микробиология деп аталады. Микроағзалар биохимияда, генетикада, агробиологияда, медицинада өте маңызды рөлге ие. Одан басқа, ол өндіріс бағытындағы биотехнологияда маңызды рөл атқарады. Бұл сұрақ 12 бөлімде талқыланады. Бактерия мен саңырауқұлақтар сияқты микроағзалар экологияда редуценттер ретінде өте маңызды рөл ойнайды.
2.3. Прокариоттар
Әдетте бактерия деп аталатын ағзалар прокариоттар патшалығына жатады. Бұл 3,5 млрд дыл бұрын пайда болған ең көне топ және жасушалық құрылымды құрайтын ұсақ ағзалар да. 2.2 кестесінде прокариоттар құрылымы туралы жинақталған. Негізі, қағида бойынша, прокариоттар біржасушалыларға жатады, біраққ көк – жасыл балдырлар жасуша тізбегін жасай алады, оны желілер деп атаймыз.
Кейбір бактериялар бір жоқ жүщім секілді ерекше жинақ жасап, бір біріне жабысып қалады, бірақ, бірімен бірі байланысып тұрған жасушалар бір – біріне тәуелсіз болып қалады. Жеке бактериялық жасушаны көру үшін тек қана микроскоптың көмегіне жүгіну арқылы көреміх, міне сондықтан да оларды микроағзалар деп атайды. Бактерияны зерттейтін ғылым – бактериология, ол микробиологияның маңызды бөлігін құрайды.
Бактериялар олардың көлемдерімен ерекшеленеді, олардың өлшемдері 0,1 мкм – ден 10 мкм – ге дейін теңселеді, ал диаметрі орташа алғанда 1 мкм болады. Орташа алғандағы ( диаметрі 5 нм) глобулярлы нәруыздың оның көлденеңіне сыйып кететіндей орын бакериялық жасушада жеткілікті. Сондықтан мұндай молекулалар 1 секундқа арақашықтыққа 60 мкм – ға тарауға қабілетті болып келеді, бұл ағзаға ешқандай арнайы транспорт механизмдері қажет еткізбейді.
Бактерия барлық жерді қамти алады: жер қыртысында, шаңда, суда жіне ауада, жануарлардың және өсімдіктердің ішінде және денесінде тіршілік етеді. Кейбір бактериялар немесе одан да үлкен болатын ыстық дерлерге қоныстанады. Ал басқалары өе тқмен температуралы тіпті мұздарда тіршілік етуге бейім болып келеді. Мұхиттың түбінде терең жарықтандыруда, қан қысымы өте жоғары және 360 өмірде сүретін бактериялар да кездеседі. Бұл мұхиттар аумағында олардан сирек қоректік тізбектер басталады. Бактериялар саны адам айтқысыз көп болып табылады. 1 грамм құнарлы жерде 2,5 млрд бактерия ұсталынады деп есептелінген; 1 см3 жаңа сүтте 3 млрд – қа артуы мүмкін. Бактерия саңылаулары басқа ағзаларға өмірлік маңызды рөл атқарады., сондықтан, нәтижесінде өзінің органикалық заттарының тіршілік әрекеттерін бұза отырып, олар табиғаттағы биогенді элементтердің айналымын қамтамасыз етеді. Одан басқа, олар адам өміріндегі маңызды заттарға ие, ол тек адам ағзасындағы ауруларды қоздырғышы емес, оның бтохимиялық реакцияларына қолданылуы мүмкін. Бұл туралы толығырақ 12 бөлімде айтылады.
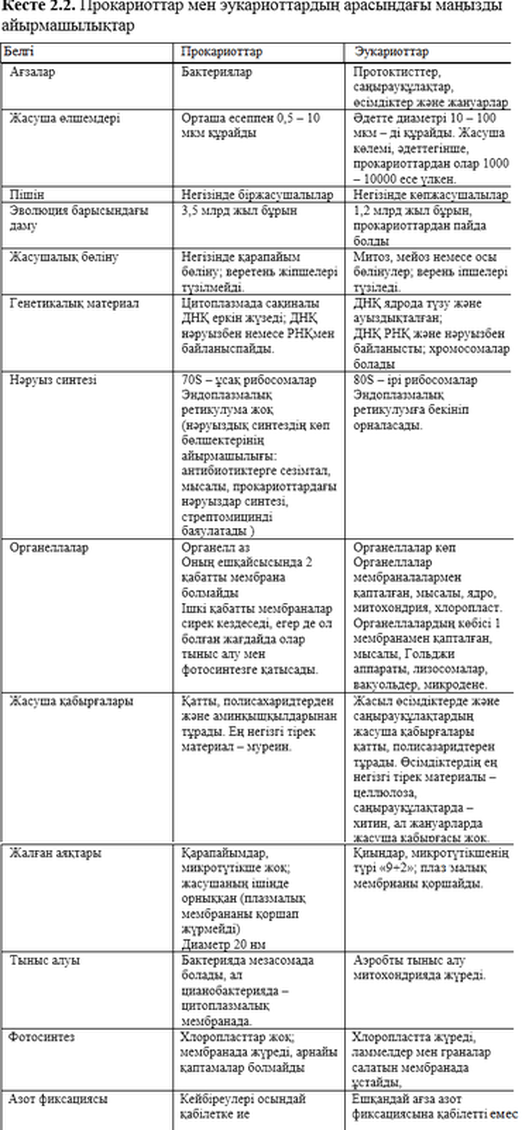
2.3.1. Бактериялардың құрылымы
2.5. Суретте жалпылама бактериялардың құрылысы көрсетілген – әдеттегі прокариоттардың жасушасы.
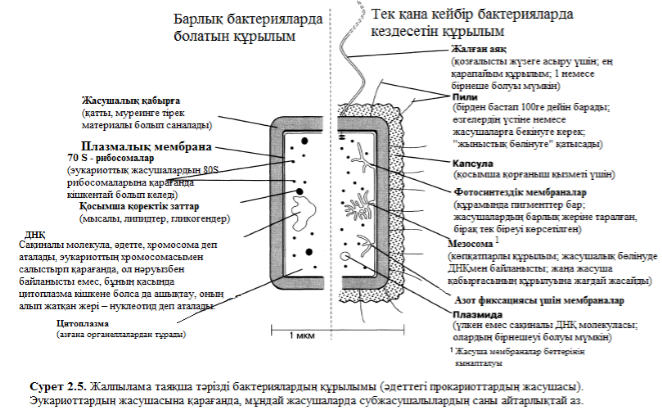
2.6. Суретінде, А – Г ең көп тараған таяқшатәрізді бактерия – Escherichia coli суреттелген. Әдетте ол мүлдем зиянды емес. Оның бары суда фекальмен ластанған судың ең үздік көрсеткіші ретінде қолдануға болады. Барлық бактериялардың ішінен осы E.Coli көп зерттелген. Одан басқа, бұл бактерияның генетикалық картасы толығымен жасалған. Эукариот жасушаларына қарағанда, E.Coli-дің жасуша ішілік құрылысы аз, осыған аса мән беру керек(5.10 және 5.11. суреттер). 2.7. суретінде таяқша тәрізді бактерия көрсетілген., оның E.Coli-ден айырмашылығы: жалған аяғы бар.
Жасушалық қабырға
Бактерияның жасушалық қабырғасы – құрылымы айтарлықтай берік және пішінін сақтауға көмектеседі. Ол муреинмен қамтамасыз етілген, муреин – полисахаридті тізбектен тұратын молекулалар, айқастырылып үздіксіз аралықтар арқылы байланысқан қысұаша аминқышқыл тізбектері. Осылайша, әрбір жасуша бір үлкен молекуланың шаруасында көрсетілгендей торлы қапшықпен қапталған. Жасушалық қабырға жасушаға су түсіп, жарылып кетуінен сақтайды (мысалы, осмостың нәтижесінде). Су иондары және кішкентай молекулалар жасушаға жасушалық қабырғаның қуыстарынан өтіп кіреді.
1884 жылы даттық биолог Кристиан Грам бояу әдісін бактериялардвң 2 топқа бөлінетіндігін , және қазір бәріне белгілі болғандай олардың жасушалық қабырғасының айырмашылығымын қамтамасыз етілетінін пайдаланып, оны дамытты. Грам әдісімен боялынатын бір бактериялар, грамоң деген атау алды, ал боянылмайтындар грамтерістік деген атау алды. Грам бойынша бояу туралы практикалық жұмыстар 12.9.2. бөлімінде жазылған.
2.5. Суретте жалпылама бактериялардың құрылысы көрсетілген – әдеттегі прокариоттардың жасушасы.
2.6. Суретінде, А – Г ең көп тараған таяқшатәрізді бактерия – Escherichia coli суреттелген. Әдетте ол мүлдем зиянды емес. Оның бары суда фекальмен ластанған судың ең үздік көрсеткіші ретінде қолдануға болады. Барлық бактериялардың ішінен осы E.Coli көп зерттелген. Одан басқа, бұл бактерияның генетикалық картасы толығымен жасалған. Эукариот жасушаларына қарағанда, E.Coli-дің жасуша ішілік құрылысы аз, осыған аса мән беру керек(5.10 және 5.11. суреттер). 2.7. суретінде таяқша тәрізді бактерия көрсетілген., оның E.Coli-ден айырмашылығы: жалған аяғы бар.
Жасушалық қабырға
Бактерияның жасушалық қабырғасы – құрылымы айтарлықтай берік және пішінін сақтауға көмектеседі. Ол муреинмен қамтамасыз етілген, муреин – полисахаридті тізбектен тұратын молекулалар, айқастырылып үздіксіз аралықтар арқылы байланысқан қысұаша аминқышқыл тізбектері. Осылайша, әрбір жасуша бір үлкен молекуланың шаруасында көрсетілгендей торлы қапшықпен қапталған. Жасушалық қабырға жасушаға су түсіп, жарылып кетуінен сақтайды (мысалы, осмостың нәтижесінде). Су иондары және кішкентай молекулалар жасушаға жасушалық қабырғаның қуыстарынан өтіп кіреді.
1884 жылы даттық биолог Кристиан Грам бояу әдісін бактериялардвң 2 топқа бөлінетіндігін , және қазір бәріне белгілі болғандай олардың жасушалық қабырғасының айырмашылығымын қамтамасыз етілетінін пайдаланып, оны дамытты. Грам әдісімен боялынатын бір бактериялар, грамоң деген атау алды, ал боянылмайтындар грамтерістік деген атау алды. Грам бойынша бояу туралы практикалық жұмыстар 12.9.2. бөлімінде жазылған.
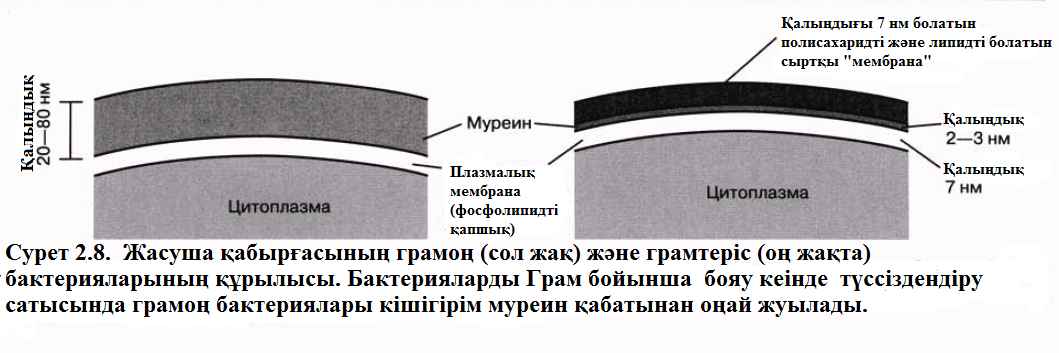
Staphylococcus, Bacillus және Lactobacillus сияқты грамоң бактерияларында муреиндік торда басқа да компоненттер бар, негізінде жасушалық қабырғаны толық қылатын полисахаридтер мен нәруыздар. Salmonella,E. Coli және Azotobacter сияқты грамтерістік бактериялардың жасушалық қабықшасы кішкентай және қиын құрылымды болып келеді. Бұл бактерияларда муреинді қабаттары сырттарынан липидті және полисахаридті кішігірім әрі жұмсақ мембранатәрізді қабатпен қаптайды, лизоцимадан жасушаларды қорғайды, лизоцома – антибиотикті бактерия, көз жасында, сілекейде және тағы да басқа биологиялық сұйықтарда және де піскен жұмыртқаның ақуызында кездеседі. Лизоцим полиахаридті муреин қаңқасын жарады, яғни жасушалық қабырғаның тесілуіне, яғни осмотикалық ісінуге және жарылуға әкеледі. Грамоң бактерияларында өсетін айқасқан байланыстарды муреинде пайда болуын құлыптап тастайды, яғни осмотикалық шокқа олардың жасушаларын көбірек әлсіз болу үшін істейді.
Плазмалық мембрана, мезосомалар және фотосинтездік мембраналар
Басқа ағзалар секілді бактерия жасушасының тірі заты жартылай өткізгіш жарғақпен қоршалған. Бактерия жасушасының плазмалық мембраналары құрылысы мен функциясы жағынан эукариоттардың плазмалық мембранасынан аумайды(5.9. бөлім). Олар тыныс алу ферменттерінің қозғалыс орындарына қызмет етеді, ал кейбір бактерияларда ол мезослма және (немесе) фотосинтездік мембранаға ұқсайды.
Мезосомалар – плазмалық мембрана жасушаларын өзінің бойынан дүрдитіп тұрушы қатпарлы құрылым. Жасушалық бөліну кезінде, бәлкім, ДНҚ – мен байланысады, өзіне сәйкес өндіруден кейінгі ДНҚ-ның 2 еншілес молекуласын қамтамасыз етеді жіне еншілес молекулалардың арасында аралық пайда болуын қарастырады.
Қап сияқты, түтікшетәрізді және қатпарлы фотосинтездік бактерияларда плазмалық мембраналар фотосинтездік пигменттерді дүрдитіп тұрады (ол кезде міндетті түрде бактериохлорофилл). Үйлес мембраналар азот фиксациясының пайда болуына қатысады.
Генетикалық мембрана (бактериялды «хромосома»)
Бактериялды ДНҚ жалғыз сақиналы ұзындығы жасушасынан 1 мм үлкен болып келетін, негіздің шамамен 5 млн. жұбы бар. Жалпы ДНҚ ның ұстанымы және оның ішіндегі сиқырланғандардың саны эукариоттардың жасушасына қарағанда бактерия жасушасында айтарлықтай аз, әдетте ДНҚ бактерияларында адамның жасушасымен салыстырып қарағанда 50 есе кіші бірнеше мың гендер бар.
Рибосомалар
Нәруыздардың орнына қызмет етеді (2.2 кестесінде және 5.5 суретінде).
Капсулалар
Кейбір бактерияларда шырышты немесе желімді құпиялар капсулаға айналады. Жағымсыз қарама – қарсылықтан кейін капсулалар жақсы көрінеді. Кейбір кездерде бұл құпиялар колония жасауда қызмет етеді. Осы құпия арқасында бактерияларда әртүрлі биіктіктерге жабысып қалу қалыптасады, тістер сияқты жартас – құздардың бөліктері. Одан басқа , капсулалар бактерия жасушаларына қосымша ұорғаныш қызметін атқарады. Мысалы, пневмококктың (өкпе қабынуын қоздырушы бактериялар) капсуларлы штаммдары оңай қабынулар арқасында адам ағзасында еркін көбейе алады, ал капсуларлы штаммдар емес кезде олар оңай шауыл жасай алады және фагоциттермен бұзылады эәне сол үшін қауіпсіз болып келеді.
Спора (гүлсіз өсімдіктердің және кейбір бір жасушалы жәндіктердің тұқымы)
Clostridium және Bacillus түрлеріне жататын кейбір бактериялар эндоспера түзеді (яғни, жасуша ішінде тіршілік ететін споралар). Споралар өздері ұзақөмірсүргіштік толық болып келеді, өте үлкен тұрақтылықпен ерекшеленіледі, әсіресе, жылытуға, қысқа толқынды сәулеленуге және құрғатылуға. Спораның жасушадағы қозғалысы әртүрлі болады, сәйкестендіру үшін маңызды беліглермен және бактерия классификациясына қызмет етеді (2.10. сурет).
Жалған аяқтар
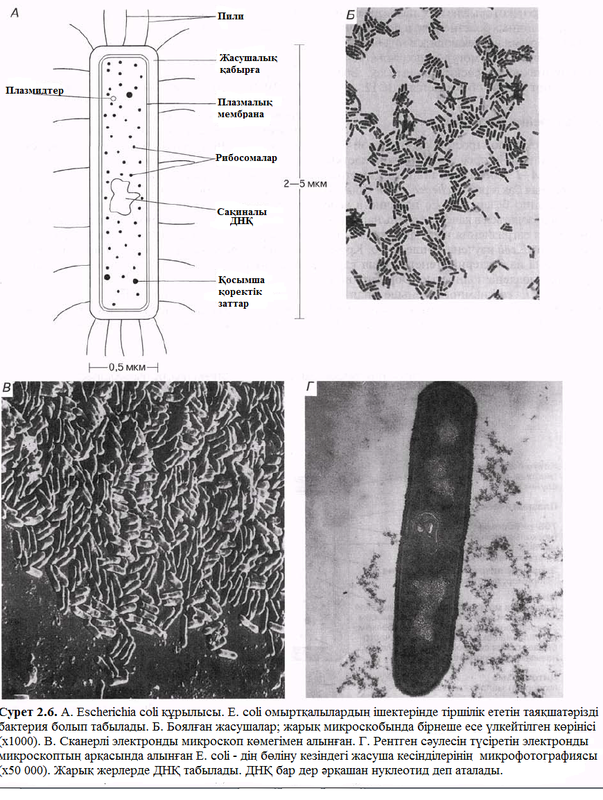
Бір немесе бірнеше жалған аяқтармен қамтамасыз етілгені үшін бактериялардың көбісі қамтамасыз етілген. Жалған аяқ – бұл бірдей нәруыз молекулалары бар қарапайым бос цилиндр. Өздерінің толқынды болғанына қарамастан, қатты болып келеді (2.7. сурет). Бактериялардың қозғалысы жалған аяқтың айналуына дейін жетеді, нәтижесінде, жалған аяқ ортада ретсіз соғысһлыстарды істемей бұралады және осындайлығымен бактерияны өзінің артынан қозғалтады. Мысал ретінде, мына бактерияларды алсақ болады: Rhizobium (бір жалған аяқ)және Azotobacter (бірнеше жалған аяқ), бұл бактериялапһр табиғаттығы азотты айналымына қатысады.
Қозғалатын бактериялар тітіркендіргіштерге жауап қайтаруда қощғалады, яғни, олар таксиске қабілетті. Аэробты бактериялар, мысалы, ортаның концентрлі оттегін арттырған кезде жылжиды (оң аэротаксис пайда болады), ал қозғалатын фотосинтезді бактериялар жарыққа қарай ағылады (оң фототаксис пайда болады). Жалған аяқтар бәрінен де шаңдату әдісін қолданған кезед электронды микроскопта жақсы көрінеді.
Пили
Жасуша қабырғасында кейбір грамтерістік бактерияларда көпсанды кішігірім таяқшатәрізді өсінді ретінде көрінеді, яғни олар пили немес фимбрии деп аталады. Пили қысқа әрі кішігірім жалған аяқтары бар және спецификалық жасушаларға немесе үстінде бекінуге қвзмет етеді. Пилидің көптеген түрлері белгілі, мысалы, F – пили, ол жыныстық көбеюге қатысады.
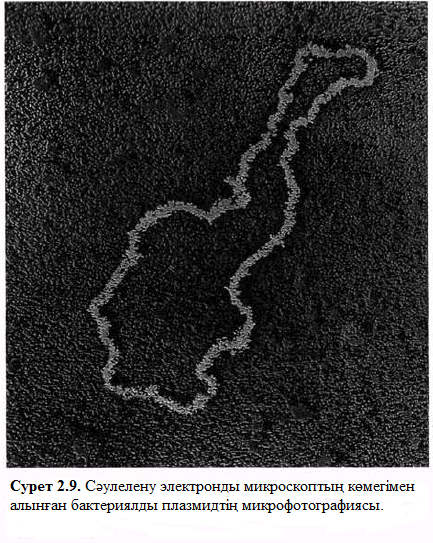
Плазмидтер
Сонымен қатар, бактерияның барлық түрінде болатын жалғыз ДНҚ молекулалары, оның кейбіреулері бір немесе бірнеше плазмидтер көрсетеді (2.9. сурет). Плазмида – ДНҚ – ның қосымша молекуласы, өздігінен қайтатүзілуге қабілетті. Плазмида өзіне көбірек өмір сүретін жасушаларды қамтамасыз ететін бірнеше гендерді алып жүреді. Кейбір бактериялар жасушаны антибиотиктерге тұрақты етеді. Мысалы, плазмида жасушасында бөлшектенген пеницилині бар пенициллин-ферментін алып жүретін кейбір стафилококтар бар.
Нәтижесінде, жасуша пенициллинге тұрақтылық арттырады. Конъюгация кезінде мұндай гендердің таралуы (2.3.3. бөлімінде) медицинада маңызды рөл атқарады. Гендердік бөлімдерінде басқа да плазмидті гендер бар,
Плазмалық мембрана, мезосомалар және фотосинтездік мембраналар
Басқа ағзалар секілді бактерия жасушасының тірі заты жартылай өткізгіш жарғақпен қоршалған. Бактерия жасушасының плазмалық мембраналары құрылысы мен функциясы жағынан эукариоттардың плазмалық мембранасынан аумайды(5.9. бөлім). Олар тыныс алу ферменттерінің қозғалыс орындарына қызмет етеді, ал кейбір бактерияларда ол мезослма және (немесе) фотосинтездік мембранаға ұқсайды.
Мезосомалар – плазмалық мембрана жасушаларын өзінің бойынан дүрдитіп тұрушы қатпарлы құрылым. Жасушалық бөліну кезінде, бәлкім, ДНҚ – мен байланысады, өзіне сәйкес өндіруден кейінгі ДНҚ-ның 2 еншілес молекуласын қамтамасыз етеді жіне еншілес молекулалардың арасында аралық пайда болуын қарастырады.
Қап сияқты, түтікшетәрізді және қатпарлы фотосинтездік бактерияларда плазмалық мембраналар фотосинтездік пигменттерді дүрдитіп тұрады (ол кезде міндетті түрде бактериохлорофилл). Үйлес мембраналар азот фиксациясының пайда болуына қатысады.
Генетикалық мембрана (бактериялды «хромосома»)
Бактериялды ДНҚ жалғыз сақиналы ұзындығы жасушасынан 1 мм үлкен болып келетін, негіздің шамамен 5 млн. жұбы бар. Жалпы ДНҚ ның ұстанымы және оның ішіндегі сиқырланғандардың саны эукариоттардың жасушасына қарағанда бактерия жасушасында айтарлықтай аз, әдетте ДНҚ бактерияларында адамның жасушасымен салыстырып қарағанда 50 есе кіші бірнеше мың гендер бар.
Рибосомалар
Нәруыздардың орнына қызмет етеді (2.2 кестесінде және 5.5 суретінде).
Капсулалар
Кейбір бактерияларда шырышты немесе желімді құпиялар капсулаға айналады. Жағымсыз қарама – қарсылықтан кейін капсулалар жақсы көрінеді. Кейбір кездерде бұл құпиялар колония жасауда қызмет етеді. Осы құпия арқасында бактерияларда әртүрлі биіктіктерге жабысып қалу қалыптасады, тістер сияқты жартас – құздардың бөліктері. Одан басқа , капсулалар бактерия жасушаларына қосымша ұорғаныш қызметін атқарады. Мысалы, пневмококктың (өкпе қабынуын қоздырушы бактериялар) капсуларлы штаммдары оңай қабынулар арқасында адам ағзасында еркін көбейе алады, ал капсуларлы штаммдар емес кезде олар оңай шауыл жасай алады және фагоциттермен бұзылады эәне сол үшін қауіпсіз болып келеді.
Спора (гүлсіз өсімдіктердің және кейбір бір жасушалы жәндіктердің тұқымы)
Clostridium және Bacillus түрлеріне жататын кейбір бактериялар эндоспера түзеді (яғни, жасуша ішінде тіршілік ететін споралар). Споралар өздері ұзақөмірсүргіштік толық болып келеді, өте үлкен тұрақтылықпен ерекшеленіледі, әсіресе, жылытуға, қысқа толқынды сәулеленуге және құрғатылуға. Спораның жасушадағы қозғалысы әртүрлі болады, сәйкестендіру үшін маңызды беліглермен және бактерия классификациясына қызмет етеді (2.10. сурет).
Жалған аяқтар
Бір немесе бірнеше жалған аяқтармен қамтамасыз етілгені үшін бактериялардың көбісі қамтамасыз етілген. Жалған аяқ – бұл бірдей нәруыз молекулалары бар қарапайым бос цилиндр. Өздерінің толқынды болғанына қарамастан, қатты болып келеді (2.7. сурет). Бактериялардың қозғалысы жалған аяқтың айналуына дейін жетеді, нәтижесінде, жалған аяқ ортада ретсіз соғысһлыстарды істемей бұралады және осындайлығымен бактерияны өзінің артынан қозғалтады. Мысал ретінде, мына бактерияларды алсақ болады: Rhizobium (бір жалған аяқ)және Azotobacter (бірнеше жалған аяқ), бұл бактериялапһр табиғаттығы азотты айналымына қатысады.
Қозғалатын бактериялар тітіркендіргіштерге жауап қайтаруда қощғалады, яғни, олар таксиске қабілетті. Аэробты бактериялар, мысалы, ортаның концентрлі оттегін арттырған кезде жылжиды (оң аэротаксис пайда болады), ал қозғалатын фотосинтезді бактериялар жарыққа қарай ағылады (оң фототаксис пайда болады). Жалған аяқтар бәрінен де шаңдату әдісін қолданған кезед электронды микроскопта жақсы көрінеді.
Пили
Жасуша қабырғасында кейбір грамтерістік бактерияларда көпсанды кішігірім таяқшатәрізді өсінді ретінде көрінеді, яғни олар пили немес фимбрии деп аталады. Пили қысқа әрі кішігірім жалған аяқтары бар және спецификалық жасушаларға немесе үстінде бекінуге қвзмет етеді. Пилидің көптеген түрлері белгілі, мысалы, F – пили, ол жыныстық көбеюге қатысады.
Плазмидтер
Сонымен қатар, бактерияның барлық түрінде болатын жалғыз ДНҚ молекулалары, оның кейбіреулері бір немесе бірнеше плазмидтер көрсетеді (2.9. сурет). Плазмида – ДНҚ – ның қосымша молекуласы, өздігінен қайтатүзілуге қабілетті. Плазмида өзіне көбірек өмір сүретін жасушаларды қамтамасыз ететін бірнеше гендерді алып жүреді. Кейбір бактериялар жасушаны антибиотиктерге тұрақты етеді. Мысалы, плазмида жасушасында бөлшектенген пеницилині бар пенициллин-ферментін алып жүретін кейбір стафилококтар бар.
Нәтижесінде, жасуша пенициллинге тұрақтылық арттырады. Конъюгация кезінде мұндай гендердің таралуы (2.3.3. бөлімінде) медицинада маңызды рөл атқарады. Гендердік бөлімдерінде басқа да плазмидті гендер бар,
- дәрі жасау ортасына тұрақтылық береді;
- әртүрлі аурулар шақырады;
- ірімшік жасау кезінде сүттің сүтқышқылдарымен ашытылуына жауап береді;
- көмірсулар сияқты тамақтарды қиын химиялық заттармен қолдануға қабілеттілік береді , және сондықтан мұнай дақтар тартысында және мұнайөнімдерінен нәруыздарды алуда қолдануға потенциалды керек .
2.3.2. Жасуша формасы
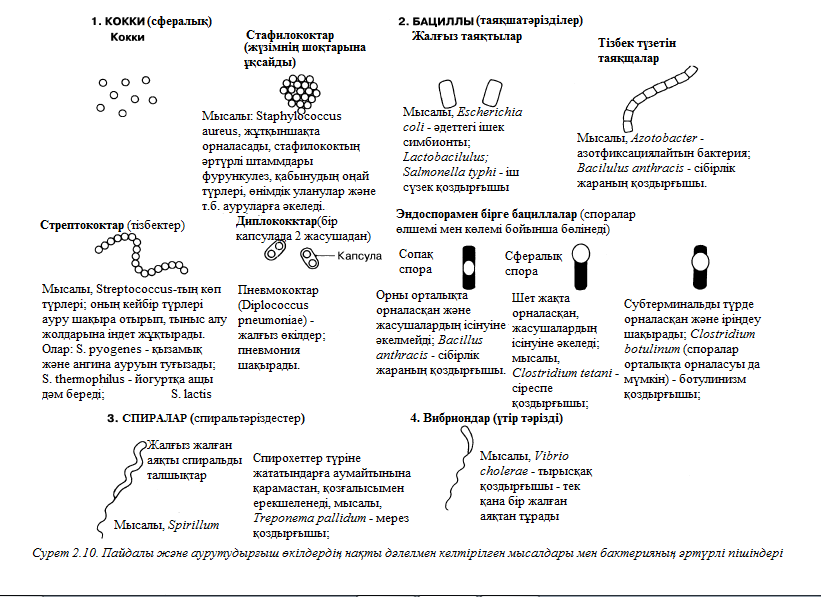
Бактерия жасушаларының формасы систематикалық белгілердің ішінде ең маңыздысы болып табылады. 2.10. суретінде жасушаның негізгі 4 түрі жазылған. Бұл суретте қалай пайдалы болатынын және ауруға әкелетін бактериялар туралы көрсетілген.
2.3.3. Көбею
Жеке өсу және жыныссыз көбею
Бактериалды жасушалардың беті/көлем арақатынастары өте маңызды рөл атқарады, диффузия мен белсенді транспорттың орнына қоршаған ортадан қоректік заттарды тез сіңіруге бейім келеді. Сондықтан бактериялар қолайлы жағдайларда тез өседі. Бактериалды жасушалардың дамуы температура, қоректік заттар, рН ортасы мен иондар концентрациясы сияқты үлкен дәрежеде орта факторларына тәуелді болып келеді. Одан басқа, міндетті түрде аэробқа оттек керек болса, міндетті түрде анаэробты түрге оның болмағаны керек.
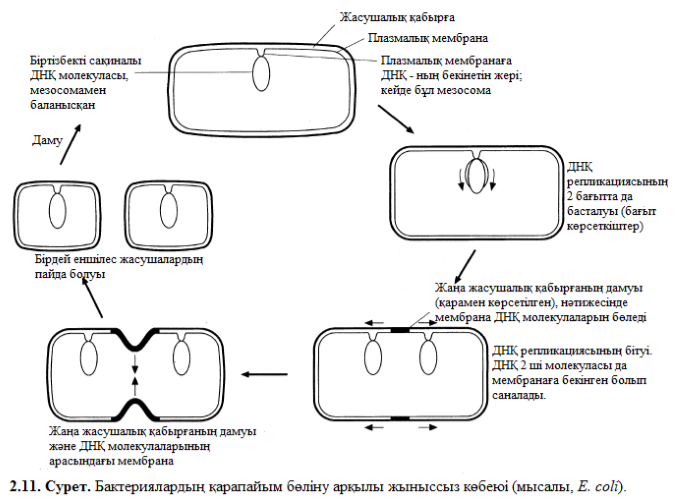
Ядро мен цитоплазманың өлшемдер арақатынасы еріксіз белгілі бір өлшемге жеткен кезде бактериялар қарапайым жыныссыз бөліну жолымен өтеді, яғни 2 бірдей еншілес жасушаларға бөліну жолымен. Жасушалық бөлінуге ең алдымен ДНҚ репликациясы жүреді, соған шейін, репликация процесі бітпейді, ДНҚ – ны мезосомалар белгілі бір жағдайда ғана ұстай алады.
Еншілес жасушалардың арасында түзілетін мезосомалар жаңа аралытарға бекінеді, жасушалық қабырғаларда синтез материалы болып табылады. Нағыз тез өсетін бактериялардың бөлінісі әрбір 20 мин сайын жүріп отырады.
Глюкозаның жеткілікті санының ортадағы нормасымен және бейорганикалық E.Coli тұздары барлық керек аминқышқылдарын өздері синтездеді. Бұл бактерияларды зерттеу нәтижесінде оларда кейде кездейсоқ мутациялар орын алады. Мутанттардың 2 түрі белгіленген: біріншісі – биотин (витамин) мен метионин аминқышқылын синтездеуге қабілетсіз, және екіншісі – треонин мен лейцин аминқышылдарын синтездеуге қабілетсіз. Дамудың барлық 4 факторын құрай алмайтын ортасына 108 жасушаны әрбір мутанттық штаммға аударған. Теоретикалық жасушалар бұл ортада өспеуі мідетті емес еді. Бірақ бірнеше жүздеген колониялар алынды (әрбір колония алғашқы жалғыз жасушадан пайда болып отырады) неге олай десеңіз, бұл жасушаларда даму 4 түрлі факторларына қажетті барлық гендер бар. Демек, жасушаларда генетикалық заттарды бөліп шығару процесіне қатысты ақпаратттарды алмастыруға болады, бірақ ол уақытта мүмкін емес болды. Соңында электронды микроскоптың арқасында E.coli жасушалары бір – бірімен тікелей қосыла алатынын орнатылған, яғни оларда конъюгация процесі жүре алады.
Бактерия жасушаларының формасы систематикалық белгілердің ішінде ең маңыздысы болып табылады. 2.10. суретінде жасушаның негізгі 4 түрі жазылған. Бұл суретте қалай пайдалы болатынын және ауруға әкелетін бактериялар туралы көрсетілген.
2.3.3. Көбею
Жеке өсу және жыныссыз көбею
Бактериалды жасушалардың беті/көлем арақатынастары өте маңызды рөл атқарады, диффузия мен белсенді транспорттың орнына қоршаған ортадан қоректік заттарды тез сіңіруге бейім келеді. Сондықтан бактериялар қолайлы жағдайларда тез өседі. Бактериалды жасушалардың дамуы температура, қоректік заттар, рН ортасы мен иондар концентрациясы сияқты үлкен дәрежеде орта факторларына тәуелді болып келеді. Одан басқа, міндетті түрде аэробқа оттек керек болса, міндетті түрде анаэробты түрге оның болмағаны керек.
Ядро мен цитоплазманың өлшемдер арақатынасы еріксіз белгілі бір өлшемге жеткен кезде бактериялар қарапайым жыныссыз бөліну жолымен өтеді, яғни 2 бірдей еншілес жасушаларға бөліну жолымен. Жасушалық бөлінуге ең алдымен ДНҚ репликациясы жүреді, соған шейін, репликация процесі бітпейді, ДНҚ – ны мезосомалар белгілі бір жағдайда ғана ұстай алады.
Еншілес жасушалардың арасында түзілетін мезосомалар жаңа аралытарға бекінеді, жасушалық қабырғаларда синтез материалы болып табылады. Нағыз тез өсетін бактериялардың бөлінісі әрбір 20 мин сайын жүріп отырады.
Глюкозаның жеткілікті санының ортадағы нормасымен және бейорганикалық E.Coli тұздары барлық керек аминқышқылдарын өздері синтездеді. Бұл бактерияларды зерттеу нәтижесінде оларда кейде кездейсоқ мутациялар орын алады. Мутанттардың 2 түрі белгіленген: біріншісі – биотин (витамин) мен метионин аминқышқылын синтездеуге қабілетсіз, және екіншісі – треонин мен лейцин аминқышылдарын синтездеуге қабілетсіз. Дамудың барлық 4 факторын құрай алмайтын ортасына 108 жасушаны әрбір мутанттық штаммға аударған. Теоретикалық жасушалар бұл ортада өспеуі мідетті емес еді. Бірақ бірнеше жүздеген колониялар алынды (әрбір колония алғашқы жалғыз жасушадан пайда болып отырады) неге олай десеңіз, бұл жасушаларда даму 4 түрлі факторларына қажетті барлық гендер бар. Демек, жасушаларда генетикалық заттарды бөліп шығару процесіне қатысты ақпаратттарды алмастыруға болады, бірақ ол уақытта мүмкін емес болды. Соңында электронды микроскоптың арқасында E.coli жасушалары бір – бірімен тікелей қосыла алатынын орнатылған, яғни оларда конъюгация процесі жүре алады.
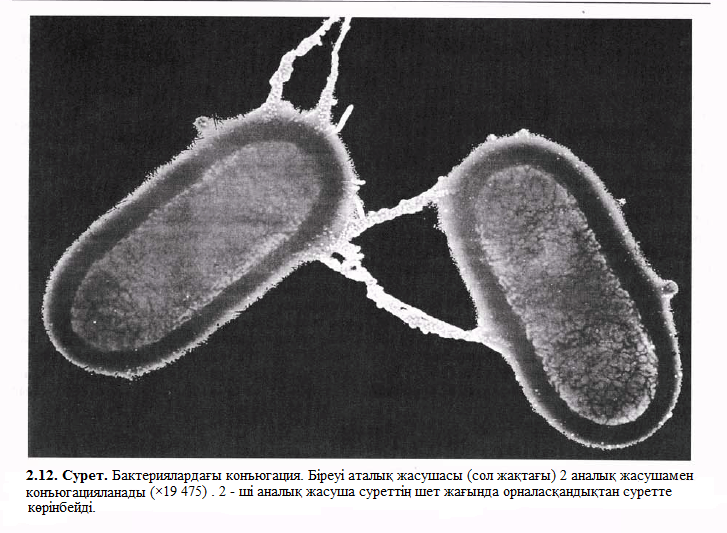
Осылайша, конъюгацияда тікелей қосылу нәтижесінде жасушалар арасында ДНҚ тасымалдау жүреді. Бір жасуша бұл жағдайда донор («аталық» жасуша») болса, екінші біреуі – рецепиент («аналық» жасуша») болады. Жасушалардың донор болып қызмет етуі жыныс факторы немесе F – фактор деп аталатын ерекше плазмидте болатын генмен анықталады(F – ағылшынша fertility – ұрықтылық дегенді білдіреді). Бұл гендерде арнайы пилдердің нәруыздары сиқырланған, бұлар F – пилдер немесе жыныстық пилдер деп аталады. F – пилдер жасушасындағы байланысқа конъюгация кезінде қатысады. Пилдер – жыныстық құрылымдар және бұл пили арқsлы ДНҚ донордан (F+) рецепиентке ( F -) тасымалданады. Бұл процесс 2.13. суретінде көрсетілген. Донорлық жасуша F – факторды сақтаса, ал рецепиенттік жасуша тауып алады және F+ ацналады. Бұның процесі жай жүреді, сондықтан да F – плизмидтерге айналғанға дейін процесс тоқтайды, F – тің бұрынғы жасушасы бір немесе бірнеше рет қайталанып өзінің копиясын шығарып алады, нәтижесінде F – - тің жасушалары сақталынады.
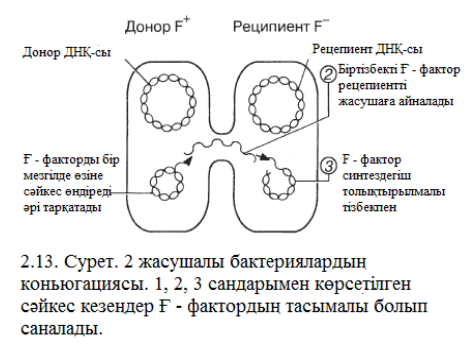
F – фактор уақыт өте келе үлкен қызығушылық танытып жүр, себебі ол 100 000 жағдайдың 1-уінде ғана орын алатын жағдай тудырып отыр, яғни жасушалар – қожасының негізгі ДНҚ-сының молекуласында кездесетініне байланысты. Ол кезде конъюгацияда тасымал тек F – факторда ғана емес, қалған басқа ДНҚ – ларда да жүреді. Бұл процес шамамен 90 мин алады, бірақ жасушалар толық ДНҚ алмасуға қарағанда ертерек тарап кетеді. Мұндай штаммдар ДНҚ – ның үлкен немесе барлық көп бөлігін басқа жасушаларға береді. Бұл штаммдар Hfr – штаммдары (ағылшынша Н – High - биік, f – frequency – жиілік, r – recombination – рекомбинация), себебі мұндай штаммдардың донорлы ДНҚсы рецепиенттің ДНҚсына қайта әрекет етеді.
2. 3. 4. Қоректену
Қоректену – энергия мен заттарды сатып алу процесі. Табиғатқа сүйеніп, энергия немесе оттек көздеріне керек –дамуға ең маңызды элемент, - тірі ағзаларды бірнеше топтарға бөлуге болады. Органикалық байланыстардың синтезі үшін тірі ағзалар тек энергияның 2 типін қолдануға ғана қабілеттті болып келеді, олар: жарық энергиясы және химиялық байланыстар энергиясы. Жарық энергиясын қолданатын ағзалар фототрофтар деп аталады, ал химиялық энергияны пайдаланатын ағзалар хемотрофтар деп аталады. Фототрофтар фотосинтез жасап шығарады.
Айтқандай – ақ, организмдер тағы автотрофты және гетеротрофты бөлінеді, ол – олардың қандай оттек көзін пайдаланатынына байланысты болып келеді, бейорганикалық байланыс (оттек диоксиді) және әртүрлі органикалық заттар. Олай болса, қоректену түрлерін 4 – ке бөлуге болады (2.3. кестесі). Бактериялар арасында берілген 4 типтің барлығы кездеседі. Топтың көшін басын бастап тұрған хемогетеротрофты бактериялар болып табылады.
Хемогетеротрофты бактериялар
Бұл топтағы бактериялар химиялық байланысты өнімнен түскен энергияны алады. Олар әртүрлі заттардың үлкен бөлігін қолдануға қабілетті болып келеді. Хемогетеротрофты бактериялар арасында 3 негізгі топты белгілеп айтуға болады, яғни сапротофтар, мутуалисттер және паразиттер.
Мутуализм (немесе симбиозом) деп 2 ағза арасындағы, яғни 2 ағзаға да тиімді болып келетін тығыз өзара байланыстың кез келген формасын атайды. Мутуализмді бактерияның мысалы ретінде Rhizobium бола алады, ол – азотты бекітуге қабілетті болып келетін және бұршақты өсімдіктерді түйнекшенің тамырында өседі, мысалы: бұршақ және жоңышқа, немесе Escherichia coli, ол адам ішектерінде мекендейді, және бір таңғалатыны, адамға В және К топтарының витаминдерімен қамтамасыз етеді.
Паразит деп терінің ішінде немесе басқа бір дененің (иесінің) бойында өсетін, одан қорегін алады да қашып кететін кез келген ағза болып табылады. Иелері әртүрлі түрдің өкілдері бола алады, әрі паразиттер иелеріне елеулізиян әкеледі. Патогендер деп ауруға әкеліп соқтыратын паразиттер деп аталады. Оның кейбіреулері 2.10. суретінде көрсетілген және 15ші тарауда қарастырылады. Және тағы бір басқа паразиттер бар, олар тек қана тірі жасушада ғана өмір сүре алатын және сол жақта өсетін паразиттер, оларды анаэробты ағзалар деп атайды. Ал факультативті деп аталатын паразиттер – иелеріне жұқтырып, оған бүлінуді шақыралы және содан кейін оның қалдықтарында сапротрофтар сияқты өмір сүреді. Паразиттер өнімде арнайы талғампаздық пен ерекшеленеді, сондықтан олар даму факторлары жөніндегі қосалқы көмекке мұқтаж, яғни олар өздігінен синтездей алмайды, бірақ иелерінен ала алады.
Фотоавтотрофты бактериялар
Фотоавтотрофты бактерияларға мысал ретінде цианобактериялар, және көк – жасыл бактериялары жатады. Балдырлар және өсімдіктер фотоавтотрофты бола алады. Олардың бәрі фотосинтезді жүзеге асырады және жалғыз көміртек көзі ретінде көмірқышқыл газын (СО2) пайдаланады (2.3. кесте). Фотосинтез процесі алғаш рет бактерияларда пайда болды, атап айтқанда цианобактерияларда. Әрі қарай көретініміздей, балдыр хлоропласттары және жер бетіндегі өсімдіктер бір кездерде өзінің уақытында гетеретрофты жасушаларда орналасқан және еркін өмір сүретін фотосинтезді бактериялардың тұқымы болып есептелінеді (2.6.1. бөлігінде). Цианобактериялар теңіздің жоғарғы қабатында және тұщы су қоймаларында кең таралған. Одан басқа, олар көлеңкелі жерлерде, құздарда, балшықтарда, сүректерде (ағаштың қабық астындағы қатты бөлігі) және кейбір тірі ағзаларда жастықтәрізді шырышты жаратылыстарында табылған.
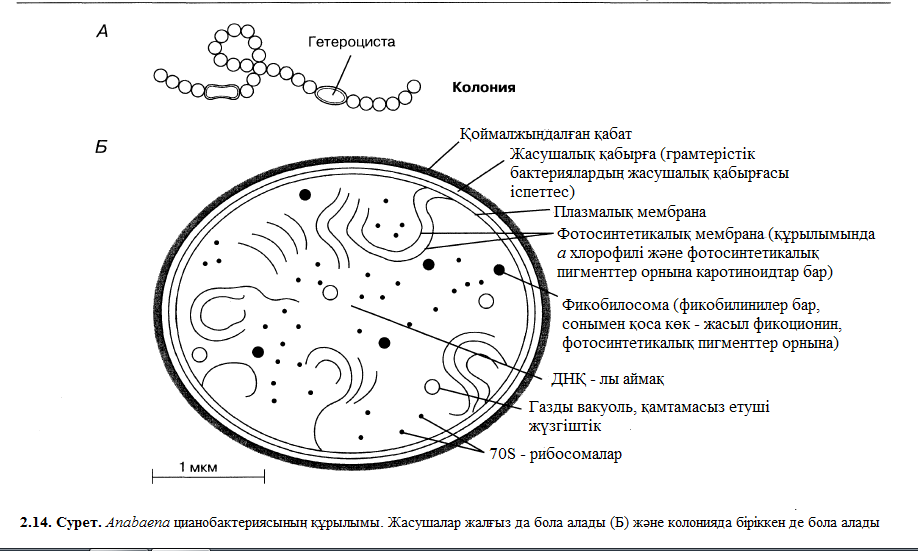
Цианобактериялардың басым көпшілігі біржасушалылар болып көрсетілген, дегенмен олардың кейбіреулері шырышты жіптерімен немес тізбектермен байланысады, мысалы, Anabaena және Spirulina. Бактериялардың басым көпшілігінен олардың айырмашылығы, балдырлар мен өсімдіктерге ұқсас келеді, фотосинтезге қабілетті, олай болса, оттекті судан шығарып алуға да қабілетті. 2.14. суретінде Anabaena ның әдеттегі құрылымы көрсетілген. Суретте көрініп тұрғандай, цитаплазма ұзындығы ерекше көрініспен фотосинтездік пигменттер орналасқан фотосинтездік мембраналарды тізеді. Пигменттер а хлорофилдері болып көрсетілген, пигменттерге және арнайы к-к жасыл пигменті – фикоцианинге өсімдіктер мен балдырлаар ұқсас болып келеді. Цианобактериялар жасушалары, әдеттегідей, өзге бактериялардан үлкенірек болып келеді. Цианобактерия қабілеті, ол фотосинтез процесі кезінде оттек бөлу, оларда фотосинтетикалық мембрана бар, және а хлорофилі цинаобактериялар толығымен калған басқа актериялар мен эукариоттапдың арасында түйіні бола алатынын дәлелдейді.
Anabaena сияқты кейбір цианобактериялар, азот бекіте алады. Басқаша айтқанда, олар ауадағы газтәрізді азотты аммиакқа айналдыра алуға қабілеті бар, сосын оны аминқышқыл, нәруыздарды және және бойында азот байланыстары бар органикалық байланыстардың синтезіне қолдануға болады. Азот жетіспеушілігінен пайда болатынн гетероцисттер мамандандырылған жасушаларда болады. Гетероцисттер өзінің бойындағы азоттық заттарды көрші жасушаларға басқа өнімдік заттарға, мысалы, көміртекке айырбастап, экспорттап отырады.
2. 3. 4. Қоректену
Қоректену – энергия мен заттарды сатып алу процесі. Табиғатқа сүйеніп, энергия немесе оттек көздеріне керек –дамуға ең маңызды элемент, - тірі ағзаларды бірнеше топтарға бөлуге болады. Органикалық байланыстардың синтезі үшін тірі ағзалар тек энергияның 2 типін қолдануға ғана қабілеттті болып келеді, олар: жарық энергиясы және химиялық байланыстар энергиясы. Жарық энергиясын қолданатын ағзалар фототрофтар деп аталады, ал химиялық энергияны пайдаланатын ағзалар хемотрофтар деп аталады. Фототрофтар фотосинтез жасап шығарады.
Айтқандай – ақ, организмдер тағы автотрофты және гетеротрофты бөлінеді, ол – олардың қандай оттек көзін пайдаланатынына байланысты болып келеді, бейорганикалық байланыс (оттек диоксиді) және әртүрлі органикалық заттар. Олай болса, қоректену түрлерін 4 – ке бөлуге болады (2.3. кестесі). Бактериялар арасында берілген 4 типтің барлығы кездеседі. Топтың көшін басын бастап тұрған хемогетеротрофты бактериялар болып табылады.
Хемогетеротрофты бактериялар
Бұл топтағы бактериялар химиялық байланысты өнімнен түскен энергияны алады. Олар әртүрлі заттардың үлкен бөлігін қолдануға қабілетті болып келеді. Хемогетеротрофты бактериялар арасында 3 негізгі топты белгілеп айтуға болады, яғни сапротофтар, мутуалисттер және паразиттер.
Мутуализм (немесе симбиозом) деп 2 ағза арасындағы, яғни 2 ағзаға да тиімді болып келетін тығыз өзара байланыстың кез келген формасын атайды. Мутуализмді бактерияның мысалы ретінде Rhizobium бола алады, ол – азотты бекітуге қабілетті болып келетін және бұршақты өсімдіктерді түйнекшенің тамырында өседі, мысалы: бұршақ және жоңышқа, немесе Escherichia coli, ол адам ішектерінде мекендейді, және бір таңғалатыны, адамға В және К топтарының витаминдерімен қамтамасыз етеді.
Паразит деп терінің ішінде немесе басқа бір дененің (иесінің) бойында өсетін, одан қорегін алады да қашып кететін кез келген ағза болып табылады. Иелері әртүрлі түрдің өкілдері бола алады, әрі паразиттер иелеріне елеулізиян әкеледі. Патогендер деп ауруға әкеліп соқтыратын паразиттер деп аталады. Оның кейбіреулері 2.10. суретінде көрсетілген және 15ші тарауда қарастырылады. Және тағы бір басқа паразиттер бар, олар тек қана тірі жасушада ғана өмір сүре алатын және сол жақта өсетін паразиттер, оларды анаэробты ағзалар деп атайды. Ал факультативті деп аталатын паразиттер – иелеріне жұқтырып, оған бүлінуді шақыралы және содан кейін оның қалдықтарында сапротрофтар сияқты өмір сүреді. Паразиттер өнімде арнайы талғампаздық пен ерекшеленеді, сондықтан олар даму факторлары жөніндегі қосалқы көмекке мұқтаж, яғни олар өздігінен синтездей алмайды, бірақ иелерінен ала алады.
Фотоавтотрофты бактериялар
Фотоавтотрофты бактерияларға мысал ретінде цианобактериялар, және көк – жасыл бактериялары жатады. Балдырлар және өсімдіктер фотоавтотрофты бола алады. Олардың бәрі фотосинтезді жүзеге асырады және жалғыз көміртек көзі ретінде көмірқышқыл газын (СО2) пайдаланады (2.3. кесте). Фотосинтез процесі алғаш рет бактерияларда пайда болды, атап айтқанда цианобактерияларда. Әрі қарай көретініміздей, балдыр хлоропласттары және жер бетіндегі өсімдіктер бір кездерде өзінің уақытында гетеретрофты жасушаларда орналасқан және еркін өмір сүретін фотосинтезді бактериялардың тұқымы болып есептелінеді (2.6.1. бөлігінде). Цианобактериялар теңіздің жоғарғы қабатында және тұщы су қоймаларында кең таралған. Одан басқа, олар көлеңкелі жерлерде, құздарда, балшықтарда, сүректерде (ағаштың қабық астындағы қатты бөлігі) және кейбір тірі ағзаларда жастықтәрізді шырышты жаратылыстарында табылған.
Цианобактериялардың басым көпшілігі біржасушалылар болып көрсетілген, дегенмен олардың кейбіреулері шырышты жіптерімен немес тізбектермен байланысады, мысалы, Anabaena және Spirulina. Бактериялардың басым көпшілігінен олардың айырмашылығы, балдырлар мен өсімдіктерге ұқсас келеді, фотосинтезге қабілетті, олай болса, оттекті судан шығарып алуға да қабілетті. 2.14. суретінде Anabaena ның әдеттегі құрылымы көрсетілген. Суретте көрініп тұрғандай, цитаплазма ұзындығы ерекше көрініспен фотосинтездік пигменттер орналасқан фотосинтездік мембраналарды тізеді. Пигменттер а хлорофилдері болып көрсетілген, пигменттерге және арнайы к-к жасыл пигменті – фикоцианинге өсімдіктер мен балдырлаар ұқсас болып келеді. Цианобактериялар жасушалары, әдеттегідей, өзге бактериялардан үлкенірек болып келеді. Цианобактерия қабілеті, ол фотосинтез процесі кезінде оттек бөлу, оларда фотосинтетикалық мембрана бар, және а хлорофилі цинаобактериялар толығымен калған басқа актериялар мен эукариоттапдың арасында түйіні бола алатынын дәлелдейді.
Anabaena сияқты кейбір цианобактериялар, азот бекіте алады. Басқаша айтқанда, олар ауадағы газтәрізді азотты аммиакқа айналдыра алуға қабілеті бар, сосын оны аминқышқыл, нәруыздарды және және бойында азот байланыстары бар органикалық байланыстардың синтезіне қолдануға болады. Азот жетіспеушілігінен пайда болатынн гетероцисттер мамандандырылған жасушаларда болады. Гетероцисттер өзінің бойындағы азоттық заттарды көрші жасушаларға басқа өнімдік заттарға, мысалы, көміртекке айырбастап, экспорттап отырады.

Хемоавтотрофты бактериялар
Бұл ағзалар әдетте хемосинтездегіштер деп аталады. Олар СО2 -ны (көміртегі диоксиді) көміртек көзі ретінде пайдаланады, бірақ энергияны химиялық реакциялар нәтижесінде алады. Қажет энергияның босатылуы бейорганикалық аттарды, мысалы, аммиак пен нитраттарды тотықсыздандыруда болады. Кейбір хемоавтотрофты бактериялар нитрификация процесіне қатысып, азот айналымында маңызды рөл атқаады. Нитрификация 2 сатыдан өтеді. Бірінші кезеңде, аммиак нитритқа дейін тотықсыздандырылады, энергия босатылулы тіркейді. Бұл кезең Nitrosomonas сияқты бактериялармен жүзеге асады. Ал екінші кезеңде түзілген нитрит нитратқа дейін толыққанды энергияның босатылуымен тотықсыздандырылады. Бұл кезең жүзеге асады, мысалы, Nitrobacter
Бұл ағзалар әдетте хемосинтездегіштер деп аталады. Олар СО2 -ны (көміртегі диоксиді) көміртек көзі ретінде пайдаланады, бірақ энергияны химиялық реакциялар нәтижесінде алады. Қажет энергияның босатылуы бейорганикалық аттарды, мысалы, аммиак пен нитраттарды тотықсыздандыруда болады. Кейбір хемоавтотрофты бактериялар нитрификация процесіне қатысып, азот айналымында маңызды рөл атқаады. Нитрификация 2 сатыдан өтеді. Бірінші кезеңде, аммиак нитритқа дейін тотықсыздандырылады, энергия босатылулы тіркейді. Бұл кезең Nitrosomonas сияқты бактериялармен жүзеге асады. Ал екінші кезеңде түзілген нитрит нитратқа дейін толыққанды энергияның босатылуымен тотықсыздандырылады. Бұл кезең жүзеге асады, мысалы, Nitrobacter
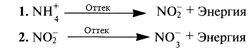
Нитрификация ұғымы азот айналымы туралы түсіндіретін 10.4.1. бөлімінде түсіндіріледі.
2.3.5. Бактерия популяциясының дамуы
2.1. Жалғыз бактерия жасушасы қашан өнімдік ортаға орналасатыны және қашан қолайлы даму жағдайында табылатыны туралы қарастырамыз. 2.4. кестесін көшіріп жазыңдар және оны толтырыңдар, бұл жасуша және оның барлық тұқымдары әрбір 20 мин сайын көбейеді екені туралы сөйлемнен тауып алып жазыңдар.
Толтырып берілген кестеге графиктер құрыңдар. Тік оське бактерия сандарын (А қисық сызығы) және бұл санның ондық дәреже көрсеткішін алып қойыңдар, ал горизонталь оське – уақытты алың. Мұндай қисық сызықтарға не айтуға болады?
2.3.5. Бактерия популяциясының дамуы
2.1. Жалғыз бактерия жасушасы қашан өнімдік ортаға орналасатыны және қашан қолайлы даму жағдайында табылатыны туралы қарастырамыз. 2.4. кестесін көшіріп жазыңдар және оны толтырыңдар, бұл жасуша және оның барлық тұқымдары әрбір 20 мин сайын көбейеді екені туралы сөйлемнен тауып алып жазыңдар.
Толтырып берілген кестеге графиктер құрыңдар. Тік оське бактерия сандарын (А қисық сызығы) және бұл санның ондық дәреже көрсеткішін алып қойыңдар, ал горизонталь оське – уақытты алың. Мұндай қисық сызықтарға не айтуға болады?
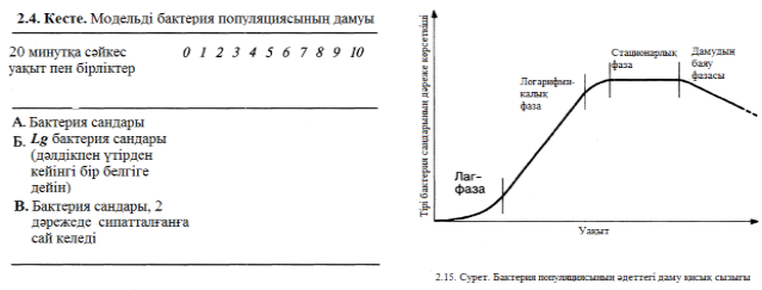
2.4. кестесінде толықтырылып көрсетілгендей, жасуша саны өскен кезде, логарифимикалық, экспоненциальдық немесе геометрикалық туралы айтылады. Бұл жағдай біз экспонциалдық қатар туралы айтамыз. Егер біз 2.4. кестесіндегі дәрежесі сай бактериялар саны 2 санымен сипатталған В қатарына қарайтын болсақ бізге түсінуге оңай болмақ. Дәреже көрсеткіші деп логарифмді немесе 2 санының экспонентін айтуға болады. Логарифмдер немесе экспоненттерді ұрпақ санымен сәйкес келетін 0, 1, 2, 3 және т.б. түзулі қатар құрайды.
2.4. кестесіне қайта оралайық: А қатарында орналасқан сандардың орнына оларды логарифмді түрде 2 ші берілген негізбен жазуға болады:
В және Г қатарларын салыстырыңыздар. Бірақ, әдеттеондық дәреже көрсеткіштерін пайдаланады (Б қатарын қарау керек). Бұл жағдайда 1 = 100, 2 =100,3, 4=100,6 және т.б.
А қатарындағы пайда болған қисық сызық (2.4. кестесі), логарифмді немесе экспоненциалды қисық сызық деп аталады. Мұндай қисық сызықты уақыт бойынша жасуша санын өзгертіп график құра отырып, тіке қайта құруға болады. Онда кемшіліксіз жағдайларда бактериялар дамуы теоретикалық түрде экспоненциалды болуы керек. Бұл математикалық модельді шынайы бактерия популяциясының дамуы туралы 2.15. суретте кекінделген қисық сызықпен салыстырып қарайық. Дамудың 4 факторы айқын бейнеленген.
2.4. кестесіне қайта оралайық: А қатарында орналасқан сандардың орнына оларды логарифмді түрде 2 ші берілген негізбен жазуға болады:
В және Г қатарларын салыстырыңыздар. Бірақ, әдеттеондық дәреже көрсеткіштерін пайдаланады (Б қатарын қарау керек). Бұл жағдайда 1 = 100, 2 =100,3, 4=100,6 және т.б.
А қатарындағы пайда болған қисық сызық (2.4. кестесі), логарифмді немесе экспоненциалды қисық сызық деп аталады. Мұндай қисық сызықты уақыт бойынша жасуша санын өзгертіп график құра отырып, тіке қайта құруға болады. Онда кемшіліксіз жағдайларда бактериялар дамуы теоретикалық түрде экспоненциалды болуы керек. Бұл математикалық модельді шынайы бактерия популяциясының дамуы туралы 2.15. суретте кекінделген қисық сызықпен салыстырып қарайық. Дамудың 4 факторы айқын бейнеленген.
- Лаг – фаза кезінде бактериялар мекендеудің аңа ортасына бейімделеді, сондықтан да даму максималды жылдамдыққа жетпейді. Бұл периодта, бактериялар жаңа ферменттерді синтездеуі мүмкін және жаңа ортада бар өдері қажетсінетін қорек заттарын сіңіруі мүмкін.
- Логарифмикалық фаза – бұл фазада бактериялар максималды жылдамдыққа дейін өседі, жасуша саны жайлап экспоненциалды бола бастайды, ал қисық сызық дамуы түзу жолға кәдімгідей түседі.
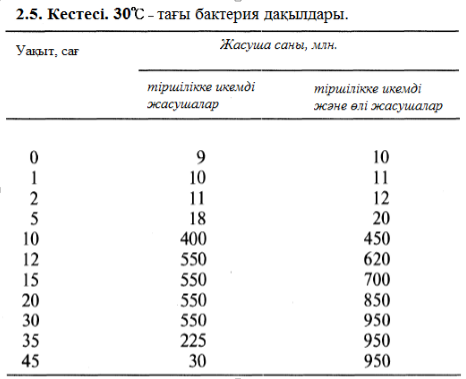
жасушалардың саны тұрақты болып қалады (2.5. ксетесі). Бұл фазаға өті ушін бірнеше факторлар арқылы қамтамасыз етілген: ортадағы қоректің өнімдердің концентрациясы төмендесе, метоболизмнің токсинді өнімдерінің жиналуы, ал аэробты бактериялар жағдайы кезінде, оттектің ортада азаюы болып саналады.
4. Соңғы фаза – дамудың баяулау фазасы кезінде – жасушалардың қырылуы көбейеді және олардың көбеюлер құртылады. Жасушаларды санап үйрену қабілеттері 12 бөлімнің соңында жазылған.
4. Соңғы фаза – дамудың баяулау фазасы кезінде – жасушалардың қырылуы көбейеді және олардың көбеюлер құртылады. Жасушаларды санап үйрену қабілеттері 12 бөлімнің соңында жазылған.
2.4. Вирустар
2.4.1. Ашылымы
1852 жылы орыс ботанигі Д. И. Ивановский алғашқы рет мозаикалық аурумен жеңілген табак өсімдігінен инфекционды эстракт алды. Мұндай эстрактты тұқтыр бактериялар фильтрі арқылы жүргізген кезде, сүзілген сұйықтық инфекционды құрылымды сақтап тұрды. 1898 жылы голлондиялық Бейернек «вирус» (латыннан уадарғанда «у» деген мағынаны береді) деген жаңа сөз ойлап шықты, бұл сөзді өсімдік сұйыөтыөтарымен сүзіліп анықталып, инфекционды табиғатқа байланысты қолданады. Әйтсе де, жоғары деңгейде тазартылған вирус сынамасы сияқты көптеген үлкен дәрежелі жеңістіктерге жетуге мүмкіндік туды мен химиялық табиғатта нуклеопротеиндер (нәруыздар мен нуклеинді қышқылдардан тұратын күрделі байланыстар) бөліктерінің өздері әлі уақытқа дейін қолға түспейтін және жұмбақ болып қалмақ, себебі оларды жарық микроскобынан көру үшін өте аз болатын. Және дәл осы үшін вирустар алғашқы биологилық құрылым болып саналады, себеі олар олардың пайда болысымен ХХғасырда 30 шы жылдарда электронды микроскоппен алғаш зерттелген.
2.4.2. Вирустар құрылымы
Вирустар мынадай құрылымдары иемденеді:
Өлшемдері
Вирустар – ұсақ тірі ағзалар, олардың өлшемдері құбылтып алғанда шамамен 20дан 300 нм ге дейін алады, орташа алғанда олар бір 50 бактерия. Оларды жарық микроскобынан өкру мүмкін емес, олра фильтр арқылы өтеі, өткізбейтін бактериялар.
Шығу тарихы
Зерттеушілер «вирустар тірі ме өзі?» деген сұрақты жиі қояды. Егер кез келген тірі ағзада генетекилық материал (ДНҚ немесе РНҚ) болатын болса және өздігінен туу қабілеті болса, онда вирустар – тірі деген жағымды жауапты кесіп айтуға болады. Егер тірі ағзаны жасушалық құрылысына қарайтын болсақ, онда вирустар – өлі деген теріс жауап аламыз. осыған байланысты, жасушалардың сырты – вирустар иесі өздігінен тууға қабілетсіз.
Вирустар туралы толық көрсетілімі үшін онығ пайда болудағы эволюциясына қарау керек. Вирустар – ешқашан прокариоттық және эукариоттық жасушалардан «жоқ болып кетпейтін» және жасушалық ортасына келгенде де жаңғырту қабілетін сақтап қалатын деген дәлелді болмас да, болжам бар. Жасушаның сыртында вирустар толығымен инертті (жігерсіз) күйде болады, бірақ оларда инструкция жинақтары (генетикалық код) бар, ол өзінің инструкциясына боұсындыра отырып, жасушаың ішіне өтіп кете алады, және бұл өзіне ұқсас вирустарды өндіріп алады. Демек, логикалық түрде болжап қарасақ, эволюция кезінде вирустар жасушадан кейін пайда болған.
Құрылысы
Вирустардың құрылысы өте қарапайым. Олар келесі құрылыстардан тұрады.:
Капсидтер құрылысы үшін симметрия типтері тән, әсіресе полиэдрикалық және шиыршықталған. Полиэдр дегеніміз – көпқырлы дене. Вирустардың полиэдрикалық формаларының көп таралған түрі – икосаэдр, ол 20 үшбұрышты қырлардан, 12 бұрыштан және 30 қабырғадан тұрады. 2.17. А суретіндебіз дұрыс икосаэдрді көреміз, ал 2.17. Б суретінде –икосаэдрдағы 162 капсомердің бөлшегіндегі ұшық вирусын көреміз.
Көрнекі иллюстрациядағы шиыршықты симметриялар 2.18. Б суретінде РНҚ – темекі мозаикасында болатын вирус (ТМТ)ретінде қызмет етеді. Бұл вирустың капсиді 2130 ұқсас нәруызды капсомерлерден тұрады. ТМТ ол таза күйінде белгіоенген ең алғашқы вирус. Бұл вируспен ауру өсімдік жапырақшасын зақымдау кезінде сары дақтар пайда болады – бұлар жапырақ мозаикалар деп аталады (2.18, В сурет). Ауру өсімдіктер немесе оның бөліктері сау өсімдіктермен жалғасқанда, не болмаса дайындау үшін арнайы зақымдалған өсімдікті пайдаланып жасалған темекі түтіні арқылы тыныс алу жолымен түскен вирустар механикалық түрде өте тез таралады.
Бактерияларға шабуылдайтын вирустар бактериофаг немесе жай ғана фагтар тобын құрайды. Кейбір бактерифагтарда шиыршықталған симметриялы анық кескінделген икосаэдрикалық бас пен құйрық бар (2.19. сурет). 2.20 немесе 2.21. суреттерде жалпылама құрыстарымен өлшемдеріне байланысты кейбір вирустарды бейнелеген.
2.4.1. Ашылымы
1852 жылы орыс ботанигі Д. И. Ивановский алғашқы рет мозаикалық аурумен жеңілген табак өсімдігінен инфекционды эстракт алды. Мұндай эстрактты тұқтыр бактериялар фильтрі арқылы жүргізген кезде, сүзілген сұйықтық инфекционды құрылымды сақтап тұрды. 1898 жылы голлондиялық Бейернек «вирус» (латыннан уадарғанда «у» деген мағынаны береді) деген жаңа сөз ойлап шықты, бұл сөзді өсімдік сұйыөтыөтарымен сүзіліп анықталып, инфекционды табиғатқа байланысты қолданады. Әйтсе де, жоғары деңгейде тазартылған вирус сынамасы сияқты көптеген үлкен дәрежелі жеңістіктерге жетуге мүмкіндік туды мен химиялық табиғатта нуклеопротеиндер (нәруыздар мен нуклеинді қышқылдардан тұратын күрделі байланыстар) бөліктерінің өздері әлі уақытқа дейін қолға түспейтін және жұмбақ болып қалмақ, себебі оларды жарық микроскобынан көру үшін өте аз болатын. Және дәл осы үшін вирустар алғашқы биологилық құрылым болып саналады, себеі олар олардың пайда болысымен ХХғасырда 30 шы жылдарда электронды микроскоппен алғаш зерттелген.
2.4.2. Вирустар құрылымы
Вирустар мынадай құрылымдары иемденеді:
- Бүлар ұсақ ағзалар.
- Оларда жасушалық құрылым болмайды.
- Вирустар тірі жасушаға ену арқылы өздерін жаңғыртуға қабілетті. Демек, олардың бәрі – облигатты эндопаразиттер. Басқа сөзбен айтұанда, вирустар тек қана өзге жасушалардық ішінде өмір сүре алады. Оның басым көпшілігі ауру тудырады.
- Вирустар жай ғана өте үш еселенген. Олар үлкен емес нукленинді қышқылдардың молекуларынан, нәруызды немесе липопротеинді не ДНҚ, не РНҚ сияқты қабықтардан тұрады.
- Олар тірі не болмаса өлілердің шегінде мекен етеді.
- Вирустар тек қана нақты жасуша типтерін айыра біледі және жұқтыра алады. Басқа сөзбен айтқанда, вирустар өздерінің иелерімен ара қатынасы өте үлкен арнайы байланыс болып саналады.
Өлшемдері
Вирустар – ұсақ тірі ағзалар, олардың өлшемдері құбылтып алғанда шамамен 20дан 300 нм ге дейін алады, орташа алғанда олар бір 50 бактерия. Оларды жарық микроскобынан өкру мүмкін емес, олра фильтр арқылы өтеі, өткізбейтін бактериялар.
Шығу тарихы
Зерттеушілер «вирустар тірі ме өзі?» деген сұрақты жиі қояды. Егер кез келген тірі ағзада генетекилық материал (ДНҚ немесе РНҚ) болатын болса және өздігінен туу қабілеті болса, онда вирустар – тірі деген жағымды жауапты кесіп айтуға болады. Егер тірі ағзаны жасушалық құрылысына қарайтын болсақ, онда вирустар – өлі деген теріс жауап аламыз. осыған байланысты, жасушалардың сырты – вирустар иесі өздігінен тууға қабілетсіз.
Вирустар туралы толық көрсетілімі үшін онығ пайда болудағы эволюциясына қарау керек. Вирустар – ешқашан прокариоттық және эукариоттық жасушалардан «жоқ болып кетпейтін» және жасушалық ортасына келгенде де жаңғырту қабілетін сақтап қалатын деген дәлелді болмас да, болжам бар. Жасушаның сыртында вирустар толығымен инертті (жігерсіз) күйде болады, бірақ оларда инструкция жинақтары (генетикалық код) бар, ол өзінің инструкциясына боұсындыра отырып, жасушаың ішіне өтіп кете алады, және бұл өзіне ұқсас вирустарды өндіріп алады. Демек, логикалық түрде болжап қарасақ, эволюция кезінде вирустар жасушадан кейін пайда болған.
Құрылысы
Вирустардың құрылысы өте қарапайым. Олар келесі құрылыстардан тұрады.:
- Өзектер – не ДНҚ, не болмаса РНҚ арқылы көрсетілген генетикалық матреиал; ДНҚ немесе РНҚ біртізбіекті немесе қостізбекті бола алады;
- Капсида – өзекті өоршап тұратын нәруызды қабаттарын қорғайды;
- Нуклеокапсида –капсид пен өзектен тұратын күрделі құрылым;
- Қабаттар – ВИЧ немесе грипп вирустары иесінің – жасущасының плазмалық мембранасынан тұратын липопротеинді қабат құрайды;
- Капсомерлер – капсидтер салынғандардан болатын қайталанатын ұқсас суббірлік.
Капсидтер құрылысы үшін симметрия типтері тән, әсіресе полиэдрикалық және шиыршықталған. Полиэдр дегеніміз – көпқырлы дене. Вирустардың полиэдрикалық формаларының көп таралған түрі – икосаэдр, ол 20 үшбұрышты қырлардан, 12 бұрыштан және 30 қабырғадан тұрады. 2.17. А суретіндебіз дұрыс икосаэдрді көреміз, ал 2.17. Б суретінде –икосаэдрдағы 162 капсомердің бөлшегіндегі ұшық вирусын көреміз.
Көрнекі иллюстрациядағы шиыршықты симметриялар 2.18. Б суретінде РНҚ – темекі мозаикасында болатын вирус (ТМТ)ретінде қызмет етеді. Бұл вирустың капсиді 2130 ұқсас нәруызды капсомерлерден тұрады. ТМТ ол таза күйінде белгіоенген ең алғашқы вирус. Бұл вируспен ауру өсімдік жапырақшасын зақымдау кезінде сары дақтар пайда болады – бұлар жапырақ мозаикалар деп аталады (2.18, В сурет). Ауру өсімдіктер немесе оның бөліктері сау өсімдіктермен жалғасқанда, не болмаса дайындау үшін арнайы зақымдалған өсімдікті пайдаланып жасалған темекі түтіні арқылы тыныс алу жолымен түскен вирустар механикалық түрде өте тез таралады.
Бактерияларға шабуылдайтын вирустар бактериофаг немесе жай ғана фагтар тобын құрайды. Кейбір бактерифагтарда шиыршықталған симметриялы анық кескінделген икосаэдрикалық бас пен құйрық бар (2.19. сурет). 2.20 немесе 2.21. суреттерде жалпылама құрыстарымен өлшемдеріне байланысты кейбір вирустарды бейнелеген.
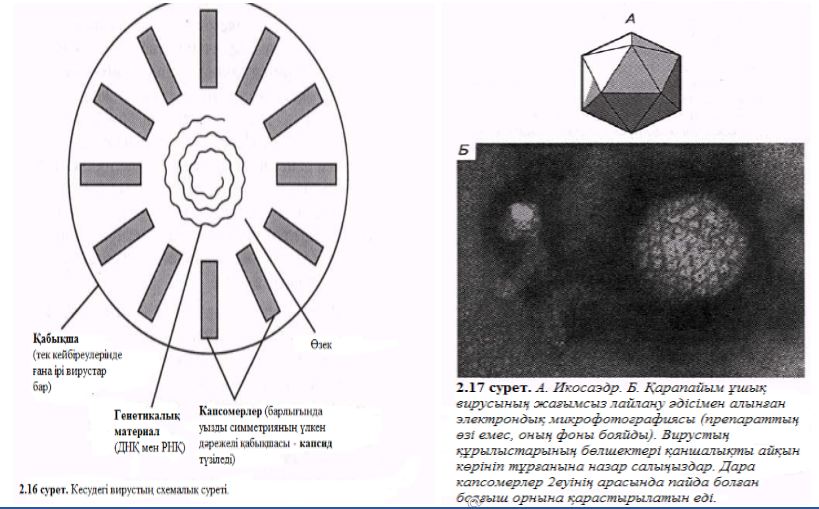
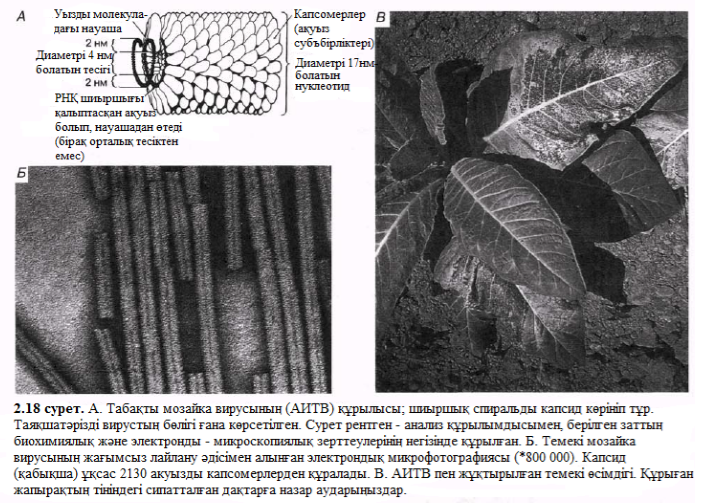
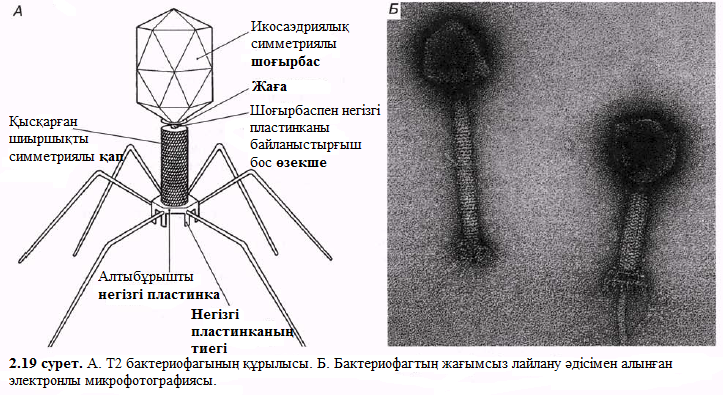
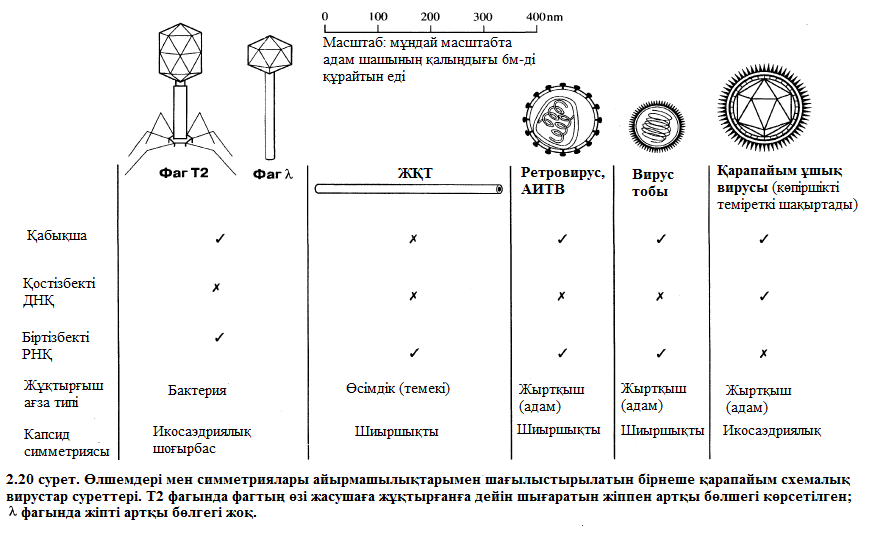
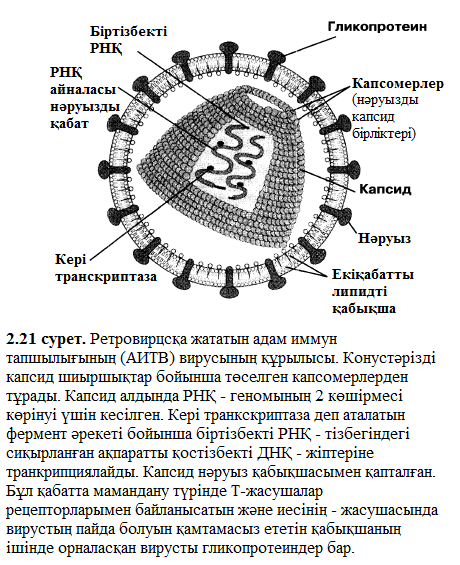
2.4.3. Бактериофагтың тіршілік кезеңі
2.22. суретінде әдеттегі бактериофагтың тіршілік кезеңі берілген. E.Coli, әдеттегі иесінің жасушасы болып табылады, ол Т – фагтарға аздап шабуылдауы мүмкін (Т1 – ден Т7 – ге дейін). Т – анық фаг болып табылады (мысалы, Т2 фаг), ол 2.19, А және Б және 2.20.
2.22. суретінде әдеттегі бактериофагтың тіршілік кезеңі берілген. E.Coli, әдеттегі иесінің жасушасы болып табылады, ол Т – фагтарға аздап шабуылдауы мүмкін (Т1 – ден Т7 – ге дейін). Т – анық фаг болып табылады (мысалы, Т2 фаг), ол 2.19, А және Б және 2.20.
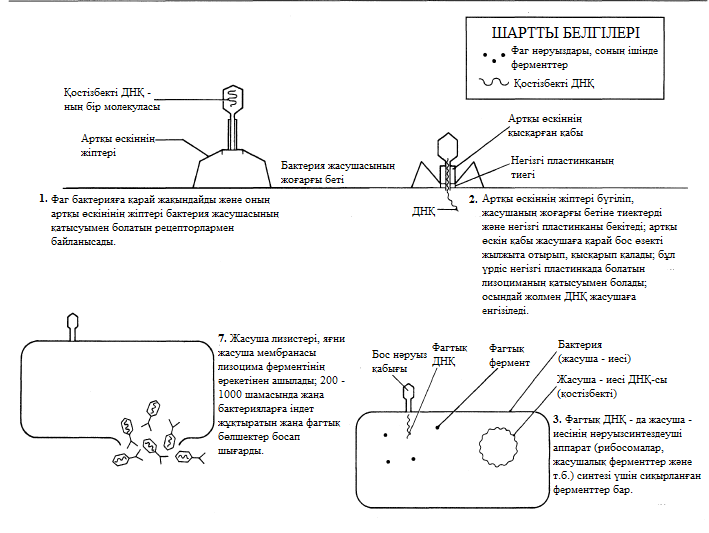
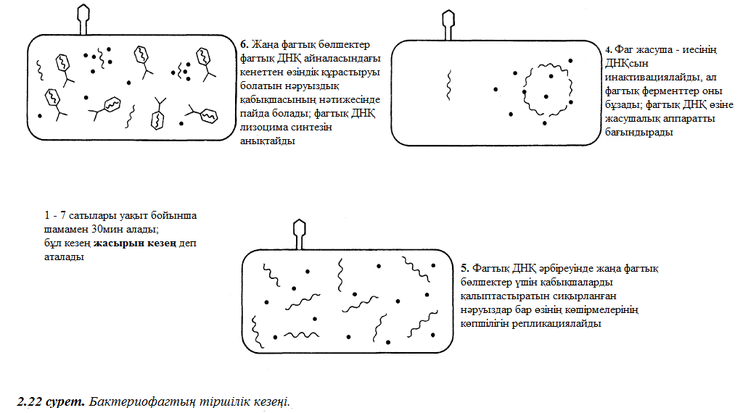
Фагтардың басым көпшілігінің тіршілік кезеңдері ұқсас болып келеді. Бірақ олардың ішінде біреуінің тіршілік кезеңі үздіксіз жүреді; бұндай кезде дамудың логикалық кезеңі жүреді. Басқа фагтарда, лямбда сияқты фагтық ДНҚ, жасушада көрініп, иесінің жасушасының кіреді және көптеген ұрпақтарда ешқандай пайда болмайды. Әрбір бөлінуде ДНҚ жасушалары жасушалы ДНҚмен көшіріледі. Мұндай белсенді емес фаг профаг деп аталады. бірақ, белгілі ьір уақытта профаг белсендіріледі, яғни жасушалы ДНҚдан босатылады және иесінің жасушасының әдеттегі жолын қырылта отырып, өзінің тіршілік кезеңін тоқтатады. Мұндай фактор лизогендіктер деп аталады, оның профагымен құрылғандар – лизогендер деп аталады.
2.4.4. Вирустар ауру тудырғыштар
Вирустар тағы эукариоттық жарақаттауға қабілеттті, бұл жерде прокариоттық жасушалардыкіндей жағдай болады, әрбір вирустың өзіндік арнайы иесі болады. ТТМ мысалы, темекі өсімдігіне ғана індет жұқтырапды. Барлық вирустар өімдіктерде, жануарларда және саңырауқұлақтарда көптеген ауру туғызады. Ал адамдардың вирустық ауруларына қызылша, қызамық, шешек, тұмау, ұшық және ЖИТС жатады.
Вирустар, шамамен, басқа барлық ағзаға ауру туғызады.
2.4.5. АИТВ мысалында келтірілген ретровирустыңқұрылысы мен тіршілік кезең
Вирустық аурулардың арасынлдағы ерекше қызығушылық ЖИТСқа түседі, сондықтан ол жаңа ауру болып табылады. Бұл ауру туралы ең алғаш 1981 жылы АҚШта белгілі болды. ЖИТС – адам бойындағы иммун тапшылық вирусы болып саналады. Немесе қысқартылып АИТВ болады. Бұл вирусқа деген қызығушылық былай түсіндіріледі:
2.4.4. Вирустар ауру тудырғыштар
Вирустар тағы эукариоттық жарақаттауға қабілеттті, бұл жерде прокариоттық жасушалардыкіндей жағдай болады, әрбір вирустың өзіндік арнайы иесі болады. ТТМ мысалы, темекі өсімдігіне ғана індет жұқтырапды. Барлық вирустар өімдіктерде, жануарларда және саңырауқұлақтарда көптеген ауру туғызады. Ал адамдардың вирустық ауруларына қызылша, қызамық, шешек, тұмау, ұшық және ЖИТС жатады.
Вирустар, шамамен, басқа барлық ағзаға ауру туғызады.
2.4.5. АИТВ мысалында келтірілген ретровирустыңқұрылысы мен тіршілік кезең
Вирустық аурулардың арасынлдағы ерекше қызығушылық ЖИТСқа түседі, сондықтан ол жаңа ауру болып табылады. Бұл ауру туралы ең алғаш 1981 жылы АҚШта белгілі болды. ЖИТС – адам бойындағы иммун тапшылық вирусы болып саналады. Немесе қысқартылып АИТВ болады. Бұл вирусқа деген қызығушылық былай түсіндіріледі:
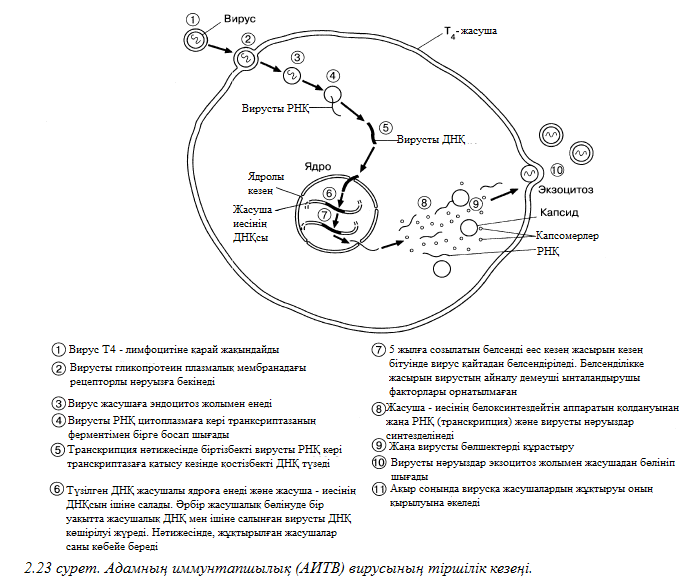
АИТВ ретровирустар деген топқа кіргізіледі, ретровирустар – келесі вирустың ерекшелігін шағылыстыраптын атау. Әдетте, генетикалық ақпараттарды тасымалдау ДНҚ РНҚға сияқты бағытпен жүреді, яғни ДНҚ – ның (генде) бір бөлігіндегі сиқырланған ақпараттарды транскипциялайды, яғни сәйкес РНҚның түзілуі мен салыстырып оқылады. Ретровирустарда генетикалық материалдың мұралығына РНҚ қызмет етеді, кері транскрипция жүреді, яғни генетикалық ақпараттарды кері салыстырып оқиды: РНҚдан ДНҚға. Кері салыстырып оқудағы ферментті кері транскриптаза деп аталады. Ол генетикалық инженерияда кеңінен қолданылады (25 бөлім).
2.21. суретінде АИТВ ның генетикалық құрылымы көрсетілген, ал 2.23. суретінде оның тіршілік кезеңі көрсетілген (тағы 2.24. суреті көрсетілген). АИТВ – т – хелпер лимфоцит атты топтағы лейкоциттерді бұзады және індет жұқтырады, нәтижесінде иммундық жүйесіндегі белсенділікті басып тастайды. Бұл вирустың шақыратын ауруын (ЖИТС) 15бөлімінде қарастырылады.
2.5. Саңырауқұлақтар патшалығы
Саңырауқұлақтар үлкен жіне сәтті дамитын ағзалар тобы болып саналады. Ол қатарфына шамамен 80000 ұқсас түрді қосады. Олардың өлшемдері ауытқымалы болып келеді, біржасушалы ашытқыдан үлкен улы саңырауқұлақтарға, жаңбыршыға және қауызға дейін жетеді. Саңырауқұлақтар ең әртүрлі мекендейтін жерлерді алады – ол суда жіне құрғақта.
Олардың одан басқа маңызды рөлдері бар, олар биосферада, медицинада адамқажеттіліне және шаруашылықта саңырауқұлақтарға сансыз өңездер тән, ол органикалық материалдарда ол (нанда, тері, аздыратын өсімдіктерде немесе өлген балықта), піскен жемістің қантты бетінде көптеп пайда болатын біржасушалы ашытқыларда және басқа да көптеген өсімдік паразиттері. Соңғысы қауқпті аурулар тудырады, яғни ақұнтақ, қара күйе және тот (көк) сияқты. Кейбір саңырауқұлақтар жануарларда паразиттейді, бірақ бұл қарым – қатынас бактерияларға қарағанда сәл қауіптілеу.
Саңырауқұлақтарды зерттейтін ғылым микология деп аталады (грекше mykes саңырауқұлақ). Ол өзін микробиологияның бір бұтағы ретінде көрсетеді, сондықтан саңырауқұлақтарды зерттеуде қолданылатын методиканың үлкен бөлігі, мысалы, зарарсыздандыру, дақылдау және т.б. дақылдау қабілеттері, із жүзінде бактерияны зерттеу барысындағы тәсілден ерекшелінбейді.
2.5.1. Саңырауқлақтардың систематикасы мен негізгі белгілері
2.2 бөлігінде айтып кеткендей, саңырауқұлақтар хлорофилі жоқ эукариоттар, демек, өлі жануарлар сияқты гетеротрофты да болып саналады. Бірақ саңырауқұлақтарда қатты жасушалық қабырға бар және олар өсімдік сияқты қозғалмайды. Олар бұрын өсімдіктерге жатқызылатын, бірақ қазір оларды бөлек патшалыққа жатқызылады (2.4 сурет).
2.5.2. Құрылысы
Саңырауқұлақтардың дене құрылысы ірегей болып келеді. Ол шоғырлар (жекеше түрде – шоғыр) деп аталатын кішігірім бұтақты түтікшелі жіпті массадан тұрады, ал бұл барлығы жиналып мицелий құрайды. Әрбір шоғыр кішігірім қатты қабырғамен қоршалады, негізгі компонентті азотты
2.6. кестесінде олардың систематикасы мен негізгі белгілері көрсетілген.
2.21. суретінде АИТВ ның генетикалық құрылымы көрсетілген, ал 2.23. суретінде оның тіршілік кезеңі көрсетілген (тағы 2.24. суреті көрсетілген). АИТВ – т – хелпер лимфоцит атты топтағы лейкоциттерді бұзады және індет жұқтырады, нәтижесінде иммундық жүйесіндегі белсенділікті басып тастайды. Бұл вирустың шақыратын ауруын (ЖИТС) 15бөлімінде қарастырылады.
2.5. Саңырауқұлақтар патшалығы
Саңырауқұлақтар үлкен жіне сәтті дамитын ағзалар тобы болып саналады. Ол қатарфына шамамен 80000 ұқсас түрді қосады. Олардың өлшемдері ауытқымалы болып келеді, біржасушалы ашытқыдан үлкен улы саңырауқұлақтарға, жаңбыршыға және қауызға дейін жетеді. Саңырауқұлақтар ең әртүрлі мекендейтін жерлерді алады – ол суда жіне құрғақта.
Олардың одан басқа маңызды рөлдері бар, олар биосферада, медицинада адамқажеттіліне және шаруашылықта саңырауқұлақтарға сансыз өңездер тән, ол органикалық материалдарда ол (нанда, тері, аздыратын өсімдіктерде немесе өлген балықта), піскен жемістің қантты бетінде көптеп пайда болатын біржасушалы ашытқыларда және басқа да көптеген өсімдік паразиттері. Соңғысы қауқпті аурулар тудырады, яғни ақұнтақ, қара күйе және тот (көк) сияқты. Кейбір саңырауқұлақтар жануарларда паразиттейді, бірақ бұл қарым – қатынас бактерияларға қарағанда сәл қауіптілеу.
Саңырауқұлақтарды зерттейтін ғылым микология деп аталады (грекше mykes саңырауқұлақ). Ол өзін микробиологияның бір бұтағы ретінде көрсетеді, сондықтан саңырауқұлақтарды зерттеуде қолданылатын методиканың үлкен бөлігі, мысалы, зарарсыздандыру, дақылдау және т.б. дақылдау қабілеттері, із жүзінде бактерияны зерттеу барысындағы тәсілден ерекшелінбейді.
2.5.1. Саңырауқлақтардың систематикасы мен негізгі белгілері
2.2 бөлігінде айтып кеткендей, саңырауқұлақтар хлорофилі жоқ эукариоттар, демек, өлі жануарлар сияқты гетеротрофты да болып саналады. Бірақ саңырауқұлақтарда қатты жасушалық қабырға бар және олар өсімдік сияқты қозғалмайды. Олар бұрын өсімдіктерге жатқызылатын, бірақ қазір оларды бөлек патшалыққа жатқызылады (2.4 сурет).
2.5.2. Құрылысы
Саңырауқұлақтардың дене құрылысы ірегей болып келеді. Ол шоғырлар (жекеше түрде – шоғыр) деп аталатын кішігірім бұтақты түтікшелі жіпті массадан тұрады, ал бұл барлығы жиналып мицелий құрайды. Әрбір шоғыр кішігірім қатты қабырғамен қоршалады, негізгі компонентті азотты
2.6. кестесінде олардың систематикасы мен негізгі белгілері көрсетілген.

полисахарид – хитин болып табылады. Хитин тағы да буын аяқтылардың құрылымдық компонеті болып табылады (2.8.6 бөлім). Шоғырларда шынайы нағыз жасушалық құрылым жоқ. Пртоплазма гифі не мүлдем бөлінбейді, не септа дейтін көлденең аралықтарға бөлінеді. Септер ішінлегі шоғырдың ұсқыны жасушаларға ұқсас бөліктерге бөледі. Септің орталығында, әдеттегінше, кішігірім тесік қалады, протоплазма арқылы бір компартменттен екіншісіне ағып өтуі мүмкін.
Шоғырдың өн бойына бір – бірінен белгілі бір аз да емес, көп те емес арақашықтықта орналасқан әрбір компартментте бір немесе екі немесе бірнеше ядролар болуы мүмкін. Аралықтары бар шоғырлар мүшелілер немесе септирокалық деп аталады, мысалы, Penicillium (2.25 сурет). Алалықтары жоқ шоғырлар беймүшелілер немесе асептирокалық деп аталады, мысалы, Mucor (2.26 сурет).
Шоғыр цитоплазмасында әдеттегі эукариот органеллалары үшін митохондрия, Гольджи аппараты, эндоплазмалық ретикулум, рибосомалар және вакуольдер бар. Ескі тарауларда мицелия вакуольдері көлемді, ал цитоплазма периферияда азғана орын алатын деп берілген болатын. Уақыт өте келе шоғырлар тығыз орналасқан құрылымдарды біріктіреді: споралар түзілетін жемісті тел. Ашытқылар біржасушалы ағзалар болғандықтан ерекше топ құрайды және сол үшін оларда құрылысы жоқ, гифке тән, мысалы, Sacharomyces (2.27 сурет).
Penicillium, Mucor және Phizopus - тар өңездер сияқты белгілі болып танылады. Бұл – кеңінен таралған сапротрофтар, яғни олар өлі органикалық материалдардың қорегіне қолданылады. Олар зерттеуде өте қолайлы, сондықтан оңай дақылданады және әдеттегі сағырауқұлақ шоғырларын құрастырады.
Pennicillium түрлері нанда және бұзылған жемісте көк, жасыл және кейде сары өңезді құрайды. Мицелий (жіпшумақ) дөңгелек кіші диаметрлі колонияларды құрайды. Шоғырлары мүшелі; ерекше колония бояуларын споралар қамтамасыз етеді (2.25, А сурет).
Pennicillium – да конидий деп аталатын спора арқылы жыныссыз көбею. Ерекше шоғырдың соңғы жақтарында түзілетін конидияларды конидия сағақтары деп атайды (2.25 сурет, Б және В). Конидий конидия сағақтары спорангийлердің қатарына кірмейді, олар қарсылыққа және пісіп жетіду кезінде еркін әлсіретіледі. Шоғыр құрылысы 2.25, Г. суретінде келтірілген. Pennicillium – ның шаруашылықтағы мәнісі 12.11.1 бөлімінде талқыланады.
Мукор белгілі өңездерді қатарға қосып, түр құрайды. Бұл саңырауқұлақ жер қыртысында көп таралған және нанда да дами түседі. Агарда дақылдау кезінде аз немесе көп дөңгелек мекендер түзіледі. Шоғырлар беймүшелік немесе молынан тармақталады (2.26, Б сурет). Спорангиеностар деп аталатын ұзын тік өсетін шоғырларда орналасқан споралар сфералық спорангийде дамиды (2.26, А және Б сурет). Мицелияның спорангиеностар көп жиналған піскен түрлері ілгекке топтанғанға ұқсайды; дәл осы үшін Mucorды бос тәрізді өңездер деп атайды. Спорангийлер үлкен емес жақындатылған микроскоптан жақсы көрінеді. Мукор тез өседі және шамамен 3 күнде 20 - та барлық Петри тостақаншасында шайылып, өсіп кетеді. Шоғырдың ішкі қуысы әдеттегі эукариот сияқты құрылымды болады, мысалы, Pennicillium (2.25, Г суреті), бір айырмашылығы мукор шоғырларында аралық жоқ. Phizopus Mucor – ға өте ұқсас болып келеді. Столондар деп аталатын кейбір шоғырларда бірнеше қиғашты формалары болады. Столондардың соңында тамырға ұқсас шағын шоғырлы байламдар құрылады, Phizopus – та бір немесе сол жерден екі немесе одан да көп Mucor – дан бөлек спорангиеностар өсіп шығады, яғни спорангиеностар бөлек өсіп шығады.
2.3. Спорангиеностар не үшін керек?
Шоғырдың өн бойына бір – бірінен белгілі бір аз да емес, көп те емес арақашықтықта орналасқан әрбір компартментте бір немесе екі немесе бірнеше ядролар болуы мүмкін. Аралықтары бар шоғырлар мүшелілер немесе септирокалық деп аталады, мысалы, Penicillium (2.25 сурет). Алалықтары жоқ шоғырлар беймүшелілер немесе асептирокалық деп аталады, мысалы, Mucor (2.26 сурет).
Шоғыр цитоплазмасында әдеттегі эукариот органеллалары үшін митохондрия, Гольджи аппараты, эндоплазмалық ретикулум, рибосомалар және вакуольдер бар. Ескі тарауларда мицелия вакуольдері көлемді, ал цитоплазма периферияда азғана орын алатын деп берілген болатын. Уақыт өте келе шоғырлар тығыз орналасқан құрылымдарды біріктіреді: споралар түзілетін жемісті тел. Ашытқылар біржасушалы ағзалар болғандықтан ерекше топ құрайды және сол үшін оларда құрылысы жоқ, гифке тән, мысалы, Sacharomyces (2.27 сурет).
Penicillium, Mucor және Phizopus - тар өңездер сияқты белгілі болып танылады. Бұл – кеңінен таралған сапротрофтар, яғни олар өлі органикалық материалдардың қорегіне қолданылады. Олар зерттеуде өте қолайлы, сондықтан оңай дақылданады және әдеттегі сағырауқұлақ шоғырларын құрастырады.
Pennicillium түрлері нанда және бұзылған жемісте көк, жасыл және кейде сары өңезді құрайды. Мицелий (жіпшумақ) дөңгелек кіші диаметрлі колонияларды құрайды. Шоғырлары мүшелі; ерекше колония бояуларын споралар қамтамасыз етеді (2.25, А сурет).
Pennicillium – да конидий деп аталатын спора арқылы жыныссыз көбею. Ерекше шоғырдың соңғы жақтарында түзілетін конидияларды конидия сағақтары деп атайды (2.25 сурет, Б және В). Конидий конидия сағақтары спорангийлердің қатарына кірмейді, олар қарсылыққа және пісіп жетіду кезінде еркін әлсіретіледі. Шоғыр құрылысы 2.25, Г. суретінде келтірілген. Pennicillium – ның шаруашылықтағы мәнісі 12.11.1 бөлімінде талқыланады.
Мукор белгілі өңездерді қатарға қосып, түр құрайды. Бұл саңырауқұлақ жер қыртысында көп таралған және нанда да дами түседі. Агарда дақылдау кезінде аз немесе көп дөңгелек мекендер түзіледі. Шоғырлар беймүшелік немесе молынан тармақталады (2.26, Б сурет). Спорангиеностар деп аталатын ұзын тік өсетін шоғырларда орналасқан споралар сфералық спорангийде дамиды (2.26, А және Б сурет). Мицелияның спорангиеностар көп жиналған піскен түрлері ілгекке топтанғанға ұқсайды; дәл осы үшін Mucorды бос тәрізді өңездер деп атайды. Спорангийлер үлкен емес жақындатылған микроскоптан жақсы көрінеді. Мукор тез өседі және шамамен 3 күнде 20 - та барлық Петри тостақаншасында шайылып, өсіп кетеді. Шоғырдың ішкі қуысы әдеттегі эукариот сияқты құрылымды болады, мысалы, Pennicillium (2.25, Г суреті), бір айырмашылығы мукор шоғырларында аралық жоқ. Phizopus Mucor – ға өте ұқсас болып келеді. Столондар деп аталатын кейбір шоғырларда бірнеше қиғашты формалары болады. Столондардың соңында тамырға ұқсас шағын шоғырлы байламдар құрылады, Phizopus – та бір немесе сол жерден екі немесе одан да көп Mucor – дан бөлек спорангиеностар өсіп шығады, яғни спорангиеностар бөлек өсіп шығады.
2.3. Спорангиеностар не үшін керек?
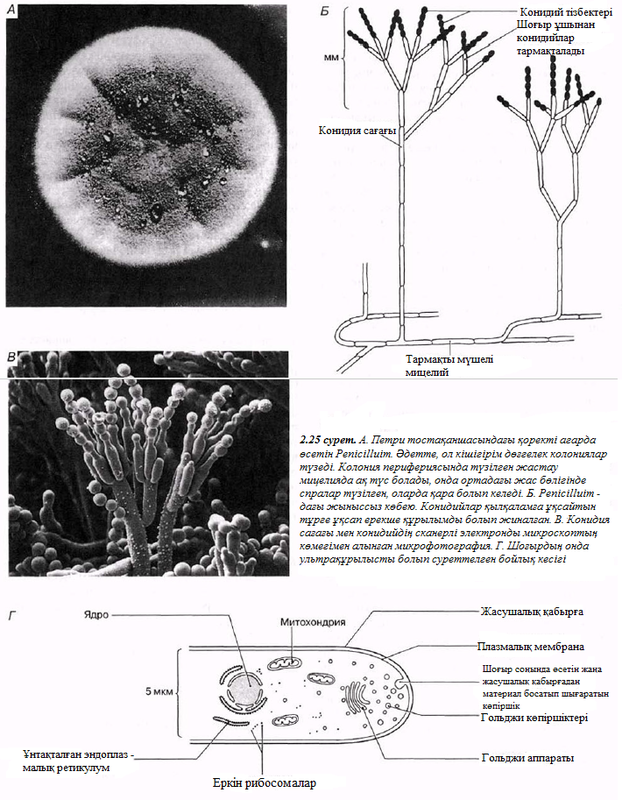
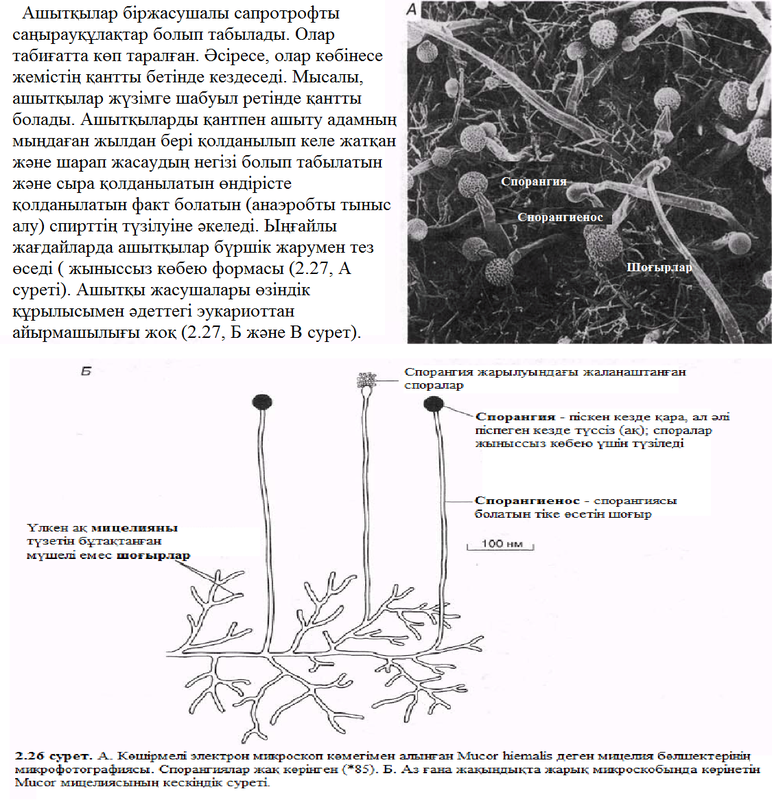
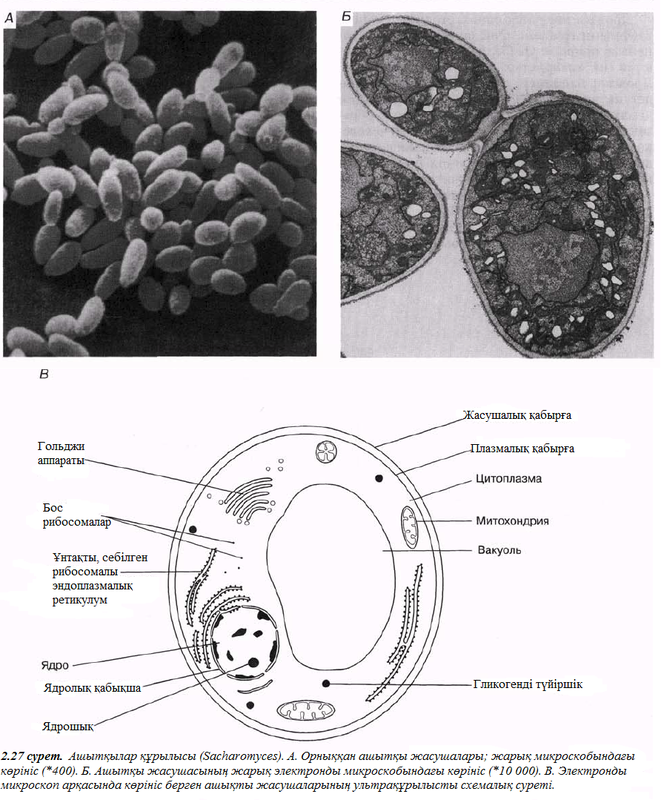
2.5.3. Қоректену
Саңырауқұлақтар – оларға органикалық көміртек көзі керек гетеротрофтар. Одан басқа, оларға, азот көздері (әдетте органикалық, мысалы, аминқышқылдары), неорганикалық иондар (К+ және Mg+ сияқты), микроэлементтер (Fe, Zn және Cu сияқты) және органикалық даму факторлары (витаминдер сияқты). Әртүрлі саңырауқұлақтарға наұты қорек заттары талап етіледі, сондықтан да саңырауқұлақтар таба алатын да субстраттар да әртүрлі. Саңырауқұлақтардың қоректенуі ортаның тура қорек заттарын сіңіру жолымен жүреді, жануарлардан айырмашылығы әдетте бірінші қоректі жұтып алады, сосын қорытады, сөйтіп дененің ішіне кіріп кетеді, тек осыдан кейін ғана қоректік заттарды сіңіру жүреді. Керек кезде саңырауқұлақтар сыртқы қоректі қорытуды жүзеге асыруға қабілетті. Ал бұл жағдайда саңырауқұлақтардың бойынан қорекке ферменттер бөлінеді.
Саңырауқұлақтар қоректену жолы сапротрофтарға, паразиттерге, мутуалисттерге бөлшектенеді. Бұл байланыста олар көпшілігі бактерияға тән болып келеді.
Сапратрофтар
Сапротрофтар деп өлі ағзадан қоректік заттарды шығарушы ағзалар. Сапротрофтарға жатқызылатын саңырауқұлақтар барлық ас қорытатын ферменттерден құралады. Егер сапротроф ас қорытатын ферменттерді 3 негізгі классқа құпиялайтын болса, яғни мына 3 топқа:
Сапротрофты саңырауқұлақтар әдетте оңай тұрақты споралардың көп бөлігін құрады. Бұл басқа да қорек көздеріне оңай таралуға септігін тигізеді. Мұндай саңырауқұлақтардың мысалы мыналар бола алады: Penicillium, Mucor және Phizopus.
Сапротрофты саңырауқұлақтар және бактериялар табиғаттағы биогенді элементтердің айналымдарында маңызды рөл ойнайтын редуценттер тобын құрайды. Әсіресе, маңызды рөлді целлюлозамен лигнинді құпиялайтын және жаңқалаушы аөдаған саңырауқұлақтар алады.
Сондықтан целлюлоза мен лигнин (сүректің негізгі кейпі болып көрінетін күрделі байланыс) өсімдіктердің жасушалық қаңқасында маңызды құрылыды элементтер болып қызмет етеді, целлюлоза мен лигнинді құпиялайтын шіріген сүректер мен басқа да өсетін қалдықтар редуценттер әрекетінің нәтижесінде аздап пайда болады.
Кейбір сапротрофты саңырауқұлақтардың шаруашылықта маңызы зор. Бұл, сонымен қатар, Sacharomyces (ашытқылар) сыра қайнатуда және нан пісіруде Pennicillium – да (12.11.1 бөлім) медицинада қолданылады.
Паразиттер
Паразитті – саңырауқұлақтар факультативті және облигатты болуы мүмкін (2.3.4 бөлім)., әрі олар жануарларға қарағанда өсімдіктерде паразитті тіршілік етеді. Облигатты паразиттер, қағида бойынша, өздерінің иелеріне өлім, бүліншілік шақырмайды, онда факультативті паразиттер бұндайды жиі істейді және сосын сапротрофтар сияқты өлі қалдықтарда тіршілік етеді. Облигатты паразиттерге шынайы ақұнтақтар, жалған ақұнтақтар, жемір саңырауқұлақтар және қаракүйе саңырауқұлақтар жатады. Бұл саңырауқұлақтар астық тұқымдасты дақылдарда негізгі кейіппен паразиттейді, бірақ басқа да ауылшаруашылықтарындағы астықты өсімдіктерді зақымдайды.
Шоғырлар өсімдік ішінде болады, олар кәдімгідей, жасушалар арасында таралып өсе бастайды. Факультативті паразиттер, әдетте, пектиназ сияқты ферменттерді өггндіреді, яғни жасушалар арасындағы орталық пластинканы сіңіріп, тінге жұмсақ өңез шақыырта отырып, оны ботқаға айналдырады. Сон соң жасушалық қабырғаны сіңіретін целлюлозаның көмегімен олар жасушаға басып кіреді жыні оны өлтіреді. Ішіндегі жасушалар, не бірден сіңіріледі, не болмаса басынан саңырауқұлақтың құпиялы ферменттерімен пісіріліп алынады, сосын барып сіңіріледі. Бұндай жолмен, сондай – ақ, Pithium саңырауқұлақтар өсімдіктеріне талқандайды (өскіндерге «суды тұншығушылықты қоздырады) және Phytophtora (картопты шірітуді қоздырғыш), Oomycota бөліміне жататын, жаңа көрсетіліммен сәйкес, саңырауқұлақтардың арғы тегі болатын және Protoctista патшалығына кіретін.
Candida albicans ашытқылары адамға жұқтыратын факультативті паразиттің мысалы бола алады. Бұның жақсысы адамның ішек микрофлорларына зиян келтірмейді, ересектер елінің шамамен 5 %-ын аңғарымды болады. Бірақ, егер біздің денеміздің бетінде немесе ішінде тіршілік ететін микроағзалардың арасында табиғаттық баланс бұзылса, соның ішінде антибиотиктерді қолдану нәидемінде немесе стероидты препараттарды ұзақ қабылдау (иммундық жүйенің басылуына жанама іс – әрекет), ашытқы дамулары бақылаудан алынып тасталып және сонда олар потогенді болып қалып, «Ауызы уылған» (кандидоз) ауруын шақырады. Сондықтан саңырауқұлақ ылғалды жерде көбейеді, ол ауыз қуысын және қынапты зақымдауы мүмкін. Соңғы жағдайларда қынапты іріңдер, сонымен қоса ұышыту немесе несептің шығуы ауруларының саны артуда. Бұл өте көп таралған, сонда да ол саңырауқұлақтарға қарсы ерген әрекет болғандықтан зиянды емес ауру.
Мутуализм (симбиоз)
Саңырауқұлақтар 2 өте маңызды симбиотикалық байланыстың туындысына қатысады, олар қыналар және микориздер. Қыналарды, бұл симбиотикалық саңырауқұлақ пен балдырлардың қауымдастығы – олар жасыл және көк – жасыл (цианобактериялар) болып келеді. Қыналар, әдетте, ашық жартастарда немесе ағаш діңгектерінде, олар тағы да ылғалды орман ағаштарында салбырап тұрады. Балдырлар саңырауқұлақты фотосинтездің органикалық өнімдерімен жетістерсе, ал саңырауқұлақтар қатты күн сәулелерінен қорғанған болса да, суды және минералды тұздарды сіңіріп алуға қабілетті деген болжамдар бар. Саңырауқұлақ, одан басқа, суды жинай алады, қыналарға басқа ешқандай өсімдіктер өспейтін жерде өсуге жағдай жасайды.
Микориза саңырауқұлақпен және өсімдік тамырларымен қауымдастығын құрайды. Саңырауқұлақ олардың ағаштарын жетістіре отырып, минералды тұздармен және суды сіңіреді, ал орнына фотосинтездің органикалық өнімдерін алады. Микориза туралы 7.10.2 бөлімінде қарастырылады.
2.6. Protoctista патшалығы
Гректен аударғанда protos – ең бірінші, ktistos – ашу, құру.
2.6.1. Протоктистердің систематикасы мен құрылысы
2.2. бөлімінде әлдеқашан Маргелиса мен Шварц систематикасының арасындағы патшалыққа (2.4 сурет) Protoctista ең таласты топтар болып табылуы мүмкін, сондықтан кіру мекендерінің арасындағы табиғи байланыстары ағзаларының ішінде өте әлсіз. Шын мәнінде, осы топ эукариот патшалықтарының кез келген біреуі бойынша қатаң өлшемдеріне сәйкес келмейтін барлық эукариот ағзалар бар. Протоктистердің көбісі – біржасушалы ағзалар болып табылады.
Protoctista патшалығына ұқсас немесе қазіргі өсімдіктермен, жануарлармен саңырауқұлақтардың тегіне тән эукариоттар кіреді (2.28 сурет). Бұл жерге тағы ертедегі өсімдіктер (балдырлар), ең алғашқы жануарлар (қарапайымдылар) және ертедегі саңырауқұлақтар (Oomycota) жатады. Одан басқа, бұл патшалық шараналы саңырауқұлақтар секілді, саңырауқұлақтардың спора түзетініне тән топтар құрайды, бірақ бұл жерге бетіндегі жорғалаушылар қабілетті, яғни қозғалатын жануарларға тән. Ең алғашқы эукариоттар жалған аяқтардың көмегімен қозғалатын керемет біржасушалар болған.
Протоктисттер тобы эволюцияға қызығатындар үшін ең тартушы топ болып табылады, сондықтан бұл ағзалар өсімдік пен жануарлар сияқты прокариоттар мен қазіргі эукариоттардың арасындағы түйіні болып қызмет етеді. ХХғ.-дың 60 – шы жылдары, мысалы, митохондриялар – жасуша станциясының күшті аэробты тыныс алу үрдісінен энергияны өндіретіні, кәдімгі өзіндік ДНҚ және рибосомалар және прокариоттаррибосомаларымен ДНҚға ұқсайтыны болған екені ашылды.
Митохондриялы ДНҚ негізгі жүйелілігінің зерттеуі бойынша аса таңғаларлық дәлелдер орын алды, яғни бұрын тектік эукариотттық жасушаларына қоныстанған және оған симбиотикалық түрде тіршілік етуін үйренген митохондриялар ертеде аэробты бактериялар (прокариоттар) болған. Қазір митохондриялар барлық эукариоттың жасушаларында бар және олар тәуелсіз өмірді алып жүруге қабілетсіз.
Саңырауқұлақтар – оларға органикалық көміртек көзі керек гетеротрофтар. Одан басқа, оларға, азот көздері (әдетте органикалық, мысалы, аминқышқылдары), неорганикалық иондар (К+ және Mg+ сияқты), микроэлементтер (Fe, Zn және Cu сияқты) және органикалық даму факторлары (витаминдер сияқты). Әртүрлі саңырауқұлақтарға наұты қорек заттары талап етіледі, сондықтан да саңырауқұлақтар таба алатын да субстраттар да әртүрлі. Саңырауқұлақтардың қоректенуі ортаның тура қорек заттарын сіңіру жолымен жүреді, жануарлардан айырмашылығы әдетте бірінші қоректі жұтып алады, сосын қорытады, сөйтіп дененің ішіне кіріп кетеді, тек осыдан кейін ғана қоректік заттарды сіңіру жүреді. Керек кезде саңырауқұлақтар сыртқы қоректі қорытуды жүзеге асыруға қабілетті. Ал бұл жағдайда саңырауқұлақтардың бойынан қорекке ферменттер бөлінеді.
Саңырауқұлақтар қоректену жолы сапротрофтарға, паразиттерге, мутуалисттерге бөлшектенеді. Бұл байланыста олар көпшілігі бактерияға тән болып келеді.
Сапратрофтар
Сапротрофтар деп өлі ағзадан қоректік заттарды шығарушы ағзалар. Сапротрофтарға жатқызылатын саңырауқұлақтар барлық ас қорытатын ферменттерден құралады. Егер сапротроф ас қорытатын ферменттерді 3 негізгі классқа құпиялайтын болса, яғни мына 3 топқа:
- көміртекті бөлшектегіш ферменттер, мысалы, амилазалар (крахмалды, гликогенді және туыс полисахаридтерді бөлшектейді); 2) липазалар (липидтерді бөлшектейді) және 3) протеиназалар (нәруыздарды бөлшектейді); сонда ғана ол әртүрлі субстраттарды қолдана алады. Pennicillium түрлері субстраттарға, жерде, қайналмаған тері, нанда және бұзылған жемістерде жасыл және көк өңездер құрады.
Сапротрофты саңырауқұлақтар әдетте оңай тұрақты споралардың көп бөлігін құрады. Бұл басқа да қорек көздеріне оңай таралуға септігін тигізеді. Мұндай саңырауқұлақтардың мысалы мыналар бола алады: Penicillium, Mucor және Phizopus.
Сапротрофты саңырауқұлақтар және бактериялар табиғаттағы биогенді элементтердің айналымдарында маңызды рөл ойнайтын редуценттер тобын құрайды. Әсіресе, маңызды рөлді целлюлозамен лигнинді құпиялайтын және жаңқалаушы аөдаған саңырауқұлақтар алады.
Сондықтан целлюлоза мен лигнин (сүректің негізгі кейпі болып көрінетін күрделі байланыс) өсімдіктердің жасушалық қаңқасында маңызды құрылыды элементтер болып қызмет етеді, целлюлоза мен лигнинді құпиялайтын шіріген сүректер мен басқа да өсетін қалдықтар редуценттер әрекетінің нәтижесінде аздап пайда болады.
Кейбір сапротрофты саңырауқұлақтардың шаруашылықта маңызы зор. Бұл, сонымен қатар, Sacharomyces (ашытқылар) сыра қайнатуда және нан пісіруде Pennicillium – да (12.11.1 бөлім) медицинада қолданылады.
Паразиттер
Паразитті – саңырауқұлақтар факультативті және облигатты болуы мүмкін (2.3.4 бөлім)., әрі олар жануарларға қарағанда өсімдіктерде паразитті тіршілік етеді. Облигатты паразиттер, қағида бойынша, өздерінің иелеріне өлім, бүліншілік шақырмайды, онда факультативті паразиттер бұндайды жиі істейді және сосын сапротрофтар сияқты өлі қалдықтарда тіршілік етеді. Облигатты паразиттерге шынайы ақұнтақтар, жалған ақұнтақтар, жемір саңырауқұлақтар және қаракүйе саңырауқұлақтар жатады. Бұл саңырауқұлақтар астық тұқымдасты дақылдарда негізгі кейіппен паразиттейді, бірақ басқа да ауылшаруашылықтарындағы астықты өсімдіктерді зақымдайды.
Шоғырлар өсімдік ішінде болады, олар кәдімгідей, жасушалар арасында таралып өсе бастайды. Факультативті паразиттер, әдетте, пектиназ сияқты ферменттерді өггндіреді, яғни жасушалар арасындағы орталық пластинканы сіңіріп, тінге жұмсақ өңез шақыырта отырып, оны ботқаға айналдырады. Сон соң жасушалық қабырғаны сіңіретін целлюлозаның көмегімен олар жасушаға басып кіреді жыні оны өлтіреді. Ішіндегі жасушалар, не бірден сіңіріледі, не болмаса басынан саңырауқұлақтың құпиялы ферменттерімен пісіріліп алынады, сосын барып сіңіріледі. Бұндай жолмен, сондай – ақ, Pithium саңырауқұлақтар өсімдіктеріне талқандайды (өскіндерге «суды тұншығушылықты қоздырады) және Phytophtora (картопты шірітуді қоздырғыш), Oomycota бөліміне жататын, жаңа көрсетіліммен сәйкес, саңырауқұлақтардың арғы тегі болатын және Protoctista патшалығына кіретін.
Candida albicans ашытқылары адамға жұқтыратын факультативті паразиттің мысалы бола алады. Бұның жақсысы адамның ішек микрофлорларына зиян келтірмейді, ересектер елінің шамамен 5 %-ын аңғарымды болады. Бірақ, егер біздің денеміздің бетінде немесе ішінде тіршілік ететін микроағзалардың арасында табиғаттық баланс бұзылса, соның ішінде антибиотиктерді қолдану нәидемінде немесе стероидты препараттарды ұзақ қабылдау (иммундық жүйенің басылуына жанама іс – әрекет), ашытқы дамулары бақылаудан алынып тасталып және сонда олар потогенді болып қалып, «Ауызы уылған» (кандидоз) ауруын шақырады. Сондықтан саңырауқұлақ ылғалды жерде көбейеді, ол ауыз қуысын және қынапты зақымдауы мүмкін. Соңғы жағдайларда қынапты іріңдер, сонымен қоса ұышыту немесе несептің шығуы ауруларының саны артуда. Бұл өте көп таралған, сонда да ол саңырауқұлақтарға қарсы ерген әрекет болғандықтан зиянды емес ауру.
Мутуализм (симбиоз)
Саңырауқұлақтар 2 өте маңызды симбиотикалық байланыстың туындысына қатысады, олар қыналар және микориздер. Қыналарды, бұл симбиотикалық саңырауқұлақ пен балдырлардың қауымдастығы – олар жасыл және көк – жасыл (цианобактериялар) болып келеді. Қыналар, әдетте, ашық жартастарда немесе ағаш діңгектерінде, олар тағы да ылғалды орман ағаштарында салбырап тұрады. Балдырлар саңырауқұлақты фотосинтездің органикалық өнімдерімен жетістерсе, ал саңырауқұлақтар қатты күн сәулелерінен қорғанған болса да, суды және минералды тұздарды сіңіріп алуға қабілетті деген болжамдар бар. Саңырауқұлақ, одан басқа, суды жинай алады, қыналарға басқа ешқандай өсімдіктер өспейтін жерде өсуге жағдай жасайды.
Микориза саңырауқұлақпен және өсімдік тамырларымен қауымдастығын құрайды. Саңырауқұлақ олардың ағаштарын жетістіре отырып, минералды тұздармен және суды сіңіреді, ал орнына фотосинтездің органикалық өнімдерін алады. Микориза туралы 7.10.2 бөлімінде қарастырылады.
2.6. Protoctista патшалығы
Гректен аударғанда protos – ең бірінші, ktistos – ашу, құру.
2.6.1. Протоктистердің систематикасы мен құрылысы
2.2. бөлімінде әлдеқашан Маргелиса мен Шварц систематикасының арасындағы патшалыққа (2.4 сурет) Protoctista ең таласты топтар болып табылуы мүмкін, сондықтан кіру мекендерінің арасындағы табиғи байланыстары ағзаларының ішінде өте әлсіз. Шын мәнінде, осы топ эукариот патшалықтарының кез келген біреуі бойынша қатаң өлшемдеріне сәйкес келмейтін барлық эукариот ағзалар бар. Протоктистердің көбісі – біржасушалы ағзалар болып табылады.
Protoctista патшалығына ұқсас немесе қазіргі өсімдіктермен, жануарлармен саңырауқұлақтардың тегіне тән эукариоттар кіреді (2.28 сурет). Бұл жерге тағы ертедегі өсімдіктер (балдырлар), ең алғашқы жануарлар (қарапайымдылар) және ертедегі саңырауқұлақтар (Oomycota) жатады. Одан басқа, бұл патшалық шараналы саңырауқұлақтар секілді, саңырауқұлақтардың спора түзетініне тән топтар құрайды, бірақ бұл жерге бетіндегі жорғалаушылар қабілетті, яғни қозғалатын жануарларға тән. Ең алғашқы эукариоттар жалған аяқтардың көмегімен қозғалатын керемет біржасушалар болған.
Протоктисттер тобы эволюцияға қызығатындар үшін ең тартушы топ болып табылады, сондықтан бұл ағзалар өсімдік пен жануарлар сияқты прокариоттар мен қазіргі эукариоттардың арасындағы түйіні болып қызмет етеді. ХХғ.-дың 60 – шы жылдары, мысалы, митохондриялар – жасуша станциясының күшті аэробты тыныс алу үрдісінен энергияны өндіретіні, кәдімгі өзіндік ДНҚ және рибосомалар және прокариоттаррибосомаларымен ДНҚға ұқсайтыны болған екені ашылды.
Митохондриялы ДНҚ негізгі жүйелілігінің зерттеуі бойынша аса таңғаларлық дәлелдер орын алды, яғни бұрын тектік эукариотттық жасушаларына қоныстанған және оған симбиотикалық түрде тіршілік етуін үйренген митохондриялар ертеде аэробты бактериялар (прокариоттар) болған. Қазір митохондриялар барлық эукариоттың жасушаларында бар және олар тәуелсіз өмірді алып жүруге қабілетсіз.
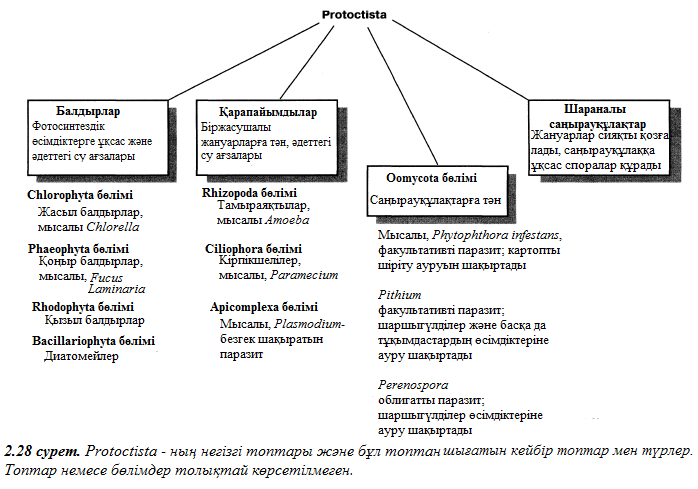
Митохондрияға ұқсас хлоропласттарда фотосинтез жүретін, сонымен қатар, өздерінің прокариоттық ДНҚ мен рибосомалары бар хлорофиллді органеллалары бар. Мүмкін, хлоропласттар – өзінің уақытында жануарларға тән гетеророфты жасушаларда автотрофты балдырларға айналдыра отырып, орналасқан фотосинтезді бактериялардан пайда болған болар. Дәл осындай, қызыл балдырларда да болуы мүмкін, қызыл балдырлар көк – жасыл бактериялардан (цианобактериялардан) пайда болуы мүмкін, ал жасыл балдырлар – прохлофиттер сияқты жасыл бактериялардан болса керек.
Митохондрия мен хлоропласттар симбиотикалық бактериялардың ұрпақтары болып табылады жеген теория эндосимбиотикалық теориялар деген атау алды. Эндосимбионт – өзге ағзаның ішінде (endo – ) симбиотикалық түрде тіршілк ететін 2.6.2. Oomycota бөлімі
Оомицеттер (Oomycota) – өздерінің құрылысы бойынша саңырауқұлақтардың жақын туыстары болып саналады, бірақ қазіргі херттеулермен келісуге де болады, себебі бұл кішкене болса да ескі топ. Оомицеттерде жасушалық қабырға ретінде тірек болатын саңырауқұлақ сияқты, хитин мен целлюлоза болып табылады. Олардың шоғырлары аралық болып бөлінбеген (беймүшелілер). Оомицеттерге жалған ақұнтақты – мильдяны қосқанда патогенді саңырауқұлақтардың барлық қатары жатады. Мұндайға мысал үшін паразиттік саңырауқұлақтар облигатты паразит болып саналатын Phitophthora infestans – ті алуға болады. Салыстыру үшін басқа облигатты паращитті қарастырамыз және ақыры соңында әдеттегі факультативті паразит Pithium – ді қарастырамыз. Облигатты паразиттерге өмір сүруге бейім және тірі жасушаларда ерекше дами алатын, ал факультативті паразиттерден айырмашылығы : өзінің иелеріне зақым әкеледі де, содан соң барып өлі қалдықтарда сапрофотты тіршілік ететін ағзалар жатады.
Phitophthora infestans
Phitophthora infestans – аса маңызды шаруашылық мағынасы бар патогенді саңырауқұлақ, ол картопта паразиттейді және өрісті бүлдіреді және ол «картоп бүліну» атты маңызды ауру шақырады. P. Infestans облигатты паразиттерге ұқсайтын жері иесінен тәуелсіз дамуға қабілетсіз. Фитотрофты жұқтыраудағы
Митохондрия мен хлоропласттар симбиотикалық бактериялардың ұрпақтары болып табылады жеген теория эндосимбиотикалық теориялар деген атау алды. Эндосимбионт – өзге ағзаның ішінде (endo – ) симбиотикалық түрде тіршілк ететін 2.6.2. Oomycota бөлімі
Оомицеттер (Oomycota) – өздерінің құрылысы бойынша саңырауқұлақтардың жақын туыстары болып саналады, бірақ қазіргі херттеулермен келісуге де болады, себебі бұл кішкене болса да ескі топ. Оомицеттерде жасушалық қабырға ретінде тірек болатын саңырауқұлақ сияқты, хитин мен целлюлоза болып табылады. Олардың шоғырлары аралық болып бөлінбеген (беймүшелілер). Оомицеттерге жалған ақұнтақты – мильдяны қосқанда патогенді саңырауқұлақтардың барлық қатары жатады. Мұндайға мысал үшін паразиттік саңырауқұлақтар облигатты паразит болып саналатын Phitophthora infestans – ті алуға болады. Салыстыру үшін басқа облигатты паращитті қарастырамыз және ақыры соңында әдеттегі факультативті паразит Pithium – ді қарастырамыз. Облигатты паразиттерге өмір сүруге бейім және тірі жасушаларда ерекше дами алатын, ал факультативті паразиттерден айырмашылығы : өзінің иелеріне зақым әкеледі де, содан соң барып өлі қалдықтарда сапрофотты тіршілік ететін ағзалар жатады.
Phitophthora infestans
Phitophthora infestans – аса маңызды шаруашылық мағынасы бар патогенді саңырауқұлақ, ол картопта паразиттейді және өрісті бүлдіреді және ол «картоп бүліну» атты маңызды ауру шақырады. P. Infestans облигатты паразиттерге ұқсайтын жері иесінен тәуелсіз дамуға қабілетсіз. Фитотрофты жұқтыраудағы
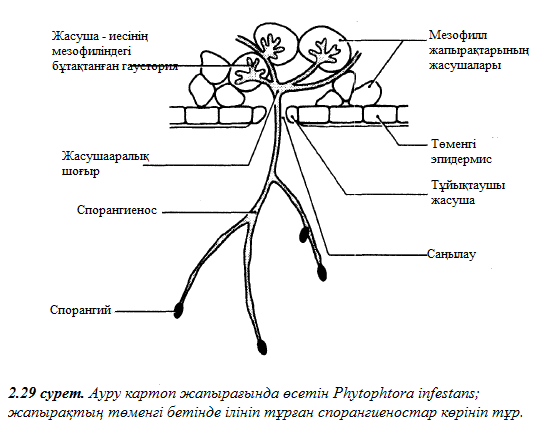
өзінің құрылысы мен қабілеті сары фиоль ауруы мен капуста және басқа да шаршыгүлді мен тұқымдастарына айтарлықтай таралған қоздырғыш болып табылатын (Crucifera) Oomycota – ның тағы бір өнімі Perenospora – ға ұқсас.
Фитофтор мицелийлері картоп түйнектерінде қыстап, өсімдіктің жапырақтарында көктемнен бастап өседі, тамызда шірудің алғашқы белгілері байқалады.
Тармақталған беймүшелі шоғырдан құратын мицелий гасторийді құрай отырып, жапырақтардың ішінде жасушааралық кеңістікте таралады, бұл гасторийлар мезофил жасушасына өтіп кетеді де, олардан қоректік заттарды сорып алады (2.29 сурет). Гаустоийлар – әдеттегі облигатты паразиттер үшін мамандандырушылар. Мұндай өсінділер (гаусторийлар) плазматикалық мембраналы бұзбай және жасушаны өлтірмей жасушаға еніп кетеді. Жапырақтың астыңғы жағынан жапырақ саңылауы және жара арқылы түсетін мицелийлер жылы және ылғал ортада спорангиеностардды құрайды. Спорангийлер болып басталып, спорангиеностар тармақталады (2.29 сурет). Жылы ортада спорангийлер споралар болып саналады, яғни бұтақ арқылы ауысады немесе өзге өсімдіктерге жаңбыр тамшысы арқылы шашыранды болып келеді, осындай жолмен инфекция таралады. Сосын жапырақ саңылауынан жасымықшадан немесе өсімдік тінінің зақымдануынан пайда болатын спорангийден шоғыр құралады. Салқын жағдайларда спорангий ішіндегілері спорангийден босатылып шығып жапырақтың үстіңгі сіңген кішігірім сұйық қабатында жүзіп жүретін жылжымалы зооспораның (бұл жабайы ағзаларға тән белгі) түзілуімен бөлінеді. Зооспоралар инцистирлейді және тіпті шоғырдың дамуына лайық жағдайды күтуге де дайын; онда жаңа өсімдіктерге зақым келе бастайды.
Бөлек жапырақтағы ауру өсімдіктерде кіші көлемді өлі, зақымдалған қоңыр түсті зоналар байқалады. Егер мұқият қарайтын болсақ, зақымдалған жапырақтардың астыңғы беткейінде өлі зонаның маңында ақ спорангийлерден шашақтарды байқауға болады. Жылы дымқыл ауа – райында өлі – еттену зоналары жапырақ беткейіне тез таралып сабаққа көшеді. Кейбір спорангийлер жерге түседі және картоп түйнектеріне зиян келтіреді; ал бұдан нфекция өте тез таралалады және түйнектіні әрқалай перифериядан центрге қарай тарап қоңырлаутүске өзгеретін өзінің тұқымынан құрғақ шірікті шақыртады.
Ең бірінші түбірлі мойыннан бастап, сосын басқа да барлық өсімдіктің мүшелері некроза зонасы сияқты 2 – ші рет сапротрофты редуцент – бактерияларына індет жұқтыратыны сияқты қою шірікке айналады. Осылайша, фитофтора жақын туысынан ажыратып тұратын өсімдік туысынан ажыратып тұратын өсімдікті өлтіреді - Perenospora, бұл облигатты паразит. Бұл жерде Phitophthora, әдеттегі, облигатты паразитке ұқсай бермейді, кейде оны факультативті паразитке де жатқызып жатады, тіпті мұндай аз ғана өзгешілікке аса мән беріп тоқтаудың қажеті жоқ.
Фитофтора, қағида бойынша, ұйқыдағы мицелия ретінде індет жұққыш картоп түйнегінің ішінде ақырын ғана қыстап шығады. Бұл саңырауқұлақтың Perenospora – дан айырмашылығы, ол жыныстық жолмен сирек қана көбейеді, егер, әрине картоп өндірген жерлерді айтпасақ қана (Мексика, Оңтүстік және Орталық Америка). Саңырауқұлақтың жыныстық көбеюі лабораториялық жағдайларда ғана индукцияланады.
Perenospora сияқты фитофторалық әрекетсіз спораларды түзеді. Қабырғасы қалың споралар антеридия аталық репродуктивті ағза мен ооганияның аналық репродуктивті ағзасының бірігуі мен пайда болады. Мұндай спора қыртыста қыстайды және келесі жылы жаңа инфекция шақырады.
Фитофтора шақыратын өткен эпидемияде үлкен зардапқа әкеп соқтырады. ХХғ. – дың соңғы 30 жылдарында Америкадан Еуропаға ауру кенеттен келді деген болжам бар. Нәтижесінде, Еуропада эпидемия толқыны түгелдей барлық жерді сырғып кетті, 1845 жылдың соңғы кездері Ирландияда картоп егні түгелімен жойылып кетті. Көп адамдардың басына аштық түсіп, қырылу болды, бұл құрбандық тек картоп ауруы болған жоқ, ол тіпті ауыр политикалық және экономикалық жүйелікке әкеліп соқтырды. Нәтижесінде, Ирландықтар Солтүстік Америкаға көшуге мәжбүр болды.
Бұл ауру тағы 1845 жылы Беркли алғаш рет оған микробты табиғат көрсеткенімен қызығушылық танытып отыр. Беркли саңырауқұлақ шірудің жанама өнімі емес, картоппен байланысы бар және өзі ауру шақыртатынын көрсетіп берді.
Картопты шірудің тіршілік кезеңінің түсіндірілуі бұл аурудың алдын алу әдістерін шығаруға әкелді. Төменде бұл әдітер көрсетілген:
Pithium – ның фитофторалардан айырмашылығы олар әртүрлі көптеген өсімдіктерді зақымдаушы және жұмсақ шіруді шақыртатын мамандандырылмаған паразит. Pithium өскіндерге «суда тұншығуды» шақыртады. Оған ылғал орта қажет, сондықтан олар жыныссыз көбеюде жүзгіш спораларды түзеді. Бұның саңырауқұлағы тірі өсімдік ретінде жіне өлі қалдықтартарда да өсе алады, демек, бұл факультативті паразит. Ол және сапротрофты болып дымқыл жерде тіршілік ете алады. Pithium ферментті сыртқы ортасына қарап бөледі, швбуыл жасуға және тез иесін өлтіруге мүмкіндігі бар. Ферменттің алғашқылары алдында өсіп тұрған саңырауқұлақты диффуздайтын пектиназдарды құрайды, ол жасушалары біріккен орташа пластинкадағы пектиндерді ерітеді. Өсімдік тіні нәтижесінде былыққа (жұмсақ өңез) айналады және өсімдік өледі. Артынан құрамында өсімдіктің жасушаларды сіңіртетін басқа ферменттер пайда болады. Бірақ Pithium – дағы гаусторийлар фитофторалардан айырмашлығы : олар түзілмейді. Қорыту өнімдірері жасуша арасында өсетін шоғырларды сорып алады.
Жер бетіндегі өсінділердің пайда болуының бүліну су басу салдары себепші болады. Сабақта алғашқы сулы нүктелер жер дәрежесінде пайда болады. Олар қарайған кезде сабақтар өледі. Өсінділердің суда тұншығуы бау – бақша шаруашылығында, орман шаруашылығында және ауыл шаруашылығында ауыр мәселелер шығаруы мүмкін. Шашақгүлділер тұқымдасы өкілдері бұл ауруға сезімтал болып келеді, әсіресе, өсінділер өте тығыз болған кезде.
2.6.3. Балдырлар
Биологиялық мәні бар және адам өмірінде өте маңызды рөл ойнайтын протоктистер тобын балдырлар тұзеді. Оларда бірде бір диагностикалық белгілер жоқ. Бәрінен бұрын таңғалатыны бұлар сулы ортада эволюцияланған және ол жерде қалмаған фотосинтездеуші эукариоттар болып шықты. Кейбір судан шыққан балдырлар құрғақ өмірге сәтті түрде бейімделіп кетеді,бірақ өсімдіктерден айырмашылығы жердегі балдырлар мұхиттың және тұщы судағы формалармен салыстырып қарағанда аз ғана айырмашылық бар. Балдырлар денесі сабаққа, тамырға және жапыраққа бөлінбейді. Мұндайға қатысты сараланбаған денені талломдар деп атайды.
Балдырлар өздерінің негізгі фотосинтездік пигменттерімен ажыратылатын анық көрсетілген табиғи топтардың қатарын құрайды. Заманауи классификацияда бұл топтар, бөлімдер мәртебесін алды. 2.28 суретінде тек қана балдырлардың 4 бөлімін келтіреді. Балдырлар сипаттамасы жалпылама негізгі 2 бөлімі 2.7 кестесінде келтірілген. Chlorella (Chlorophyta бөлімі) және Fucus (Phaeophyta бөлімі) сияқты балдырлардың 2 өкілі төменде толығырақ сипатталады.
Фитофтор мицелийлері картоп түйнектерінде қыстап, өсімдіктің жапырақтарында көктемнен бастап өседі, тамызда шірудің алғашқы белгілері байқалады.
Тармақталған беймүшелі шоғырдан құратын мицелий гасторийді құрай отырып, жапырақтардың ішінде жасушааралық кеңістікте таралады, бұл гасторийлар мезофил жасушасына өтіп кетеді де, олардан қоректік заттарды сорып алады (2.29 сурет). Гаустоийлар – әдеттегі облигатты паразиттер үшін мамандандырушылар. Мұндай өсінділер (гаусторийлар) плазматикалық мембраналы бұзбай және жасушаны өлтірмей жасушаға еніп кетеді. Жапырақтың астыңғы жағынан жапырақ саңылауы және жара арқылы түсетін мицелийлер жылы және ылғал ортада спорангиеностардды құрайды. Спорангийлер болып басталып, спорангиеностар тармақталады (2.29 сурет). Жылы ортада спорангийлер споралар болып саналады, яғни бұтақ арқылы ауысады немесе өзге өсімдіктерге жаңбыр тамшысы арқылы шашыранды болып келеді, осындай жолмен инфекция таралады. Сосын жапырақ саңылауынан жасымықшадан немесе өсімдік тінінің зақымдануынан пайда болатын спорангийден шоғыр құралады. Салқын жағдайларда спорангий ішіндегілері спорангийден босатылып шығып жапырақтың үстіңгі сіңген кішігірім сұйық қабатында жүзіп жүретін жылжымалы зооспораның (бұл жабайы ағзаларға тән белгі) түзілуімен бөлінеді. Зооспоралар инцистирлейді және тіпті шоғырдың дамуына лайық жағдайды күтуге де дайын; онда жаңа өсімдіктерге зақым келе бастайды.
Бөлек жапырақтағы ауру өсімдіктерде кіші көлемді өлі, зақымдалған қоңыр түсті зоналар байқалады. Егер мұқият қарайтын болсақ, зақымдалған жапырақтардың астыңғы беткейінде өлі зонаның маңында ақ спорангийлерден шашақтарды байқауға болады. Жылы дымқыл ауа – райында өлі – еттену зоналары жапырақ беткейіне тез таралып сабаққа көшеді. Кейбір спорангийлер жерге түседі және картоп түйнектеріне зиян келтіреді; ал бұдан нфекция өте тез таралалады және түйнектіні әрқалай перифериядан центрге қарай тарап қоңырлаутүске өзгеретін өзінің тұқымынан құрғақ шірікті шақыртады.
Ең бірінші түбірлі мойыннан бастап, сосын басқа да барлық өсімдіктің мүшелері некроза зонасы сияқты 2 – ші рет сапротрофты редуцент – бактерияларына індет жұқтыратыны сияқты қою шірікке айналады. Осылайша, фитофтора жақын туысынан ажыратып тұратын өсімдік туысынан ажыратып тұратын өсімдікті өлтіреді - Perenospora, бұл облигатты паразит. Бұл жерде Phitophthora, әдеттегі, облигатты паразитке ұқсай бермейді, кейде оны факультативті паразитке де жатқызып жатады, тіпті мұндай аз ғана өзгешілікке аса мән беріп тоқтаудың қажеті жоқ.
Фитофтора, қағида бойынша, ұйқыдағы мицелия ретінде індет жұққыш картоп түйнегінің ішінде ақырын ғана қыстап шығады. Бұл саңырауқұлақтың Perenospora – дан айырмашылығы, ол жыныстық жолмен сирек қана көбейеді, егер, әрине картоп өндірген жерлерді айтпасақ қана (Мексика, Оңтүстік және Орталық Америка). Саңырауқұлақтың жыныстық көбеюі лабораториялық жағдайларда ғана индукцияланады.
Perenospora сияқты фитофторалық әрекетсіз спораларды түзеді. Қабырғасы қалың споралар антеридия аталық репродуктивті ағза мен ооганияның аналық репродуктивті ағзасының бірігуі мен пайда болады. Мұндай спора қыртыста қыстайды және келесі жылы жаңа инфекция шақырады.
Фитофтора шақыратын өткен эпидемияде үлкен зардапқа әкеп соқтырады. ХХғ. – дың соңғы 30 жылдарында Америкадан Еуропаға ауру кенеттен келді деген болжам бар. Нәтижесінде, Еуропада эпидемия толқыны түгелдей барлық жерді сырғып кетті, 1845 жылдың соңғы кездері Ирландияда картоп егні түгелімен жойылып кетті. Көп адамдардың басына аштық түсіп, қырылу болды, бұл құрбандық тек картоп ауруы болған жоқ, ол тіпті ауыр политикалық және экономикалық жүйелікке әкеліп соқтырды. Нәтижесінде, Ирландықтар Солтүстік Америкаға көшуге мәжбүр болды.
Бұл ауру тағы 1845 жылы Беркли алғаш рет оған микробты табиғат көрсеткенімен қызығушылық танытып отыр. Беркли саңырауқұлақ шірудің жанама өнімі емес, картоппен байланысы бар және өзі ауру шақыртатынын көрсетіп берді.
Картопты шірудің тіршілік кезеңінің түсіндірілуі бұл аурудың алдын алу әдістерін шығаруға әкелді. Төменде бұл әдітер көрсетілген:
- Бірде - бір шірік түйнек отығызылмау керек екені мұқият қаралуы тиіс;
- Сондықтан саңырауқұлақ жер қыртысында 1 жыл сақталады, бұл ауру болып кеткен жерге картоп егуге болмайды. Бұл жағдайларда дұрыс егіс айналымдар болған жөн.
- Түйнекті тартып шығаруға дейін барлық ауру өсімдіктер бөліктерін құртып тастап отыру керек, мысалы, оларды өртеп немесе улы ерітінділермен сеуіп тастап, мысалы күкірт қышқылы. Бұл сасық сабақтар (яғни, сабақтар) және басқа да жер үсті бөліктері түйнекті де бүлдіруі мүмкін.
- Сондықтан да бұл саңырауқұлақ қозылмаған түйнектерде қыстап шығуына болады және бүлінген аулада бірде бір түйнек қалмауын мұқият қадағалау керек.
- Саңырауқұлақты мысты функциялармен, мысалы, бордосстық сұйықтыларымен жоюға болады. Бүркітуді ауыруды ескертуге үлгеру үшін, нақты белгілі бір уақытта өткізу керек, олай болмаса зардап шеккен өсімдіктерге ештеңе де істеу алмаймыз: өсімдіктерге әдетте бүркуді әрбір 2 апта сайын істейді, яғни бірнеше сантиметрге дейін өскенннен және толықтай өскенге дейін соңғы егуге жиналып қойған түйнектер сырттан сұйық хлорлы сынап ерітінділеріне батырып алып стирильдеп алуға болады.
- Метеорологиялық жағдайларға үзбестен көз салып отыру және фермерлерге ерте хабарлау егуді қашан бүрку керек екенін анықтауға болады.
- Шірудің тұрақтылығына байланысты картоптарға селекция бір уақыт өтіп шықты. Белгілі болғандай, Solanum demissum атты жабайы картоп фитофторға деген жоғары деңгейін тұрақтылығы бар, сондықтан оны селекцияға қолданды. Керек иммунитеттің алынуына ең үлкен тосқауыл бұл саңырауқұлақтың көптеген штаммдары болуында және әлі де болсын барлық штаммға тұрақты бола білетін картоптың бір сорты да табылған жоқ. Картоптың жаңа сорттарын дақылға келтіру шаралары тағы бұл саңырауқұлақтың жаңа штаммдары келуіне әкелуі мүмкін. Көрсетілген мәселе фитопатологтарға бұрыннан мәлім, ол бізге артық рет біздің заманауи ауылшаруашылық дақылдарының жабайы тектіктің тектік қорын әртүрлі аурудың түрлерін гендердің тұрақтылық көзі сияқты сақтау керек екенін еске түсіріп қояды.
Pithium – ның фитофторалардан айырмашылығы олар әртүрлі көптеген өсімдіктерді зақымдаушы және жұмсақ шіруді шақыртатын мамандандырылмаған паразит. Pithium өскіндерге «суда тұншығуды» шақыртады. Оған ылғал орта қажет, сондықтан олар жыныссыз көбеюде жүзгіш спораларды түзеді. Бұның саңырауқұлағы тірі өсімдік ретінде жіне өлі қалдықтартарда да өсе алады, демек, бұл факультативті паразит. Ол және сапротрофты болып дымқыл жерде тіршілік ете алады. Pithium ферментті сыртқы ортасына қарап бөледі, швбуыл жасуға және тез иесін өлтіруге мүмкіндігі бар. Ферменттің алғашқылары алдында өсіп тұрған саңырауқұлақты диффуздайтын пектиназдарды құрайды, ол жасушалары біріккен орташа пластинкадағы пектиндерді ерітеді. Өсімдік тіні нәтижесінде былыққа (жұмсақ өңез) айналады және өсімдік өледі. Артынан құрамында өсімдіктің жасушаларды сіңіртетін басқа ферменттер пайда болады. Бірақ Pithium – дағы гаусторийлар фитофторалардан айырмашлығы : олар түзілмейді. Қорыту өнімдірері жасуша арасында өсетін шоғырларды сорып алады.
Жер бетіндегі өсінділердің пайда болуының бүліну су басу салдары себепші болады. Сабақта алғашқы сулы нүктелер жер дәрежесінде пайда болады. Олар қарайған кезде сабақтар өледі. Өсінділердің суда тұншығуы бау – бақша шаруашылығында, орман шаруашылығында және ауыл шаруашылығында ауыр мәселелер шығаруы мүмкін. Шашақгүлділер тұқымдасы өкілдері бұл ауруға сезімтал болып келеді, әсіресе, өсінділер өте тығыз болған кезде.
2.6.3. Балдырлар
Биологиялық мәні бар және адам өмірінде өте маңызды рөл ойнайтын протоктистер тобын балдырлар тұзеді. Оларда бірде бір диагностикалық белгілер жоқ. Бәрінен бұрын таңғалатыны бұлар сулы ортада эволюцияланған және ол жерде қалмаған фотосинтездеуші эукариоттар болып шықты. Кейбір судан шыққан балдырлар құрғақ өмірге сәтті түрде бейімделіп кетеді,бірақ өсімдіктерден айырмашылығы жердегі балдырлар мұхиттың және тұщы судағы формалармен салыстырып қарағанда аз ғана айырмашылық бар. Балдырлар денесі сабаққа, тамырға және жапыраққа бөлінбейді. Мұндайға қатысты сараланбаған денені талломдар деп атайды.
Балдырлар өздерінің негізгі фотосинтездік пигменттерімен ажыратылатын анық көрсетілген табиғи топтардың қатарын құрайды. Заманауи классификацияда бұл топтар, бөлімдер мәртебесін алды. 2.28 суретінде тек қана балдырлардың 4 бөлімін келтіреді. Балдырлар сипаттамасы жалпылама негізгі 2 бөлімі 2.7 кестесінде келтірілген. Chlorella (Chlorophyta бөлімі) және Fucus (Phaeophyta бөлімі) сияқты балдырлардың 2 өкілі төменде толығырақ сипатталады.

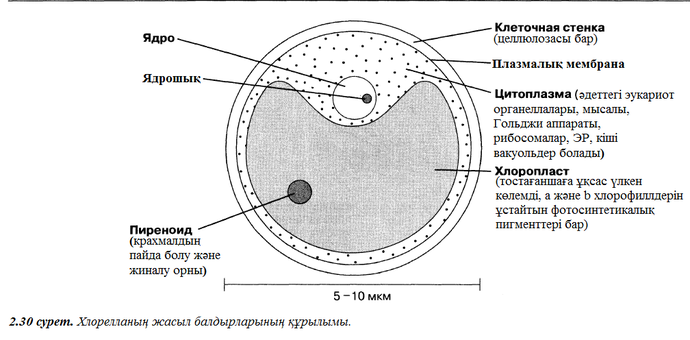
2.6.4. Chlorophyta бөлімі (жасыл балдырлар)
Хлорелла
Хлорелла – біржасушалы қозғалмайтын жасыл балдыр. Оның 2.30 суретінде көрсетілген. Оны тұщы сулы тағамдарда және арықтарда кездестіруге болады. Хлорелланы оңай дақылдауға болады және ол фотосинтезді зерттеу эксперименттерінде кеңінен қолданылады (7.6 бөлім), сонымен қатар қорекктің баламалы көзі ретінде (12.12.3 бөлімі; біржасуашалы ақуыздар).
2.6.5. Phaeophyta бөлімі (қоғыр балдырлар)
Фукус
Фукус – күрделі құрылымды ірі қоңыр балдыр болып келеді. Оның денесі қысқа шыбыққа, негізгі дөңгелекке және мүкке бөлінген (бұл жалған сабақ, тамыр және жапырақтар екенін айта кету керек) өзін талломмен көрсетіледі. Бұл теңіз балдыры жартасты Британ жағалауында жиі кездеседі. ол жағалаудың кезектесіп құрғақ жерінің артуына және кемуіне, бірде жалаңаштануына, бірде сумен жабылуына бейімделген. Фукустың 3 түрлі зоналарда немесе тереңдіктерде кең таралған 3 түрі белгілі, жағалаудағы аймақтық бөліну деп аталады (10.6.4 бөлімі). Балдырларды аймаққа байланысты бөлу олардың ауада қаншалықты бола алатынына байланысты болып келеді. Төменде оларды қалай тану керектігі туралы негізгі белгілер көрсетілген, сонымен қатар, олардың таралу орындары көрсетілген.
F. spiralis (тегіс балдыр) – оларды жағалаудың ең биік жоғарылаудың нүктесіне лақтырады. Таллом батқан жағдайда шиыршыққа оңайлықпен бұрала алады.
F. serratus (қарапайым тісті немесе ара тісі тәрізді балдыр) – жартылай жоғарылау аймағында. Таллом шеті кетілдірілген.
F. vesicullosus (көпіршікті балдыр) – төмендеудің ең төмен белгісі. Ауа көпіршіктері бар жүзгіш балдырлармен қамтамасыз етілген 2.31 суретінде F. vesicullosus – тың сыртқы белгілері көрсетілген.
ҚОРШАҒАН ОРТАҒА БЕЙІМДЕЛГІШТІК. Фукустың қорек ортасына бейімделгіштігін айтпастан бұрын, оның айтарлықтай қонақжай емес туралы табиғаты туралы айта кету керек. жоғары – төмен аймағының өсімдіктері бола тұра, әр түрлі дәрежедегі әр түрлі түрдің балдырлары төмендеу кезінде ауа ортасының әсерінен душар етіледі. Сондықтан олар кеуіп кетуден қорғануы тиіс. Одан басқа төмендеуден кейін қалған суық теңіз толқындары қыздырылған шалшықтарға құйылған кезде температурада кенеттен өзгеріп кетуі де мүмкі. Өсімдіктер тағы бір факторға, атап айтқанда, судың тұздылығының кенеттен өзгерісіне, яғни, оның төмендеуден кейін пайда болатын кішкентай қоймалардан буланудың артуына неболмаса оның жаңбыр кезіндегі төмендеуіне бейім болу керек., соқпа мен соққы толқындар сияқты факторларға қарсы тұру үшін механикалық беріктік қажет. Үлкен толқындар балдырларға үлкен зақым келтіре отырып, оларды жаншитын тастарды аунатып жіберуге қабілетті болып келеді.
Хлорелла
Хлорелла – біржасушалы қозғалмайтын жасыл балдыр. Оның 2.30 суретінде көрсетілген. Оны тұщы сулы тағамдарда және арықтарда кездестіруге болады. Хлорелланы оңай дақылдауға болады және ол фотосинтезді зерттеу эксперименттерінде кеңінен қолданылады (7.6 бөлім), сонымен қатар қорекктің баламалы көзі ретінде (12.12.3 бөлімі; біржасуашалы ақуыздар).
2.6.5. Phaeophyta бөлімі (қоғыр балдырлар)
Фукус
Фукус – күрделі құрылымды ірі қоңыр балдыр болып келеді. Оның денесі қысқа шыбыққа, негізгі дөңгелекке және мүкке бөлінген (бұл жалған сабақ, тамыр және жапырақтар екенін айта кету керек) өзін талломмен көрсетіледі. Бұл теңіз балдыры жартасты Британ жағалауында жиі кездеседі. ол жағалаудың кезектесіп құрғақ жерінің артуына және кемуіне, бірде жалаңаштануына, бірде сумен жабылуына бейімделген. Фукустың 3 түрлі зоналарда немесе тереңдіктерде кең таралған 3 түрі белгілі, жағалаудағы аймақтық бөліну деп аталады (10.6.4 бөлімі). Балдырларды аймаққа байланысты бөлу олардың ауада қаншалықты бола алатынына байланысты болып келеді. Төменде оларды қалай тану керектігі туралы негізгі белгілер көрсетілген, сонымен қатар, олардың таралу орындары көрсетілген.
F. spiralis (тегіс балдыр) – оларды жағалаудың ең биік жоғарылаудың нүктесіне лақтырады. Таллом батқан жағдайда шиыршыққа оңайлықпен бұрала алады.
F. serratus (қарапайым тісті немесе ара тісі тәрізді балдыр) – жартылай жоғарылау аймағында. Таллом шеті кетілдірілген.
F. vesicullosus (көпіршікті балдыр) – төмендеудің ең төмен белгісі. Ауа көпіршіктері бар жүзгіш балдырлармен қамтамасыз етілген 2.31 суретінде F. vesicullosus – тың сыртқы белгілері көрсетілген.
ҚОРШАҒАН ОРТАҒА БЕЙІМДЕЛГІШТІК. Фукустың қорек ортасына бейімделгіштігін айтпастан бұрын, оның айтарлықтай қонақжай емес туралы табиғаты туралы айта кету керек. жоғары – төмен аймағының өсімдіктері бола тұра, әр түрлі дәрежедегі әр түрлі түрдің балдырлары төмендеу кезінде ауа ортасының әсерінен душар етіледі. Сондықтан олар кеуіп кетуден қорғануы тиіс. Одан басқа төмендеуден кейін қалған суық теңіз толқындары қыздырылған шалшықтарға құйылған кезде температурада кенеттен өзгеріп кетуі де мүмкі. Өсімдіктер тағы бір факторға, атап айтқанда, судың тұздылығының кенеттен өзгерісіне, яғни, оның төмендеуден кейін пайда болатын кішкентай қоймалардан буланудың артуына неболмаса оның жаңбыр кезіндегі төмендеуіне бейім болу керек., соқпа мен соққы толқындар сияқты факторларға қарсы тұру үшін механикалық беріктік қажет. Үлкен толқындар балдырларға үлкен зақым келтіре отырып, оларды жаншитын тастарды аунатып жіберуге қабілетті болып келеді.
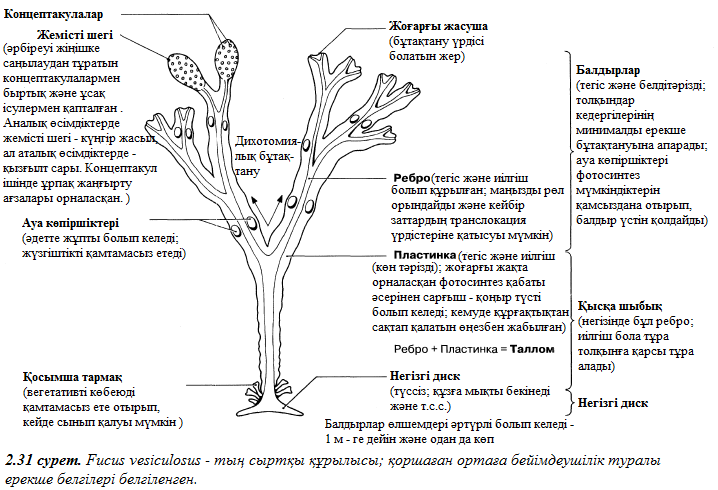
МОРФОЛОГИЯЛЫҚ БЕЙІМДЕЛГШТІКТЕР (ОРТАҚ ҚҰРЫЛЫС). Таллом балдырлары су түбіне негізгі дөңгелек арқылы берік бекінеді (2.31 сурет). Субстратпен байланыс бұл әдетте тастар, олар өте берік болып келеді, тіпті балдырдыңодан айырылуы өте қиын. Шын мәнісінде бірінші әдеттегідей тас емес,базальды диск те төзе алмайды.
Таллом жихомиялық түрде бұтақтанады (яғни, ол әрбір бұтақтану нүктесінде 2 бұтақ құрайды). Бұл 2 бұтақ арасындағы ағын суына минималды қарсылыққа әкеледі. Оның үстіне, таллом берік және иілгіш, бірақ қатты емес, ал балдырдың қабырғасы берік және солқылдақ.
F. vesicullosus – та оның түзгіштігін қамтамасыз ететін ауа көпіршіктері бар; бұл балдырды су бетінде жақын жерде ұстайды, яғни фотосинтез үшін максималды әсерді қамтамасыз жоғарысында орналасқан.
Фотосинтетикалық пигменттердің арасында қоңыр пигмент фукоксантин басым болады. Мұнда су астындағы фотосинтезге бейімделгіштік пайда болады, сондықтан фукоксантин үлкен ұзын тасқынды жарыққа қарағанда, мысалы, қызыл, су қалыңдығына едәуір әрі қарай кіретін көк аймақтарда көзге көрінетін спектр мен жарықты қатты жұтады.
Таллом балдыр қуысының барлық ішкі жағын толтыратын және оны жоғарыға бөлетін өңездердің басым көпшілігін құпиялайды.
Өңездер төмендеу кезінде сусыздандыруға қармсы тұрып, суды ұстауға көмектеседі.
Балдырлар жасушасының осмотикалық потенциалы теңіз суының осмотикалық потенциалына қарағанда жоғарылау (сәл теріс) және сондықтан осмосқа қатысты судың жоғалуы бұл жерде болмайды.
ЖЫНЫСТЫҚ КӨБЕЮГЕ ҚАБІЛЕТТІЛІК. Фукуста гаметтің босатылуы жоғарылаумен синхрондалған. Таллом төмендеу кезінде құрғап кетеді және өңездің кеіп кетуінен қорғанған репродуктивті ағзалардың сыртын сығылады. Жоғарылау кезінде репродуктивті ағзалардың қабырғалары гаметаларды босата отырып, ериді.
Аталық гаметалар қозғалғыш және оларда өздерін химиялық құпия жаққа қарай аналық гаметалар бөлінетін ауыстырып қамтамасыз ететін оң хемотаксистер бар.
Зиогаталардың дамуы ұрықтанужан кейін болады. Бұл мұхитты шайып кетуге дейін минималды қауіпке әкеледі.
2.6.6. Қарапайымдылар
Балдырлар сияқты қарапайымдылар да протиптер сияқты үлкен топты құрайды. Бұл біржасушашылар гетеротрофты қоректенетін жануарларға тән болып келедіү су бар жерде барлығын көрсететін 50 000 белгілі түрі бар. Олардың көбісі – қозғалудың әртүрлі қабілетіне байланысты сипатталатын еркін тіршілік ететін ағзалар. Кейбіреулері, паразиттер болып табылады. бұл, сондай – ақ, Plasmodium, бұл басқаларына қарағанда көп адам өлтірген бағасы бар ауру шақыратын паразит. Бұл ауру – малярия – бұл әлі де әлемде ең орқынышты өлтіруші болып қалмақ.
Біз қарапайымдылар зерттеуінде мысал ретінде еркін тіршілік етуші ағза – туфлька инфузориясын алдық (Paramecium caudatum), сондықтан ол бұл ұжым дәрежесіне типті. Безгек плазмодийі 15 бөлімде қарастырылады.
2.6.7. Ciliphora бөлімі (кірпікшелілер)
Кірпікшелілер – бұл қарапайымдылардың бір типі (2.28 сурет). Олар келесі белгілерге тән:
Пеликулла астында жылтыр тығыз цитоплазманың гель консистенциясы – эктоплазма орналасқан. Эктоплазмада кірпікшелілер табылмайтын негізгі түйіршіктер бар (центриоьдері ұқсас), ал негізгі түйіршіктер арасында кірпікшелер дүрсілінің үйлестірілуіне қатысатын кішкене фибриллдердің байланысы бар.
Таллом жихомиялық түрде бұтақтанады (яғни, ол әрбір бұтақтану нүктесінде 2 бұтақ құрайды). Бұл 2 бұтақ арасындағы ағын суына минималды қарсылыққа әкеледі. Оның үстіне, таллом берік және иілгіш, бірақ қатты емес, ал балдырдың қабырғасы берік және солқылдақ.
F. vesicullosus – та оның түзгіштігін қамтамасыз ететін ауа көпіршіктері бар; бұл балдырды су бетінде жақын жерде ұстайды, яғни фотосинтез үшін максималды әсерді қамтамасыз жоғарысында орналасқан.
Фотосинтетикалық пигменттердің арасында қоңыр пигмент фукоксантин басым болады. Мұнда су астындағы фотосинтезге бейімделгіштік пайда болады, сондықтан фукоксантин үлкен ұзын тасқынды жарыққа қарағанда, мысалы, қызыл, су қалыңдығына едәуір әрі қарай кіретін көк аймақтарда көзге көрінетін спектр мен жарықты қатты жұтады.
Таллом балдыр қуысының барлық ішкі жағын толтыратын және оны жоғарыға бөлетін өңездердің басым көпшілігін құпиялайды.
Өңездер төмендеу кезінде сусыздандыруға қармсы тұрып, суды ұстауға көмектеседі.
Балдырлар жасушасының осмотикалық потенциалы теңіз суының осмотикалық потенциалына қарағанда жоғарылау (сәл теріс) және сондықтан осмосқа қатысты судың жоғалуы бұл жерде болмайды.
ЖЫНЫСТЫҚ КӨБЕЮГЕ ҚАБІЛЕТТІЛІК. Фукуста гаметтің босатылуы жоғарылаумен синхрондалған. Таллом төмендеу кезінде құрғап кетеді және өңездің кеіп кетуінен қорғанған репродуктивті ағзалардың сыртын сығылады. Жоғарылау кезінде репродуктивті ағзалардың қабырғалары гаметаларды босата отырып, ериді.
Аталық гаметалар қозғалғыш және оларда өздерін химиялық құпия жаққа қарай аналық гаметалар бөлінетін ауыстырып қамтамасыз ететін оң хемотаксистер бар.
Зиогаталардың дамуы ұрықтанужан кейін болады. Бұл мұхитты шайып кетуге дейін минималды қауіпке әкеледі.
2.6.6. Қарапайымдылар
Балдырлар сияқты қарапайымдылар да протиптер сияқты үлкен топты құрайды. Бұл біржасушашылар гетеротрофты қоректенетін жануарларға тән болып келедіү су бар жерде барлығын көрсететін 50 000 белгілі түрі бар. Олардың көбісі – қозғалудың әртүрлі қабілетіне байланысты сипатталатын еркін тіршілік ететін ағзалар. Кейбіреулері, паразиттер болып табылады. бұл, сондай – ақ, Plasmodium, бұл басқаларына қарағанда көп адам өлтірген бағасы бар ауру шақыратын паразит. Бұл ауру – малярия – бұл әлі де әлемде ең орқынышты өлтіруші болып қалмақ.
Біз қарапайымдылар зерттеуінде мысал ретінде еркін тіршілік етуші ағза – туфлька инфузориясын алдық (Paramecium caudatum), сондықтан ол бұл ұжым дәрежесіне типті. Безгек плазмодийі 15 бөлімде қарастырылады.
2.6.7. Ciliphora бөлімі (кірпікшелілер)
Кірпікшелілер – бұл қарапайымдылардың бір типі (2.28 сурет). Олар келесі белгілерге тән:
- Бұл гетеротрофты формасындағы біржасушашылар;
- Олар кішкентай түкшелі өсінділері бар,су тогын шақыртатын соққысы бар, жасушаны не өзінің қозғалысына немесе оның қорек бөліктерін алуға қолданылатын кірпікшелермен қамдалған.
- Олар целикула деп аталатын цитоплазманың кіші иілгіш сыртқы қабатымен қамтамасыз етілген нақты формасымен сипатталады.
- Кірпішелі жасушалары айтарлықтай күрделі құрылымды болып келеді, мысалы, оларда макронуклеус және микронуклеус деген бар.
Пеликулла астында жылтыр тығыз цитоплазманың гель консистенциясы – эктоплазма орналасқан. Эктоплазмада кірпікшелілер табылмайтын негізгі түйіршіктер бар (центриоьдері ұқсас), ал негізгі түйіршіктер арасында кірпікшелер дүрсілінің үйлестірілуіне қатысатын кішкене фибриллдердің байланысы бар.
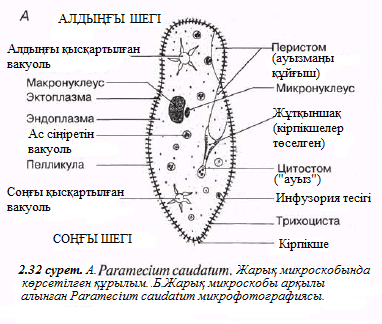
|
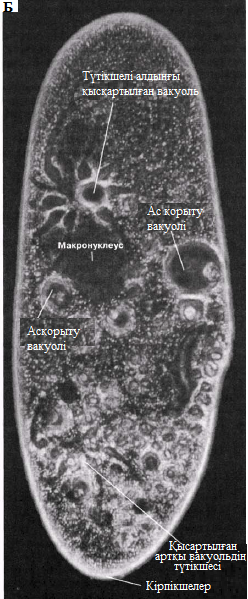
|
Эндоплазманың негізгі массасы эктоплазмаға ұарағанда көп бөлігі сұйық консистенциямен толған эндоплазма құрайды. Дәл эндоплазмада органелланың көп бөлігі орналасқан. Туфельканың құрсақтұс жағының жоғарғы жағында және оның алдыңғы жағының соңғысында ауызмаңы құйғыш болады, оның түбінде ауыз не болмаса цитостом орналасқан. Ауыз кішігірім канал – цитофаринкске немесе жұтқыншаққа апарады. Ауызмаңы құйғыш сияқты су қоймасына қарай бағытталатын қозғалыс, өзімен бірге әртүрлі қорек бөліктерін тасып жүретін бактериялар сияқты жұтқыншақ та кірпікшелермен төселе алады. Цитоплазма айналасына экзоцитоз жолымен қорытылмаған астарды сыртқа шығарылатын инфузория тесігі ауысады.
Цитоплазмада тағы жасушада орны дәл белгіленіп қойған қысқарған вакуольдер бар (2.32 сурет). Бұл вакуольдер осморегуляцияға жауап береді, яғни жасушаның су потенциалын ұстап тұрады (20 бөлім). Тұщы судағы өмірде су жасушаға осмос нәтижесінде енеді, осысымен бұл өмір қиын болып келеді, бұл су жарылыстың алдын алу үшін жасушадан үздіксіз іске аспай қалып отыру керек.
Бұл энергия жоғалтуды көздейтін белсенді транспорт процесі арқылы жүреді. Әрбір қысқарған вакуольде айналасында вакуольге қарай босатпастан бұрын жинап алатын айырылғыш сәулелер өзектерінің қатарымен орналасқан.
Перемеция жасушасында 2 ядро бар. Оның үлкені – полиплоидты – макронуклеус; екіден астам хромосомасы бар және көбеюмен қатысы жоқ зат алмасу үрдістерін басқарады. Микронуклеус – диплоидты ядро. Ол көбеюді және ядро бөлінуіндегі – макронуклеусердің түзілуін басқарады.
Парамеция жыныссыз (көлденең екіге бөліну) және жынысты (конъюгация жолымен) болып көбейе алады.
2.6.8. Apicomplexa бөлімі
бұл топтың қарапайымдылары жасушаға белгілі бір пішін беретін пелликула бар. Бірақ олардың басым көпшілігінде қозғалуына арнайы құрылым жоқ. Apicomplexa – ның айырмашылығы – споралардың түзілуінде және жынысты көбеуінде болып табылады. бұл қарапайымдыларға мысал ретінде қызмет ететін адамда малярия қоздырғышы болып табылатын және 15 бөлімде қарастырылатын Plasmadium паразиті.
2.7. Өсімдіктер патшалығы
Біздің жер шарымыз 3,5 млрд. жыл бұрын туды, ал алғашқы ағзалар жерге 420 млн. жыл бұрын қоныстанып бастады. Бұл ең бірінші өсімдіктер еді. Өсімдіктер – ауа ортасына бейімделген автотрофты эукариоттар. Су өміріне мамандандырылған өзге автотрофтар арасындағы жалғыз эукариот – балдырлар. Еске саламыз: автотрофтар деп – органикалыз заттар синтезі үшін бейорганикалық көміртегін, атап айтқанда көміртек диоксидін қайнар көзі ретінде қолданатын ағзалар. Бірақ, сонымен қатар, мұндай синтез үшін көміртек көзі ретінде энергия мен өсімдік қажет етіледі, фотоавтотрофтытар бола тұра қайнар көзі ретінде жарық энергиясын да қолданады. Бұл қоректену қабілетін фотосинтез деп атау қабылданды.
Өсімдік эволюциясының тарихы – бұл құрғақ жердің тіршілігіне бейімделгішдік жолмен үйрету. Және дәл осы тақырып біздің өсімдік зерттеуіміз де маңызды болып табылады. өсімдік систематикасы берілген кітапта 2.33 суретте көрсетілген. Ол жерде бұл бөлімде қарастырылатын өсімдік эволюциясының кейбір маңызды үрдістердің кішігірім тізбегі мен құрғақ жердегі бейімделгіш туралы қарастырылады.
2.7.1. Bryophyta бөлімі (бауырмүктәрізділер және мүктер)
Бауырмүктер мен мүктер – жер өсімдіктеріне қарағанда қарапайымдылау болып келеді. Олар жасыл балдырлардан пайда болған деген болжамдар бар. Bryophyta (мүк тәрізділер) бөліміне 2 негізгі класс кіреді – Hepiticae (бауыр мүктер) және Musci (мүктер). Бұл 2 топ та жерде өмірге қабілеттері нашар және сол үшін олар дымқыл көлеңкелі жерлерге таңылған. Мүк тәрізділердің систематикасы мен негізгі белгілері 2.8 кестесінде келтірілген.
Цитоплазмада тағы жасушада орны дәл белгіленіп қойған қысқарған вакуольдер бар (2.32 сурет). Бұл вакуольдер осморегуляцияға жауап береді, яғни жасушаның су потенциалын ұстап тұрады (20 бөлім). Тұщы судағы өмірде су жасушаға осмос нәтижесінде енеді, осысымен бұл өмір қиын болып келеді, бұл су жарылыстың алдын алу үшін жасушадан үздіксіз іске аспай қалып отыру керек.
Бұл энергия жоғалтуды көздейтін белсенді транспорт процесі арқылы жүреді. Әрбір қысқарған вакуольде айналасында вакуольге қарай босатпастан бұрын жинап алатын айырылғыш сәулелер өзектерінің қатарымен орналасқан.
Перемеция жасушасында 2 ядро бар. Оның үлкені – полиплоидты – макронуклеус; екіден астам хромосомасы бар және көбеюмен қатысы жоқ зат алмасу үрдістерін басқарады. Микронуклеус – диплоидты ядро. Ол көбеюді және ядро бөлінуіндегі – макронуклеусердің түзілуін басқарады.
Парамеция жыныссыз (көлденең екіге бөліну) және жынысты (конъюгация жолымен) болып көбейе алады.
2.6.8. Apicomplexa бөлімі
бұл топтың қарапайымдылары жасушаға белгілі бір пішін беретін пелликула бар. Бірақ олардың басым көпшілігінде қозғалуына арнайы құрылым жоқ. Apicomplexa – ның айырмашылығы – споралардың түзілуінде және жынысты көбеуінде болып табылады. бұл қарапайымдыларға мысал ретінде қызмет ететін адамда малярия қоздырғышы болып табылатын және 15 бөлімде қарастырылатын Plasmadium паразиті.
2.7. Өсімдіктер патшалығы
Біздің жер шарымыз 3,5 млрд. жыл бұрын туды, ал алғашқы ағзалар жерге 420 млн. жыл бұрын қоныстанып бастады. Бұл ең бірінші өсімдіктер еді. Өсімдіктер – ауа ортасына бейімделген автотрофты эукариоттар. Су өміріне мамандандырылған өзге автотрофтар арасындағы жалғыз эукариот – балдырлар. Еске саламыз: автотрофтар деп – органикалыз заттар синтезі үшін бейорганикалық көміртегін, атап айтқанда көміртек диоксидін қайнар көзі ретінде қолданатын ағзалар. Бірақ, сонымен қатар, мұндай синтез үшін көміртек көзі ретінде энергия мен өсімдік қажет етіледі, фотоавтотрофтытар бола тұра қайнар көзі ретінде жарық энергиясын да қолданады. Бұл қоректену қабілетін фотосинтез деп атау қабылданды.
Өсімдік эволюциясының тарихы – бұл құрғақ жердің тіршілігіне бейімделгішдік жолмен үйрету. Және дәл осы тақырып біздің өсімдік зерттеуіміз де маңызды болып табылады. өсімдік систематикасы берілген кітапта 2.33 суретте көрсетілген. Ол жерде бұл бөлімде қарастырылатын өсімдік эволюциясының кейбір маңызды үрдістердің кішігірім тізбегі мен құрғақ жердегі бейімделгіш туралы қарастырылады.
2.7.1. Bryophyta бөлімі (бауырмүктәрізділер және мүктер)
Бауырмүктер мен мүктер – жер өсімдіктеріне қарағанда қарапайымдылау болып келеді. Олар жасыл балдырлардан пайда болған деген болжамдар бар. Bryophyta (мүк тәрізділер) бөліміне 2 негізгі класс кіреді – Hepiticae (бауыр мүктер) және Musci (мүктер). Бұл 2 топ та жерде өмірге қабілеттері нашар және сол үшін олар дымқыл көлеңкелі жерлерге таңылған. Мүк тәрізділердің систематикасы мен негізгі белгілері 2.8 кестесінде келтірілген.
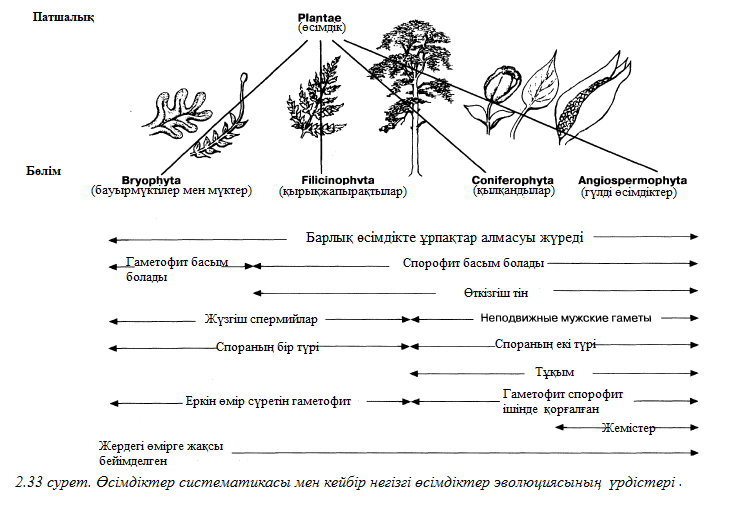
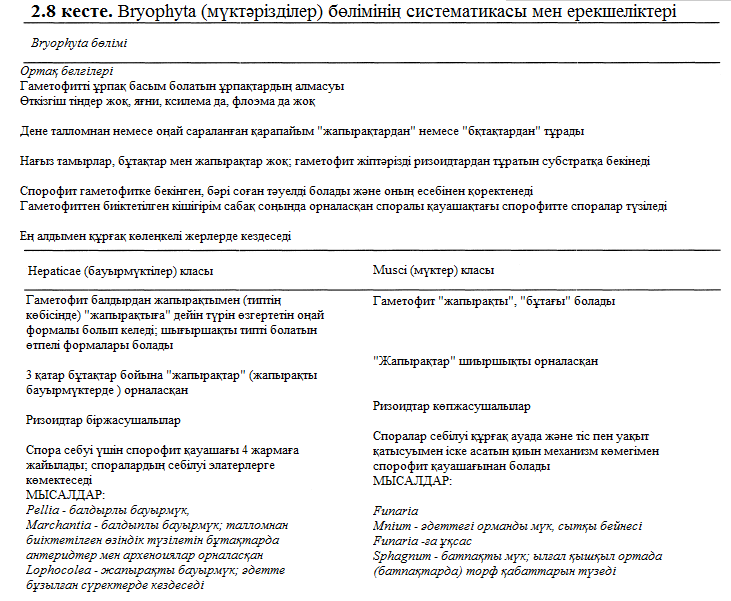
Bryophyta – бұл жеңіл құрылымды кішігірім өсімдіктер. Олардың дамуындағы тіректі және өткізгіш тіндер әлсіз немесе мүлдем болмайды. Оларда сараланған ксилемалар мен флоэмалар, сонымен қоса нағыз тамырларда жоө. Жер бетінде олар ризоидтар деп аталатын аса нәзік жіптермен ұсталып тұр. барлық дененің жоғарысы су және минералды тұздар, сонымен қоса, ризоидтар да сіңіріледі. Ал бұл ризоидтардың нағыз тамырларының айырмашылығы : жердегі өсімдіктің бекінуіне қызмет етеді. (Нағыз тамырларда және нағыз сабақтарда немесе жапырақтардаөткізуші тіндер болады). Мүктәрізділердің үстері сірқабықпен жабылмаған, сірқабықтап жіп – жіңішке болуы соншалық, олар судың жоғалуына немесе кіруіне тосқауыл бола алмайды. Дегенмен, көптеген мүктәрізділер мүлдем түсініксіз механизмдерді қолдана отырып, құрғақ кезеідер ұстануға қабілетті болды. Мысалы, Grimmia pulvinata сияқты белгілі құрғақсүйгішті мүк 20 - та жыл бойы шынымен құрғақ жағдайда тірі қала алады деп анықталған. Өсімдік ылғалды ортаға түскен кезінен бастап, ода барлық функциялары бірден қондырылады.
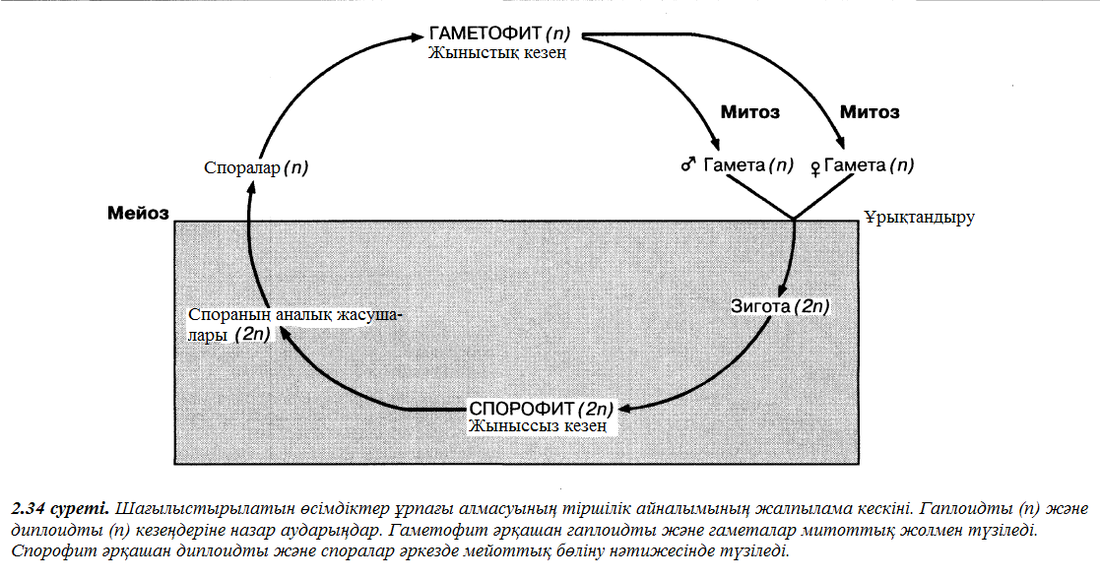
Ұрпақтар алмасуы
Барлық өсімдіктер және кейбір ламинария сияқты жоғары ұйымдасқан балдырлар және мүктәрізділер сияқтыларда ұқрпақтар алмасулары байқалады. Тіршілік кезең үстінде 2 типті ағзаның ауысымы жүреді: гаплоидты гаметофит ұрпағымен диплоидты спорофиттің ұрпағы кезектесе отырып ауысады, схемалары 2.34 суретінде көрсетілген. Гаплоидты ұрпақ жыныстық көбеюге және гаметалар түзуге қабілетті гаметофит (грек. gamet – аналық, gametē – аталық, phytόn - өсімдік) деп аталады.сондықтан гаметалар түзілуі митоз нәтижесінде болады, олар да гаплоидты. Гаметалардың қосылуынанкелесі ұрпақ диплоидты спорофит ұрпақ деп аталатындиплоидты зиогталар құрылады. Олар спорофиттер деп аталады, себебі олар споралардың түзілуімен жыныссыз көбеюге қабілетті. Споралар мейоз нәтижесінде түзіледі, яғни мұнда гаплоиты жағдайға қайтарылым жүреді. Галоидты споралар гаметофитті ұрпаққа бастапқысып береді. Бұл 2 ұрпақтықбіреуі әрқашан өзгелерді басөарады және оның үлесіне үлкен тіршілік кезеңдері келеді бұл ұрпақ доминантты деп аталады. мүктәрізділерді гаметофит ұрпағы доминанттайды, ал басқа жер өсімдіктерін – спорофиттер ұрпағы тіршілік кезеңі кескінінде доминантты ұрпақ сызбада жоғары бөлігінде ауысу қабылданды.
2.34 суретін мұқият зерттеңдер, онда жер өсімдіктері мен әбден жоғары бірлескен гүлді өсімдіктеріні жалпылама түрде тіршілік кезеңдері көрсетілген. Өсімдік гаметалары жануарлардыкі секілді мейоз нәтижесінде емесе, митоз нәтижесінде түзілетінін ешқашан ұмытпаңдар; спор түзілуінде мейоттық бөліну жүреді.
Hepiticae класы - бауырмүктілер
Hepiticae класының сипатталатын белгілері 2.8 кестесінде берілген. Бауырмүктердің өзіндік құрылыстары мүктерге қарағанда тіптен оңай және олар жалпылама дымқыл және көлеңкелі жерге тіркеулі болып келеді. Оларды өзен, бұлақтарда, дымқыл тастарда және қалқанды өсімдіктер арасында табуға болады. Көптеген бауырмүктерде дұрыс үлестер немесе толықтай кішігірім қарапайым жапырақтар мен көрсетілген бұтақтар көрінеді. Бәрінен де жауынмүкті және бұтақ пен жапыраққа бөлінбеген тіптен оңай құрылған талломды бауырмүктер болып табылады.
Барлық өсімдіктер және кейбір ламинария сияқты жоғары ұйымдасқан балдырлар және мүктәрізділер сияқтыларда ұқрпақтар алмасулары байқалады. Тіршілік кезең үстінде 2 типті ағзаның ауысымы жүреді: гаплоидты гаметофит ұрпағымен диплоидты спорофиттің ұрпағы кезектесе отырып ауысады, схемалары 2.34 суретінде көрсетілген. Гаплоидты ұрпақ жыныстық көбеюге және гаметалар түзуге қабілетті гаметофит (грек. gamet – аналық, gametē – аталық, phytόn - өсімдік) деп аталады.сондықтан гаметалар түзілуі митоз нәтижесінде болады, олар да гаплоидты. Гаметалардың қосылуынанкелесі ұрпақ диплоидты спорофит ұрпақ деп аталатындиплоидты зиогталар құрылады. Олар спорофиттер деп аталады, себебі олар споралардың түзілуімен жыныссыз көбеюге қабілетті. Споралар мейоз нәтижесінде түзіледі, яғни мұнда гаплоиты жағдайға қайтарылым жүреді. Галоидты споралар гаметофитті ұрпаққа бастапқысып береді. Бұл 2 ұрпақтықбіреуі әрқашан өзгелерді басөарады және оның үлесіне үлкен тіршілік кезеңдері келеді бұл ұрпақ доминантты деп аталады. мүктәрізділерді гаметофит ұрпағы доминанттайды, ал басқа жер өсімдіктерін – спорофиттер ұрпағы тіршілік кезеңі кескінінде доминантты ұрпақ сызбада жоғары бөлігінде ауысу қабылданды.
2.34 суретін мұқият зерттеңдер, онда жер өсімдіктері мен әбден жоғары бірлескен гүлді өсімдіктеріні жалпылама түрде тіршілік кезеңдері көрсетілген. Өсімдік гаметалары жануарлардыкі секілді мейоз нәтижесінде емесе, митоз нәтижесінде түзілетінін ешқашан ұмытпаңдар; спор түзілуінде мейоттық бөліну жүреді.
Hepiticae класы - бауырмүктілер
Hepiticae класының сипатталатын белгілері 2.8 кестесінде берілген. Бауырмүктердің өзіндік құрылыстары мүктерге қарағанда тіптен оңай және олар жалпылама дымқыл және көлеңкелі жерге тіркеулі болып келеді. Оларды өзен, бұлақтарда, дымқыл тастарда және қалқанды өсімдіктер арасында табуға болады. Көптеген бауырмүктерде дұрыс үлестер немесе толықтай кішігірім қарапайым жапырақтар мен көрсетілген бұтақтар көрінеді. Бәрінен де жауынмүкті және бұтақ пен жапыраққа бөлінбеген тіптен оңай құрылған талломды бауырмүктер болып табылады.
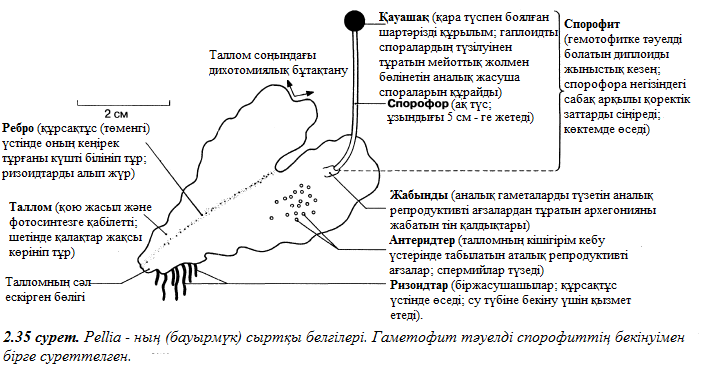
Мысал ретінде Pellia бауырмүгін алуға болады, ол Ұлыбританияда кеңінег таралған болатын. Бұл өсімдік күңгірт жасыл түспен боялған тегіс бұтақтың ұзындығы, шамамен, 1 см-ді құрайды. Pellia – ның сыртқы белгілері 2.35 суретінде көрсетілген.
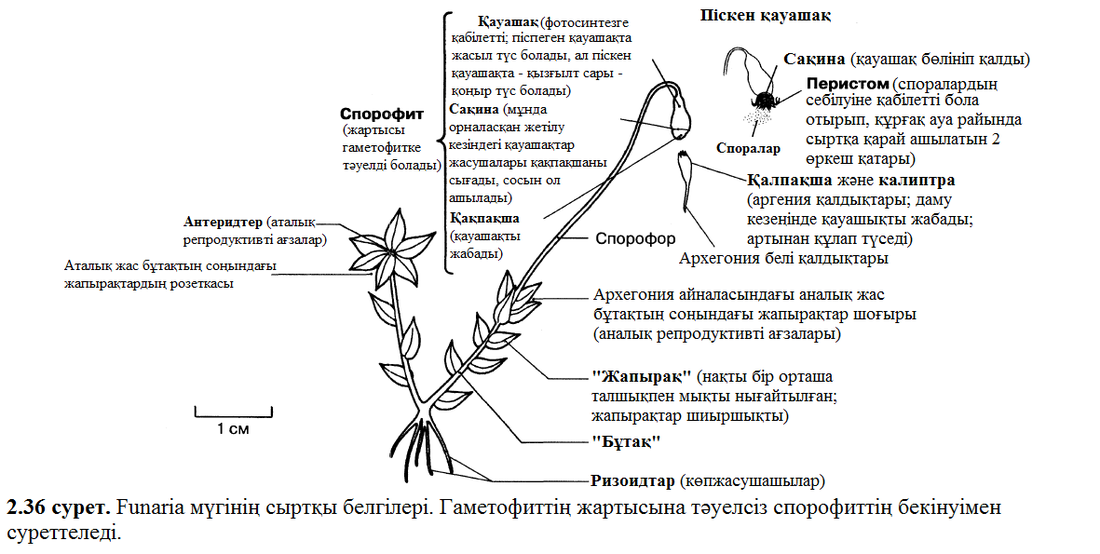
Musci класы -жапырақсабақты мүктер
2.8 кестесінде жапырақсабақты мүктердің негізгі белгілері көрсетілген. Жапырақсабақты мүктер бауырмүктілерге қарағанда көп сараланған, бірақ, бауырмүктілер сияқты дымқыл орталарда негізгі кейіппен кездесетін кішігірім өсімдіктер болып табылады. Олар жиі тығыз көпшіктер құрайды.
Funaria – әдеттегі алғашқылардың бірі болып егінге, кесілген жерге және шаппа орынға жайғастырылатын жапырақсабақтылардың бір өкілі. Funaria әсіресе арпабас өртенген және қаулаған өртте орналасқанды жақсы көреді. Бұл жылыжай және бақтарда болатын ең қарапайым арамшөптердің ьірі. 2.36 суретінде Funaria – ның сыртқы көрінісі берілген.
Бауырмүктер сияқты Funaria – да ұрықтану үшін суды қажет етеді. Таллом үсті ылғалданғанда, піскен антеридтер суды сіңіреді және үстіне аталық гаметаларды (спермийлар) босата отырып, жарылады. Әрбір спермий екі жалған аяқпен жабдықталған. Спермийлар әрбіреуінде аналық гамета немесе жұмыртқа жасушасы болатын архегонияныңастына кіріп кетіп отырады. Ұрықтану, яғни спермий ядросымен жұмыртқа жасушасының бірігуі архегонияда жүреді. Бірігу нәтижесінде, жаңа спорофит туындататын архегонияда өскен диплоидты зигота түзіледі.
2.7.2. Filicinophyta бөлімі (қырыққұлақтәрізділер)
Filicinophyta – ның негізгі белгілері 2.9 кестесінде берілген. Қырыққұлақ көлеңкелі дымқыл жерлерде кездеседі. кейбір қырыққұлақтылар ашық жерде өсуі мүмкін, тіпті ең қарапайым қыранат та ерекшелігін құрайды. Қырыққұлақтәріздестер температурасы, жарықтылығы және ылғалдылығы келіскен тропикалық жаңбырлы ормандарда кең таралған.
Қырыққұлақтар – тамырлы өсімдіктер; басқа сөзбен айтқанда оларда өсімдік бойында қорек заттары және су транслакация функциясын орындайтын ксилемалар мен флоэмалардан тұратын өткізгіш тін болады. Ксилема тамырлары бойынша негізінде су мен минералды тұздар тасымалы болады, онда флоэма сияқты болып сахара сияқты ең алдымен органикалық заттар ерітінділері ауыстырады. Өткізгіш тіндер кейбір мүктәрізділер мен балдырлардың қарапайым өткізгіш жасушалармен салыстырып қарағанда салмақты эволюционды пайда әкелетін болып табылады. Бұл тіндер спорофитті ұрпақтарда анықталған болатын, міне, дәл осы үшін спорофиттер басқа да барлық тамырлы өсімдіктерге доминантты болып табылады.
2.8 кестесінде жапырақсабақты мүктердің негізгі белгілері көрсетілген. Жапырақсабақты мүктер бауырмүктілерге қарағанда көп сараланған, бірақ, бауырмүктілер сияқты дымқыл орталарда негізгі кейіппен кездесетін кішігірім өсімдіктер болып табылады. Олар жиі тығыз көпшіктер құрайды.
Funaria – әдеттегі алғашқылардың бірі болып егінге, кесілген жерге және шаппа орынға жайғастырылатын жапырақсабақтылардың бір өкілі. Funaria әсіресе арпабас өртенген және қаулаған өртте орналасқанды жақсы көреді. Бұл жылыжай және бақтарда болатын ең қарапайым арамшөптердің ьірі. 2.36 суретінде Funaria – ның сыртқы көрінісі берілген.
Бауырмүктер сияқты Funaria – да ұрықтану үшін суды қажет етеді. Таллом үсті ылғалданғанда, піскен антеридтер суды сіңіреді және үстіне аталық гаметаларды (спермийлар) босата отырып, жарылады. Әрбір спермий екі жалған аяқпен жабдықталған. Спермийлар әрбіреуінде аналық гамета немесе жұмыртқа жасушасы болатын архегонияныңастына кіріп кетіп отырады. Ұрықтану, яғни спермий ядросымен жұмыртқа жасушасының бірігуі архегонияда жүреді. Бірігу нәтижесінде, жаңа спорофит туындататын архегонияда өскен диплоидты зигота түзіледі.
2.7.2. Filicinophyta бөлімі (қырыққұлақтәрізділер)
Filicinophyta – ның негізгі белгілері 2.9 кестесінде берілген. Қырыққұлақ көлеңкелі дымқыл жерлерде кездеседі. кейбір қырыққұлақтылар ашық жерде өсуі мүмкін, тіпті ең қарапайым қыранат та ерекшелігін құрайды. Қырыққұлақтәріздестер температурасы, жарықтылығы және ылғалдылығы келіскен тропикалық жаңбырлы ормандарда кең таралған.
Қырыққұлақтар – тамырлы өсімдіктер; басқа сөзбен айтқанда оларда өсімдік бойында қорек заттары және су транслакация функциясын орындайтын ксилемалар мен флоэмалардан тұратын өткізгіш тін болады. Ксилема тамырлары бойынша негізінде су мен минералды тұздар тасымалы болады, онда флоэма сияқты болып сахара сияқты ең алдымен органикалық заттар ерітінділері ауыстырады. Өткізгіш тіндер кейбір мүктәрізділер мен балдырлардың қарапайым өткізгіш жасушалармен салыстырып қарағанда салмақты эволюционды пайда әкелетін болып табылады. Бұл тіндер спорофитті ұрпақтарда анықталған болатын, міне, дәл осы үшін спорофиттер басқа да барлық тамырлы өсімдіктерге доминантты болып табылады.

Өткізгіш тіндер 2 маңызды функцияны орындайды. Біріншіден, олар ірі күрделі денені дамыта отырып, көпжасушалы дегенге су мен минералды тұзді алмастыратын транспортты жүйені құрайды. Екіншіден, олар тіректі функцияны орындайды, сондықтан ксилемалар, өткізгіш тіндері бола тұра, өте мықты және қатты болатын лигнификацияланған жасушалар - склеренхиманы құрайды (6.2.1. бөлім).
Спорофитті ұрпақта ағыз тамыр, бұтақтар әне жапырақтар болады. Тамырлар жерге ене отырып, өсімдікке су мен ерітілген заттардың кіруін жеңілдетеді. Олар ксилема бойынша өсімдіктің басқа да бөліктері арқылы енеді. Өсімдік бойлары қолдау тапқан бойда, олар жер бетінің үстіне шығып артады,олар жарық үшін бірден бәсекелестік тудыруға міндетті болу керек еді және нәтижесінде ұзын көлемділердің көп дамуы үрдіі пайда болар еді. Қырыққабаттылар және ағаштәрізді қырыққабаттылар құрғақтықта шамамен девон кезеңінен перм кезеңіне дейін 70 млн, жыл бойы доминантты болып келді. Содан кейін олар айтарлықтай дәрежеде бастапқыда қылқандығы ығыстырылып шығып, содан кейін олар гүлді өсімдіктерге көшті (3 томдағы 4 қосымшаны қарау).
Спорофитті ұрпақта ағыз тамыр, бұтақтар әне жапырақтар болады. Тамырлар жерге ене отырып, өсімдікке су мен ерітілген заттардың кіруін жеңілдетеді. Олар ксилема бойынша өсімдіктің басқа да бөліктері арқылы енеді. Өсімдік бойлары қолдау тапқан бойда, олар жер бетінің үстіне шығып артады,олар жарық үшін бірден бәсекелестік тудыруға міндетті болу керек еді және нәтижесінде ұзын көлемділердің көп дамуы үрдіі пайда болар еді. Қырыққабаттылар және ағаштәрізді қырыққабаттылар құрғақтықта шамамен девон кезеңінен перм кезеңіне дейін 70 млн, жыл бойы доминантты болып келді. Содан кейін олар айтарлықтай дәрежеде бастапқыда қылқандығы ығыстырылып шығып, содан кейін олар гүлді өсімдіктерге көшті (3 томдағы 4 қосымшаны қарау).
Ауа ортасындағы спорофитті ұрпақтың прогрессивті бейімделгіштерінің бар екеніне қарамастан, өскін деп аталатын қырыққабаттәрізділердің гаметофиті ұлкен мәселелерге тап бола берді. Ол әзірге шейін көлемі бойынша кішілеу және мүктәрізділердің гаметофите қарағанда құрғап қалуға сәл де болсын тірей алады. Өскіндерде мүктәрізділер сияқты спермиялар түзіледі, аналық гаметаларға оларда жөзіп қана отырып жетіге болады.
Аталық усасыр (Dryopteris filix – mas)
Бұл Ұлыбританияда ең көп таралған қырыққабат болып табылады; ол барлық дымқыл орманды, орман паркі және тағы да юасқа көлеңкелі елдерді кездеседі. биіктігі 1 мге немесе одан да көп спорофит вайилары (жапырақтар) жуан көлденең бұтақтан немесе тамырсабақтан өседі. Тамырсабақта қосалқы тамырлар болады. Негізгі тамырсабақтан бөлек бұтақтар сынып қалуы мүмкін және жаңа өсімдіктерге жаіа бастау береді. Бұл – вегетативті көбеюдің бірден бір формасы. Тамырсабақтардың негізінде суықтан және құрғап қалудан қорғайтын құрғақ қоңыр қабыршақтар жабылған. Жас жапырақтар ерекше қырыққабат бұрамаларына ыңғайлы қылып, оранып алған. Жапырақ сабының жоғарғысында қабыршақ өлшемдері жаймен кішірейеді және олардық арасындағы арақашықтық ұлғаяды. Вайи сабы негізгі сап деп аталады, ал 2 жақта да жоқ жапырақтар – мамықты жапырақтар болып саналады. Жапырқтаңы кішігірім дөңес жерлері екілік жапырақтар деп аталады. Dryopteris filix – mas спорофитінің сыртқы белгілерімен 2.37 суретінде танысуға болады; спорофит 2.38 суретінде көрсетілген.
Споралар спорангийлер деп аталатын арнай құрылымды болып жаздың соңында құралады. Екілік жапырақтардың төменгі жағында ерекше спорангиялар жиынтығы деп аталатын жинақта болады (2.37. суретінде, В, Г және Д). Әрбір спорангиялар жиынтығы индузий деп аталатын қорғаныш жабындысымен қапталған. Әрбір спорангий ішінде диплойдты аналық жасушалар споралары нәтижесінде гаплоидты споралар пайда болатын мейоттық бөліну жүреді. Индузийлер пісіп жетілгенде тыржиып және қопарылып түседі, ал спорангий қабырғалары ашылған кезде құрғап бастайды. Соңында, қабырғасы жайылады және споралар спорангийден катапульта сияөты атылып шығады (2.37, Д сурет).
Споралар гаметофитті ұрпақта туындата отырып, өседі. Гемотофит диаметрі, шамамен, 1 см болатын кішігірім жүректәрізді жасуша болып табылады (2.38 сурет). Бұл пластинка жасыл түсті, фотосинтезге қабілетті және жерге біржасушалы ризоидтармен бекінеді. Сондықтан мұндай нәзік өскінде кутикулалар жоқ, ол тез құрғайды және демек, ол тек ылғалды ортада тіршілік ете алады.
Гаметофиттің (өскін) төменгі жақтарында қарапайым архегонийлар және антеридийлар бар. Бұл репродуктивті ағзалар оның құрамындағы гаметаларды қорғайды. Гаметалар аналық жыныс жасушасындағы митоздың нәтижесінде пайда болады; бұл ретте, антеридийларда мүктәрізділер сияқты спермиялар пайда болады, ал әрбір архегонияда – бір жұмыртқажасушасынан болады. Спермиялар жалған аяқтармен жабдықталған. Ылғалды жағайларда піске спермийлар антеридийдан босап, су қабыршағында архегонияға қарай жүзеді. Ұрықтану нәтижесінде, диплоидты зигота түзіледі. Қырыққабаттәрізділер де мүктәрізділер сияқты ұрықтанулары суға тәуелді екеніне назар салыңыздар.
Зиготалар спорофитті ұқпақты туындатады. Жас ұқпақ өнім заттары функциясын өзінің жапырағы мен тамырлары орындағанша, ол гаметофиттн ала тұрады (2.38, Б сурет). Гаметофит көп кешіпей солып, құриды.
2.7.3. Тұқымдық өсімдіктер
Өсімдіктің ең гүлденуші тобы ұрық түзеді. Бұл өсімдіктер, тегі, қырыққабаттылардың және олардың туғандарының құрып біткен тұқым өкілдерінен түзілген. Тұқымдық өсімдіктердің систематикасы мен негізгі белгілері 2.10 кестесінде жинақталған.
Аталық усасыр (Dryopteris filix – mas)
Бұл Ұлыбританияда ең көп таралған қырыққабат болып табылады; ол барлық дымқыл орманды, орман паркі және тағы да юасқа көлеңкелі елдерді кездеседі. биіктігі 1 мге немесе одан да көп спорофит вайилары (жапырақтар) жуан көлденең бұтақтан немесе тамырсабақтан өседі. Тамырсабақта қосалқы тамырлар болады. Негізгі тамырсабақтан бөлек бұтақтар сынып қалуы мүмкін және жаңа өсімдіктерге жаіа бастау береді. Бұл – вегетативті көбеюдің бірден бір формасы. Тамырсабақтардың негізінде суықтан және құрғап қалудан қорғайтын құрғақ қоңыр қабыршақтар жабылған. Жас жапырақтар ерекше қырыққабат бұрамаларына ыңғайлы қылып, оранып алған. Жапырақ сабының жоғарғысында қабыршақ өлшемдері жаймен кішірейеді және олардық арасындағы арақашықтық ұлғаяды. Вайи сабы негізгі сап деп аталады, ал 2 жақта да жоқ жапырақтар – мамықты жапырақтар болып саналады. Жапырқтаңы кішігірім дөңес жерлері екілік жапырақтар деп аталады. Dryopteris filix – mas спорофитінің сыртқы белгілерімен 2.37 суретінде танысуға болады; спорофит 2.38 суретінде көрсетілген.
Споралар спорангийлер деп аталатын арнай құрылымды болып жаздың соңында құралады. Екілік жапырақтардың төменгі жағында ерекше спорангиялар жиынтығы деп аталатын жинақта болады (2.37. суретінде, В, Г және Д). Әрбір спорангиялар жиынтығы индузий деп аталатын қорғаныш жабындысымен қапталған. Әрбір спорангий ішінде диплойдты аналық жасушалар споралары нәтижесінде гаплоидты споралар пайда болатын мейоттық бөліну жүреді. Индузийлер пісіп жетілгенде тыржиып және қопарылып түседі, ал спорангий қабырғалары ашылған кезде құрғап бастайды. Соңында, қабырғасы жайылады және споралар спорангийден катапульта сияөты атылып шығады (2.37, Д сурет).
Споралар гаметофитті ұрпақта туындата отырып, өседі. Гемотофит диаметрі, шамамен, 1 см болатын кішігірім жүректәрізді жасуша болып табылады (2.38 сурет). Бұл пластинка жасыл түсті, фотосинтезге қабілетті және жерге біржасушалы ризоидтармен бекінеді. Сондықтан мұндай нәзік өскінде кутикулалар жоқ, ол тез құрғайды және демек, ол тек ылғалды ортада тіршілік ете алады.
Гаметофиттің (өскін) төменгі жақтарында қарапайым архегонийлар және антеридийлар бар. Бұл репродуктивті ағзалар оның құрамындағы гаметаларды қорғайды. Гаметалар аналық жыныс жасушасындағы митоздың нәтижесінде пайда болады; бұл ретте, антеридийларда мүктәрізділер сияқты спермиялар пайда болады, ал әрбір архегонияда – бір жұмыртқажасушасынан болады. Спермиялар жалған аяқтармен жабдықталған. Ылғалды жағайларда піске спермийлар антеридийдан босап, су қабыршағында архегонияға қарай жүзеді. Ұрықтану нәтижесінде, диплоидты зигота түзіледі. Қырыққабаттәрізділер де мүктәрізділер сияқты ұрықтанулары суға тәуелді екеніне назар салыңыздар.
Зиготалар спорофитті ұқпақты туындатады. Жас ұқпақ өнім заттары функциясын өзінің жапырағы мен тамырлары орындағанша, ол гаметофиттн ала тұрады (2.38, Б сурет). Гаметофит көп кешіпей солып, құриды.
2.7.3. Тұқымдық өсімдіктер
Өсімдіктің ең гүлденуші тобы ұрық түзеді. Бұл өсімдіктер, тегі, қырыққабаттылардың және олардың туғандарының құрып біткен тұқым өкілдерінен түзілген. Тұқымдық өсімдіктердің систематикасы мен негізгі белгілері 2.10 кестесінде жинақталған.
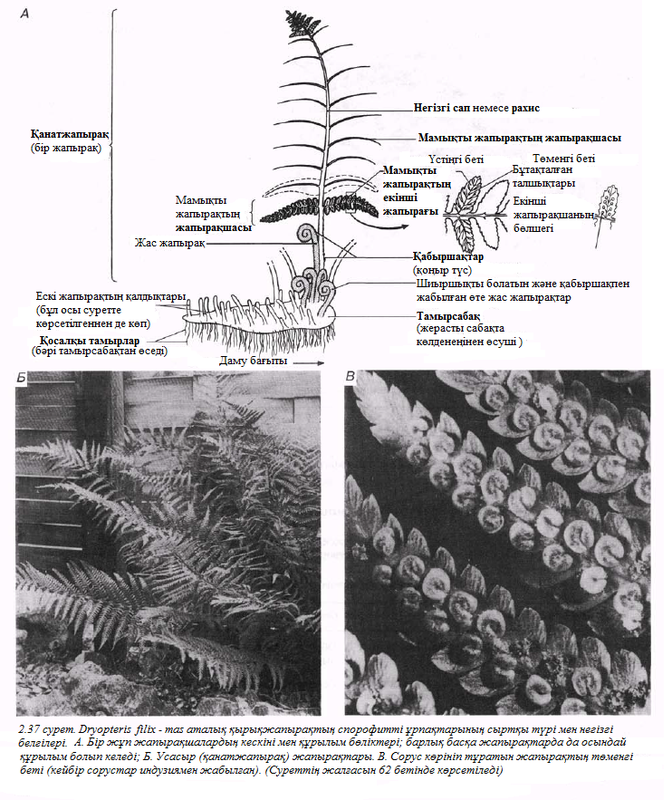
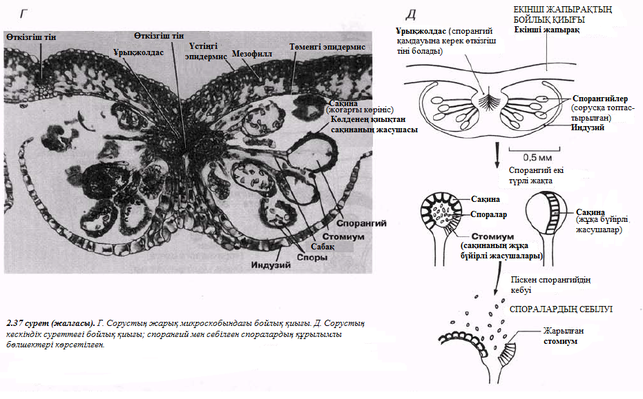
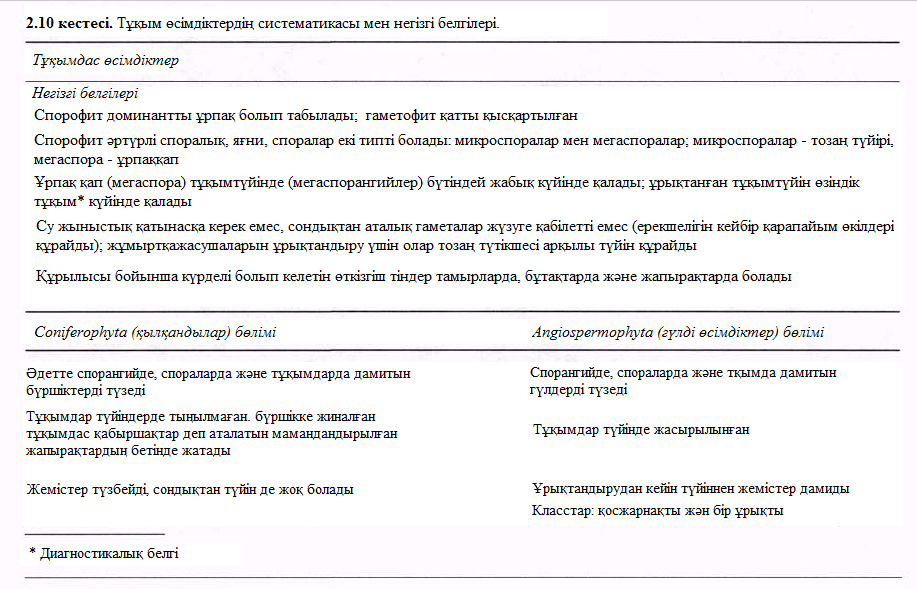
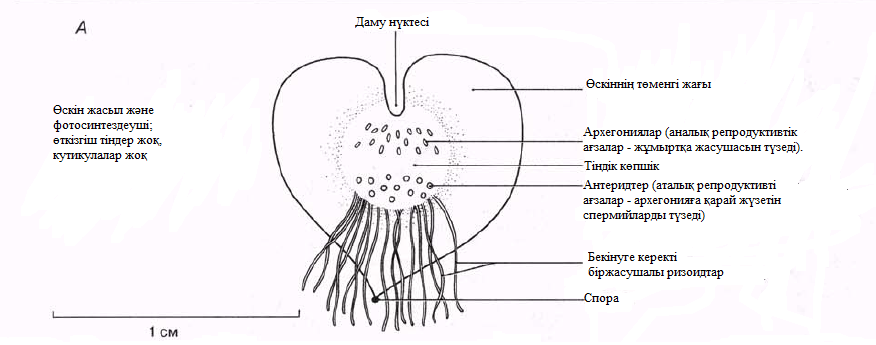
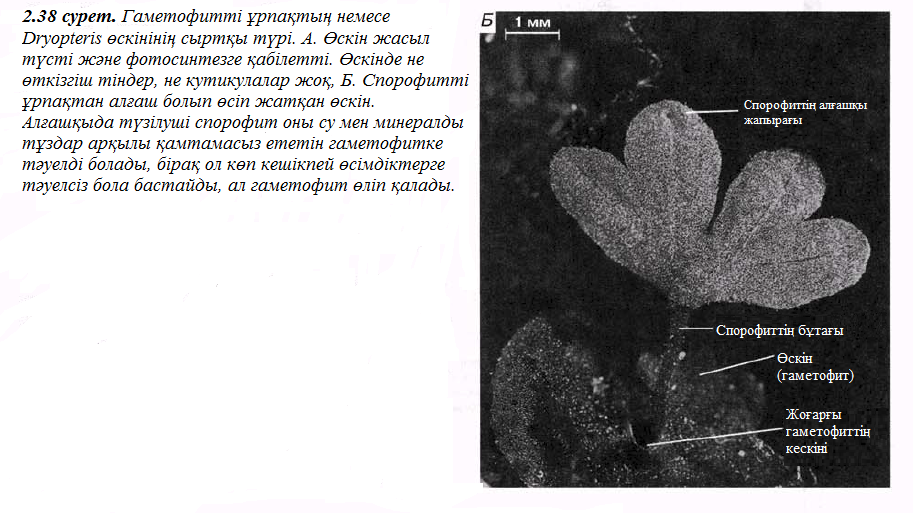
2.7.4. Coniferophyta (қылқандылар) бөлімі
Coniferophyta – ның негізгі белгілері 2.10 кестесінде жинақталған.
Қылқандылар (ашықтұқымдылар) – барлық жер шарына таралған дамушы өсімдіктер; олардың үлесіне, шамамен, планетаның барлық орманының 1/3іне келеді. Бұлар ағаштар және бұталар, негізінде инетәрізді жапырақты мәңгіжасылдар. Түрдің басым көпшілігі үлкен ендіктерде тіршілік етеді және солтүстік ар жақтағы барлық басқа да ағаштарда таралған. Қылқандылар тек қана ағаш пен кесілген ағаш материалдарына қолданылып қана қоймайтын, сонымен қоса, скипидар мен ағаш сүрек қабатына да қолданылатын, және «жұмсақ сүрек» ретінде шаруашылықта маңызы өте зор болып табылады. Қылқандыларға қарағайлар, балқарағайлар (қыста жапырақпен қазылатын), майқарағайлар, шыршалар және самырсындар жатады. Қылқандылардың әдеттегі өкілі – қарапайым қарағай (Pinus sylvestris).
Coniferophyta – ның негізгі белгілері 2.10 кестесінде жинақталған.
Қылқандылар (ашықтұқымдылар) – барлық жер шарына таралған дамушы өсімдіктер; олардың үлесіне, шамамен, планетаның барлық орманының 1/3іне келеді. Бұлар ағаштар және бұталар, негізінде инетәрізді жапырақты мәңгіжасылдар. Түрдің басым көпшілігі үлкен ендіктерде тіршілік етеді және солтүстік ар жақтағы барлық басқа да ағаштарда таралған. Қылқандылар тек қана ағаш пен кесілген ағаш материалдарына қолданылып қана қоймайтын, сонымен қоса, скипидар мен ағаш сүрек қабатына да қолданылатын, және «жұмсақ сүрек» ретінде шаруашылықта маңызы өте зор болып табылады. Қылқандыларға қарағайлар, балқарағайлар (қыста жапырақпен қазылатын), майқарағайлар, шыршалар және самырсындар жатады. Қылқандылардың әдеттегі өкілі – қарапайым қарағай (Pinus sylvestris).
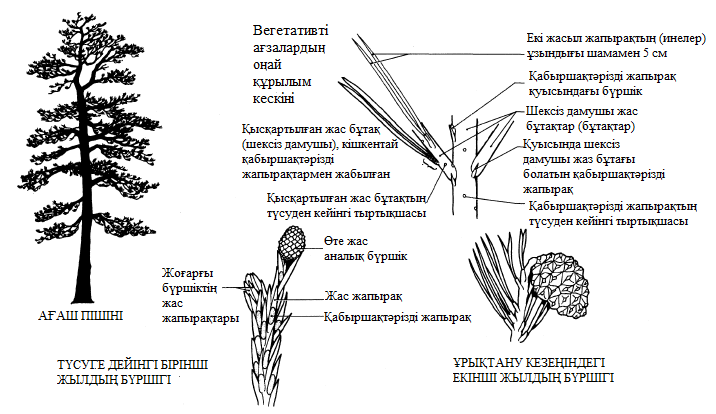

Бұл түр орталық және солтүстік Еуропада, бұрынғы КСРО және солтүстік Америкада таралған. Ол тағы Ұлыбританияда да жерсіндірілген болатын, бірақ табиғат жағдайларында тек қана Шотландияда өседі. Бұл биіктігі 36 м – ге дйін жететін әдемі ұлы ағаш қызғылт- немесе сары – қоңыр түсті қабықты қорғайтынды өсіреді және сәндік иақсаттарға, және ағаш немесе кесілген ағаш материалдарына қолданылады. Қарағайлар көбінесе құмды және жарлы таулы жерлерде өседі, сондықтан да тамырлы жүйесі оларда әдетте жердің жоғарғы бетінде жайылады және өте қатты бұтақтанады. Қарағайдың сыртқы бейнесі 2.39 суретінде көрсетілген.
Әр жыл сайын бүйір бүршек жақтағы күлтебастан діңгектің үстіңгі жағында жаңа күлтебас бұтағы өседі. Қарағайға және қылқанжапырақтыларға тән үшкірлік, қысқаларының (жастарының) бұтақтары үстінен астына дейін ұзындарымен ауыстырылу айналымынан анықталған. Есейе келе төменгі бұтақтар құрып, жоқ болып кетеді; сондықтан үлкен ағаштардың жоғарғы бөлігі әдетте бас жағында сақталады (2.39 сурет).
Негізгі бұтақтар мен өзек жоғарғы бүршік дамуының нәтижесінде жылдан жылға өсуін жалғастырады. Сондықтан, қылқандылар үшін шексіз даму тән деп айтады. Қабыршақты жапырақтар шиыршықты орналасқан; мұндай жапырақ қуыстарында ықшамдалған жас бұтақтар деп аталатын бойынан кіші бұтақтар өсетін (ұзындығы 2 – 3мм) бүршік бар. Бұл – жоғарысында 2 жапырақтан өсетін шектеулі бұтақтар болып табылады. Жас бұтақ өскен бойда қабыршақты жапырақ базасында жоқ болып және оның орнына тыртық қалады. Жапырақтар инеге ұқсас, оның жоғары жағының ауданын кішірейтеді, ал демек, бұл судың жоғалуы. Одан басқа, жапырақ ірі ақшыл сірқабықпен жабылған, ал саңылау жапырақ тініне тереңірек батырылған – бұл суға деген тағы бір бейімдеушілік болып табылады. Бұл ксероморфты сызықтар мәңгіжасыл жапырақтардан суық кезеңдерде, су мұз боп қатқанда, немесе оны жерден алып шығу қиынға соққан кезде бұлар судың ең төмен дейгейіне дейін жоғалып кетуіне апарады. 2 – 3 жылдан кейін қысқарған жас бұтақтар жапырақтармен бірге құлап түсіп, олардың орнына тағы 1 тыртық қалады.
Ағаштың өзі спорофитті ұрпақ болып табылады. Көктемде бір ағашта аталық және аналық бүршіктер пайда болады. Аталық бүршіктердің диаметрі 0,5 см айналасын құрайды; олар дөңгелек және базасында бүршік жоғарысындағы жас бұтақтар тобымен орналасады. Олар қуыста қабыршақтәрізді жапырақтармен ықшамдалған жас бұтақтармен бірге дамиды. Аналық бүршіктер ұуыста қабыршақтәрізді жапырақтармен жаңа мықты жас бұтақтардың соңында аталық бүршіктерден белгілі бір қашықтықта пайда болады және жинақсыз болып орналасады. Сондықтан бүршіктердің толық дамуы 3 жылды құрайды, бүршіктер өлшемдері бойынша өте қатты ажыратылады, және бір ағаштан 0,5 тен 6 см шамасындағы бүршіктерді байқауға болады. Жас бүршіктер жасыл түсті болып келеді, бірақ екінші жылы олар қоңыр немесе қызыл – қоңыр болып ауысады. Аталық және аналық бүршіктер орталық ось айналасында шиыршық орналасқан бір – бірімен тығыз қысылған спорофилдерден (модификацияланған жапырақтар) тұрады.
Әрбір аталық бүршіктің спорофилінің төменгісінің жоғарғы жағында 2 микроспорангия немесе тозың қап болады. Әрбір тозаң қап ішінде гаплоидты астық тозаңын немесе микроспордың пайда болуына әсер ететін мейоттық бөлініс жүреді. Олардың аталық гаметалар болады. Астық тозаңында олардың жаңбырмен таралуын жеңілдететінін 2 үлкен ауа қаптары болады. Мамырда бүршіктер барлық ауада оларда ұшатын тозаңдардың кесірінен сары болып қалады. Жаз соңында олар солып, құлап түседі.
Аналық бүршіктің әрбір спорофиллі төменгі жабылатын қабыршақтан және тұқымтүйінннен тұратын ірілеу жоғарғы қабыршақтардан құралады. Ірі қабыршақтардың жоғарғы бөлігінде іргелері арқылы тиіп тұрған ішінде аналық гаметалары түзілетін 2 тұқымтүйін бар. Бүршіктерді тозаңдандыру олардың дамуының ең бірінші жылында болады, бірақ ұрықтандыру тозаң түтіктері өскенше келесі көктемге дейін тоқтатылады. Ұрықтанған тұқымтүйінділерінен қанатты тұқымдар пайда болады. Тұқымның өсіп жетілуі 2 жылға дейін созылады, ал 3 – ші жылы олар тек қана төгіледі. Бұл уақытқа дейін бүршік ірі болып, қуарып қалады, ал қабыршақтар тұқымды жаңбырмен бірге жоймас бұрын жалаңаштап, сыртқа қарай түреді.
2.7.5. Angispermophyta бөлімі (жабықтұқымылар немесе гілді өсімдіктер)
2.10 кестесінде жабықтұқымдылардың негізгі белгілері берілген.
Жабықтұқымдылар басқа өсімдіктерге қарағанда құрғақтықтағы тіршілікке бейімделген. Олар бор кезеңінде шамамен 135 млн. жыл бұрын пайда болған, олар жер шарындағы өсімдіктер арасында басымдылық етуші орынды алып, олар тез қылқандыларды қысып шығарды және ең әртұрлі мекендейтін жерлердің негізін салды. Жабықтұқымдылардан кейбіреулері тұщы суға қайта оралды, ал бірнешетүрлері – тұзды су тіршілігіне кетті.
Жабықтұқымдылардың басқа жабық тұқымдардан ең ерекше белгісі, айтылып кеткендей, бұл бүршіктер мен бірге гүлдердің пайда болуы боп табылады. бұл гүлердің болуы бұл өсімдіктерге жәндіктердің, кейде тіпті құстардың және ұшатын тышқандардың тозақтандыруға тартуға көмектеседі. Гүлдердің жарқығ бояулары, хош иіс, жеуге болатын тозаң және шәрбас - бұлардың бәрі жануар тозаңдануларына тарту ортасы. Кейбір жағдайларда тозаңдандыру және гүлдер эволюциялар параллельді жүрген болатын, ол көп үлкенмамандандырылуға және өзара тиімді байланыстардың пайда болуына әкелді. Гүлдер бейімдеушілігі, қағида бойынша, тозаңдардың жәндіктерге айналуына үлкен дәрежедегі мүмкіндіктерге бағытталған болатын, сондықтан бұл үрдіс жаңбырмен тозаңдандыруға қарағанда үміті зор болып табылады. Жәндіктермен тозаңданатын өсімдіктерге жаңбырмен тозаңдандыруға керегі секілді көп мөлшердегі тозаңдар керек емес. Дегенмен, гүлді өсімдіктер жаңбыр тозаңына бейімделіп қалды.
Қос жарнақты және бір ұрықтылар
Жабықтұқымды өсімдіктер класс белгісін беруге болатын 2 үлкен топқа бөлінеді. Көбінесе бұлар бірұрықтылар және қосжарнақтылар деп 2-ге бөлінеді.
Әр жыл сайын бүйір бүршек жақтағы күлтебастан діңгектің үстіңгі жағында жаңа күлтебас бұтағы өседі. Қарағайға және қылқанжапырақтыларға тән үшкірлік, қысқаларының (жастарының) бұтақтары үстінен астына дейін ұзындарымен ауыстырылу айналымынан анықталған. Есейе келе төменгі бұтақтар құрып, жоқ болып кетеді; сондықтан үлкен ағаштардың жоғарғы бөлігі әдетте бас жағында сақталады (2.39 сурет).
Негізгі бұтақтар мен өзек жоғарғы бүршік дамуының нәтижесінде жылдан жылға өсуін жалғастырады. Сондықтан, қылқандылар үшін шексіз даму тән деп айтады. Қабыршақты жапырақтар шиыршықты орналасқан; мұндай жапырақ қуыстарында ықшамдалған жас бұтақтар деп аталатын бойынан кіші бұтақтар өсетін (ұзындығы 2 – 3мм) бүршік бар. Бұл – жоғарысында 2 жапырақтан өсетін шектеулі бұтақтар болып табылады. Жас бұтақ өскен бойда қабыршақты жапырақ базасында жоқ болып және оның орнына тыртық қалады. Жапырақтар инеге ұқсас, оның жоғары жағының ауданын кішірейтеді, ал демек, бұл судың жоғалуы. Одан басқа, жапырақ ірі ақшыл сірқабықпен жабылған, ал саңылау жапырақ тініне тереңірек батырылған – бұл суға деген тағы бір бейімдеушілік болып табылады. Бұл ксероморфты сызықтар мәңгіжасыл жапырақтардан суық кезеңдерде, су мұз боп қатқанда, немесе оны жерден алып шығу қиынға соққан кезде бұлар судың ең төмен дейгейіне дейін жоғалып кетуіне апарады. 2 – 3 жылдан кейін қысқарған жас бұтақтар жапырақтармен бірге құлап түсіп, олардың орнына тағы 1 тыртық қалады.
Ағаштың өзі спорофитті ұрпақ болып табылады. Көктемде бір ағашта аталық және аналық бүршіктер пайда болады. Аталық бүршіктердің диаметрі 0,5 см айналасын құрайды; олар дөңгелек және базасында бүршік жоғарысындағы жас бұтақтар тобымен орналасады. Олар қуыста қабыршақтәрізді жапырақтармен ықшамдалған жас бұтақтармен бірге дамиды. Аналық бүршіктер ұуыста қабыршақтәрізді жапырақтармен жаңа мықты жас бұтақтардың соңында аталық бүршіктерден белгілі бір қашықтықта пайда болады және жинақсыз болып орналасады. Сондықтан бүршіктердің толық дамуы 3 жылды құрайды, бүршіктер өлшемдері бойынша өте қатты ажыратылады, және бір ағаштан 0,5 тен 6 см шамасындағы бүршіктерді байқауға болады. Жас бүршіктер жасыл түсті болып келеді, бірақ екінші жылы олар қоңыр немесе қызыл – қоңыр болып ауысады. Аталық және аналық бүршіктер орталық ось айналасында шиыршық орналасқан бір – бірімен тығыз қысылған спорофилдерден (модификацияланған жапырақтар) тұрады.
Әрбір аталық бүршіктің спорофилінің төменгісінің жоғарғы жағында 2 микроспорангия немесе тозың қап болады. Әрбір тозаң қап ішінде гаплоидты астық тозаңын немесе микроспордың пайда болуына әсер ететін мейоттық бөлініс жүреді. Олардың аталық гаметалар болады. Астық тозаңында олардың жаңбырмен таралуын жеңілдететінін 2 үлкен ауа қаптары болады. Мамырда бүршіктер барлық ауада оларда ұшатын тозаңдардың кесірінен сары болып қалады. Жаз соңында олар солып, құлап түседі.
Аналық бүршіктің әрбір спорофиллі төменгі жабылатын қабыршақтан және тұқымтүйінннен тұратын ірілеу жоғарғы қабыршақтардан құралады. Ірі қабыршақтардың жоғарғы бөлігінде іргелері арқылы тиіп тұрған ішінде аналық гаметалары түзілетін 2 тұқымтүйін бар. Бүршіктерді тозаңдандыру олардың дамуының ең бірінші жылында болады, бірақ ұрықтандыру тозаң түтіктері өскенше келесі көктемге дейін тоқтатылады. Ұрықтанған тұқымтүйінділерінен қанатты тұқымдар пайда болады. Тұқымның өсіп жетілуі 2 жылға дейін созылады, ал 3 – ші жылы олар тек қана төгіледі. Бұл уақытқа дейін бүршік ірі болып, қуарып қалады, ал қабыршақтар тұқымды жаңбырмен бірге жоймас бұрын жалаңаштап, сыртқа қарай түреді.
2.7.5. Angispermophyta бөлімі (жабықтұқымылар немесе гілді өсімдіктер)
2.10 кестесінде жабықтұқымдылардың негізгі белгілері берілген.
Жабықтұқымдылар басқа өсімдіктерге қарағанда құрғақтықтағы тіршілікке бейімделген. Олар бор кезеңінде шамамен 135 млн. жыл бұрын пайда болған, олар жер шарындағы өсімдіктер арасында басымдылық етуші орынды алып, олар тез қылқандыларды қысып шығарды және ең әртұрлі мекендейтін жерлердің негізін салды. Жабықтұқымдылардан кейбіреулері тұщы суға қайта оралды, ал бірнешетүрлері – тұзды су тіршілігіне кетті.
Жабықтұқымдылардың басқа жабық тұқымдардан ең ерекше белгісі, айтылып кеткендей, бұл бүршіктер мен бірге гүлдердің пайда болуы боп табылады. бұл гүлердің болуы бұл өсімдіктерге жәндіктердің, кейде тіпті құстардың және ұшатын тышқандардың тозақтандыруға тартуға көмектеседі. Гүлдердің жарқығ бояулары, хош иіс, жеуге болатын тозаң және шәрбас - бұлардың бәрі жануар тозаңдануларына тарту ортасы. Кейбір жағдайларда тозаңдандыру және гүлдер эволюциялар параллельді жүрген болатын, ол көп үлкенмамандандырылуға және өзара тиімді байланыстардың пайда болуына әкелді. Гүлдер бейімдеушілігі, қағида бойынша, тозаңдардың жәндіктерге айналуына үлкен дәрежедегі мүмкіндіктерге бағытталған болатын, сондықтан бұл үрдіс жаңбырмен тозаңдандыруға қарағанда үміті зор болып табылады. Жәндіктермен тозаңданатын өсімдіктерге жаңбырмен тозаңдандыруға керегі секілді көп мөлшердегі тозаңдар керек емес. Дегенмен, гүлді өсімдіктер жаңбыр тозаңына бейімделіп қалды.
Қос жарнақты және бір ұрықтылар
Жабықтұқымды өсімдіктер класс белгісін беруге болатын 2 үлкен топқа бөлінеді. Көбінесе бұлар бірұрықтылар және қосжарнақтылар деп 2-ге бөлінеді.
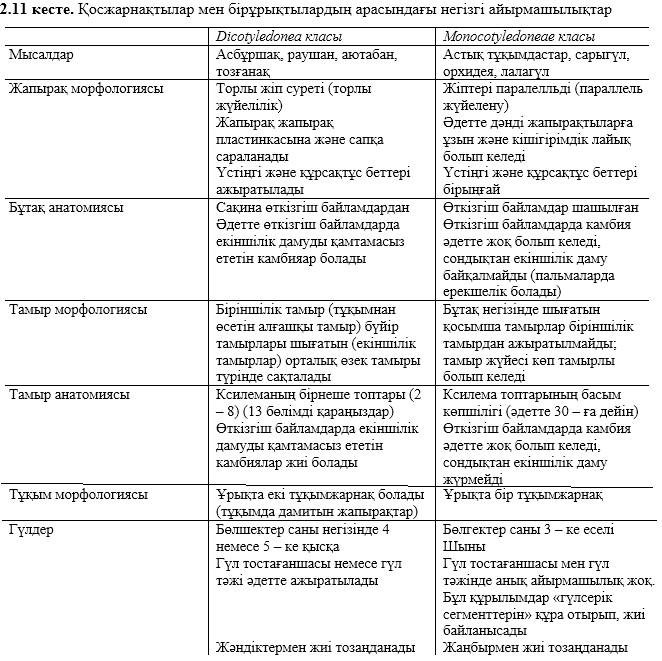
|
2.11 кестесінде олардың айырмашылығының негізгі белгілері берілген (2.40 суретін де қараңыздар). Қазіргі заманауи көрсеткіштер бойынша бірұрықтылар қосжарнақтыдан пайда болғаны келісімді.
Жабықтұқымдылар шөпті (яғни, қуарылмайтын) және ағашты болады. Соңғысына бұталар және ағаштар жатады. Бұл өсімдіктерде тіректі функциясын орындайтын, одан басқа өткізгіш тін болып қызмет ететін екілік ксилемалардың (сүрек) үлкен санын құрайды. Ксилема өткізгіш шоқ камбия белсенділігінен пайда болады. Бұл камбий бұтақтардағы және тамырлардағы ксилемалар мен флоэмалар арасында орналасқан жасуша қабықшасы болып саналады. Камбий жасушалары бөлініске қабілеттілікті сақтайды. Бұдан пайда болған ксилема екілік ксилема немесе сүрек деп аталады. Шөпті өсімдіктер немесе шөптер жасуша серпімділігіне және колленхима, склеренхима және ксилема сияқты механикалық тіндердің басым көпшілігіне ғана тірек болады делінеді; сондықтан олардың өздері де маңызды емес екені таңғаларлықтай емес. Шөпті өсімдіктерде не камбия мүлдем жоқ, немесе ода болса да ешқандай мәні жоқ. Көп шөпті өсімдіктер біржылдық, яғни тұқымнан тұқымға дейінгі барлық айналым 1 жылда біте салады. Кейбір шөпті өсімдіктердің бадана, түйнекбадана немесе түйнек сияқты түрлерінің ағзалары көпжылдық болады, олай болатыны өсімдіктің қыстап шығуының арқасында және ыңғайсыз жағдайлар кезеңдеріне шыдауында, мысалы құрғақшылықта (20 бөлім). Мұндай шөпті өсімдіктер екіжылдық немесе көпжылдық деп аталады. бірінші жағдайда олар тұқым түзеді, ал екінші жылы құриды, ал екіншісінде көп жыл тіршілік етеді. Бұтақтар мн ағаштар – көпжылдықты өсімдіктер. Олар мәңгіжасылдар немесе жапырақтылар болуы мүмкін. Мәңгіжасылды өсімдіктер жапырақты оны жыл бойы түзіп, лақтырып отырады, сондық өсімдікте әрдайым жапырақтар болады. Жапырақтылар жылдың суық немесе құрғақ мезгілдерінде жапырақтарды лақтырады. Жабықтұқымдылардың әртүрлілігі туралы мысал ретінде бұл өсімдік топының әдеттегі өкілдерін жинақтай отырып, 2.41 – 2.44 суреттерінде көрсетілген. |


|
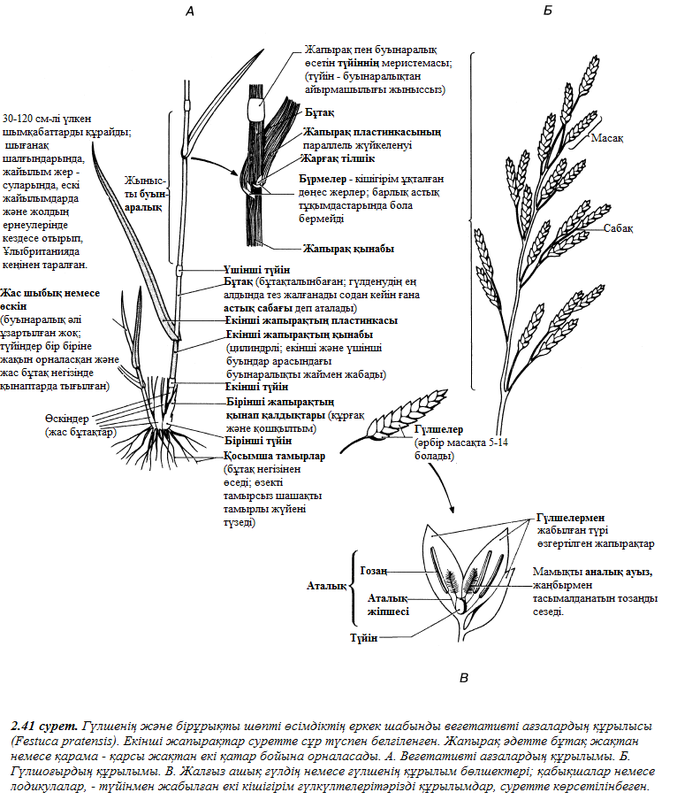
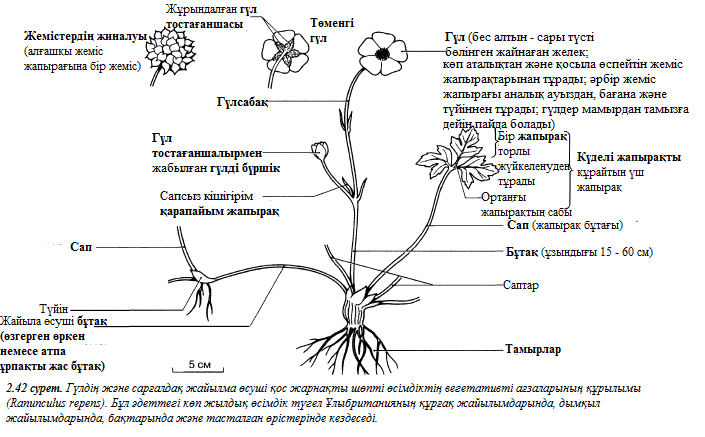
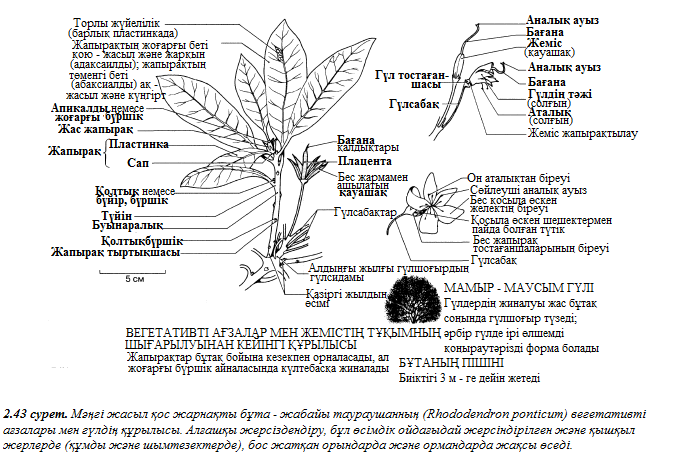
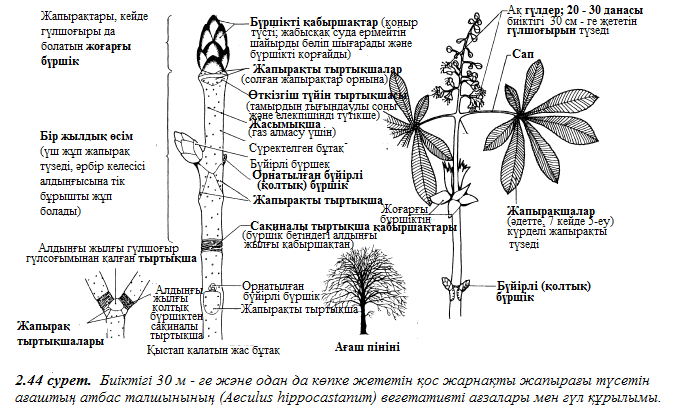
2.7.6. Өсімдіктердің құрғақтық өміріне бейімделгіштігі
Енді біз мүктәрізді, қырыққұлақтәрізді, ашықтұқымдастылар мен жабықтұқымдастылар (гүлділер) сияқты 4 топтың айырмашылық белгілерімен танысқаннан кейін бізге өсімдіктердің құрғақ өмірге бейімделгішін сипаттайтын эволюциялық үрдісді елестету оңайға соғады.
Мәселелер
Ауыр қиындықтардың бірі болып саналған сулы өмірден жерге ауысуды жеңіп алу үшін құрғату мәселесі орын алды. Сол немесе басқа жолмен қорғалған кез келген өсімдік, мысалы, ақсары кутикуламен жабылмаған, өте қатты құрғалады және шүбәсіз өледі. Тіпті бұл мәселені деңіп шықсақ та, басқа да шешілмеген мәселелер қалады. Және бәрінен де бұрын сұрақ былай: қалайша ұрықтандыруды ұтымды жандандырауға болады. Алғашқы өсімдіктерде ұрықтануға тек қана суда жүзіп жүріп қана аналық гаметаларға жақындайтын аталық гаметалардың қатысуымен болған.
Әдетте, құрлықта пайда болған алғашқы өсімдіктер эволюционды өкілдерінің озаты болған репродуктивті ағзалар пайда болды, атап айтқанда архегония (аналық) және антеридтер (аталық); бұл ағзаларда олар жасырын болды, ал демек гаметалар да қорғалған. Бұл жайт басқа да барлық нақты бейімдеушіліктің құрғақтықтан қашуға көмектесетін қатары болып табылады; кейбір жасыл балдырларға құрғақтықты иеленуіне рұқсат берді.
Өсімдіктердің ең маңызды эволюционды үрдістерінің бірі – бұл олардың жай – жайдан суға тәуелсіздігінің артуы.
Төменде судан жерге дейінгі тіршілікке ауысу байланыстарының негізгі қиындықтары берілген:
Мүктер жер жағдайларындағы споралардың таралуына қабілетті болып келеді: бұл қауашақтардың құрғақ болуынан және кіші жаңбыр спораларының себілуіне тәуелді болып келеді. Бірақ бұл өсімдіктер келесі себептер бойынша суға байланысты табылуда.
Қырықжапырақтылар
2.5. Қырықжапырақтылар бауырмүктілер мен мүктерге қарағанда жердегі өмірге жақсы бейімделген. Бұл нені білдіреді?
2.6. Мүктер, қырықжапырақтылар мен бауырмүктілер қандай маңызды себептермен жердегі өмірге нашар бейімделгіштікті көрсетеді?
Тұқымдас өсімдіктер – қылқандылар мен гүлділер
Жер өсімдіктерімен қақтығысатын негізгі мәселелер гаметофитті ұрпақтардың әлсіздігімен байланысты болады. Мысалы, гаметофит усасыры – бұл жұмаыртқа жасушасына жету үшін суды қажетсінетін аталық гаметаларды (спермийлар) түзетін жұмсақ өскін. Бірақ тұқымдас өсімдіктерде гаметофит қорғанышты және өте қатты қысқартылған.
Тұқымдас өсімдіктердің маңызды 3 пайдалы жақтары бар: біріншіден, әртүрлі споралылық; екіншіден, жүзе алмайтын аталық гаметалардың болуы және үшіншіден, тұқымның түзілуі.
ӘРТҮРЛІ СПОРАЛЫЛЫҚ ЖӘНЕ ЖҮЗЕ АЛМАЙТЫН АТАЛЫҚ ГАМЕТАЛАР. Өсімдіктер эволюциясында өте маңызды рөлді усасыр мен оның спораның 2 түрін түзетін жақын туыстарының пайда болуымен орын алды. Бұның пайда болуы әртүрлі споралылық, ал өсімдіктер – әртүрлі споралы. Барлық тұқымдас өсімдіктер - әртүрлі споралыға байланысты болады. Олар мегаспоралар деп аталатын спорангийде бір типті (мегаспорангий) болатын ірі спораларды түзеді және микроспоралар деп спорангийлерде екінші бір типті (микроспорангийлер) болатын кіші спораларды айтады.
Енді біз мүктәрізді, қырыққұлақтәрізді, ашықтұқымдастылар мен жабықтұқымдастылар (гүлділер) сияқты 4 топтың айырмашылық белгілерімен танысқаннан кейін бізге өсімдіктердің құрғақ өмірге бейімделгішін сипаттайтын эволюциялық үрдісді елестету оңайға соғады.
Мәселелер
Ауыр қиындықтардың бірі болып саналған сулы өмірден жерге ауысуды жеңіп алу үшін құрғату мәселесі орын алды. Сол немесе басқа жолмен қорғалған кез келген өсімдік, мысалы, ақсары кутикуламен жабылмаған, өте қатты құрғалады және шүбәсіз өледі. Тіпті бұл мәселені деңіп шықсақ та, басқа да шешілмеген мәселелер қалады. Және бәрінен де бұрын сұрақ былай: қалайша ұрықтандыруды ұтымды жандандырауға болады. Алғашқы өсімдіктерде ұрықтануға тек қана суда жүзіп жүріп қана аналық гаметаларға жақындайтын аталық гаметалардың қатысуымен болған.
Әдетте, құрлықта пайда болған алғашқы өсімдіктер эволюционды өкілдерінің озаты болған репродуктивті ағзалар пайда болды, атап айтқанда архегония (аналық) және антеридтер (аталық); бұл ағзаларда олар жасырын болды, ал демек гаметалар да қорғалған. Бұл жайт басқа да барлық нақты бейімдеушіліктің құрғақтықтан қашуға көмектесетін қатары болып табылады; кейбір жасыл балдырларға құрғақтықты иеленуіне рұқсат берді.
Өсімдіктердің ең маңызды эволюционды үрдістерінің бірі – бұл олардың жай – жайдан суға тәуелсіздігінің артуы.
Төменде судан жерге дейінгі тіршілікке ауысу байланыстарының негізгі қиындықтары берілген:
- Құрғақтық. Ауа – бұл кеуіп кетуге қабілетті орта, ал су барлық себептер қатары өміріне қажетті болып келеді (3.1.2 бөлімі). Демек, бұл судың алынуына және жиналуына бейімдеушілікті тудыруға қажет.
- Көбею. Жұмсақ жыныстық жасушалар қорғану қажет, ал қозғалмалы аталық гаметалар (спермийлар) аналық гаметалармен тек қана суда кездесу мүмкін.
- Тірек. Ауаның судан айырмашылығы өсіміктердің тірегі болып қызмет ете алмайды.
- Қоректену. Өсімдіктерге жарық пен фотосинтез үшін көміртек диоксиді (СО2) қажет, сондықтан өсімдіктің бір бөлігі болса да жердің бетіне шығып тұру қажет.
- Газ алмасу. Фотосинтез бен тыныс алу үшін көміртек диоксиді мен оттек алмасулары айналадағы ерітінді мен еес, қоршаған ортамен жүру керек.
- Қоршаған ортаның факторлары. Су көп болған кезде, айтқандай, мұхит пен көлде қоршаған орта жағдайларының үлкен бірқалыптылығын қамтамасыз етеді. Жердегі мекендену ортасы едәуір үлкен дәрежеде маңызды факторларды сипаттайды, олар температура, жарық қарқындылығы, иондар концентрация және рН.
Мүктер жер жағдайларындағы споралардың таралуына қабілетті болып келеді: бұл қауашақтардың құрғақ болуынан және кіші жаңбыр спораларының себілуіне тәуелді болып келеді. Бірақ бұл өсімдіктер келесі себептер бойынша суға байланысты табылуда.
- Су олардың көбеюі үшін керек, сондықтан спермийлар архегонияда жүзуі керек. Бұл өсімдіктерде оларға спермийларды тек қана ылғалды ортада босатуға рұқсат берілген бейімдеушіліктер пайда болды, себебі антеридтер осындай ортада ғана ашылады. Бұл өсімдіктер жаймен жердегі тіршілікке үйреніскен, сондықтан гаметалар оларда антерид пен архегония сияқты қорғанышты орындарда түзіледі.
- Оларда арнайы тірек тіндері жоқ және сондықтан өсімдіктің жоғарыға дамулары шектеулі.
- Мүктәрізділерде субстратқа алыстан еніп кетуге қабілеттілік жоқ және олар тек жер бетінде және оның ылғалды және минералды тұздар жоғарғы қабаттарында ғана тіршілік ете алады. Бірақ оларда су түбіне бекінетін ризоидтар бар; бұл – қатты субстрат тіршілігіне бейімдеушіліктің бірі болып табылады.
Қырықжапырақтылар
2.5. Қырықжапырақтылар бауырмүктілер мен мүктерге қарағанда жердегі өмірге жақсы бейімделген. Бұл нені білдіреді?
2.6. Мүктер, қырықжапырақтылар мен бауырмүктілер қандай маңызды себептермен жердегі өмірге нашар бейімделгіштікті көрсетеді?
Тұқымдас өсімдіктер – қылқандылар мен гүлділер
Жер өсімдіктерімен қақтығысатын негізгі мәселелер гаметофитті ұрпақтардың әлсіздігімен байланысты болады. Мысалы, гаметофит усасыры – бұл жұмаыртқа жасушасына жету үшін суды қажетсінетін аталық гаметаларды (спермийлар) түзетін жұмсақ өскін. Бірақ тұқымдас өсімдіктерде гаметофит қорғанышты және өте қатты қысқартылған.
Тұқымдас өсімдіктердің маңызды 3 пайдалы жақтары бар: біріншіден, әртүрлі споралылық; екіншіден, жүзе алмайтын аталық гаметалардың болуы және үшіншіден, тұқымның түзілуі.
ӘРТҮРЛІ СПОРАЛЫЛЫҚ ЖӘНЕ ЖҮЗЕ АЛМАЙТЫН АТАЛЫҚ ГАМЕТАЛАР. Өсімдіктер эволюциясында өте маңызды рөлді усасыр мен оның спораның 2 түрін түзетін жақын туыстарының пайда болуымен орын алды. Бұның пайда болуы әртүрлі споралылық, ал өсімдіктер – әртүрлі споралы. Барлық тұқымдас өсімдіктер - әртүрлі споралыға байланысты болады. Олар мегаспоралар деп аталатын спорангийде бір типті (мегаспорангий) болатын ірі спораларды түзеді және микроспоралар деп спорангийлерде екінші бір типті (микроспорангийлер) болатын кіші спораларды айтады.
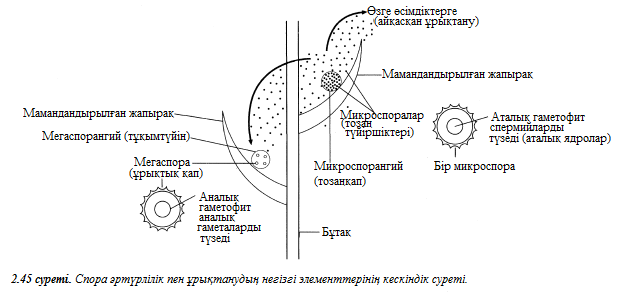
Олар өсе келе гаметофиттерді түзеді (2.34 сурет).мегаспоралар аналық гаметофиттерде, ал микроспоралар аталық гаметофиттерде болады. Тұқымдас өсімдіктерде мегаспора мен микроспораны түзетін гаметофиттер өлшемдері бойынша өте аз және спорадан ешқашан босап шықпайды. Гаметофиттер құрғақтықтан қорғанышты болып саналады, осылайша олардың өзіндік маңызды эволюциондық жетістігі бар. Бірақ, аталық гаметофиттің спермийлары микроспоралардың себілуін жеңілдету үшін аналық гаметофитке ауысуы керек болады. Олар ұсақ бола тұра, көп санды болып түзіле алады және ата – аналар спорофитінен алыста жүріп ақ жаңбырмен таралып кете алады. Кенеттен олар тұқымдас өсімдіктерде ата – аналық спорофиттерден бөлінбейтін мегаспоралармен тығыз жақындықта болуы мүмкін (2.45 сурет). Дәл осындай жолмен өсімдіктерде өздерін микроспоралармен көрсететін тозаң түйіршіктері арқылы болатын тозаңдандыру жүреді. Тозаң түйіршіктерінде аталық гаметалар түзіледі.
Тұқымдас өсімдіктерде тағы бір эволюциялықпайда бой көрсетті. Аталық гаметаларға бұдан әрі аналық гаметаларға қарай жүзі қажеті жоқ, соныдқтан тұқымдас өсімдіктерде тозаң түтікшелері пайда болды. Олар тозаң тостағандарынан дамиды және аналық гаметалар бағытында өседі. Бұл түиік арқылы аталық гаметалар аналық гаметаларға жетеді және оны ұрықтатады. Жүзгіш спермийлар бұдан әрі түзілмейді, ұрықтануға тек қана аталық ядролар қатысады.
Демек, өсімдіктерде суға тәуелсіз бір ұрықтану іске асты. Тұқымдас өсімдіктер жер дамуында басқа өсімдіктерден кәдімгідей асып түсті, бұл дегеніміз дәл осы себеп тағы да іске асты. Алғашқы тозаңдандыру тек қана жаңбыр көмегімен жүрді, бұл үрдіс айтарлықтай кенеттен болды, бұл көптеген тозаңдардың жоғалуымен сипатталады. Бірақ эволюцияның ерте кезеңдерінде шамамен 300 млн жыл бұрын тас көмір дәуірінде ұшатын жәндіктер пайда бола бастады, ал олардың көмегімен тозаңдануды тиімдірек қолдануға мүмкіндңк туды. Гүлді өсімдіктерде тозаңданудың жәндіктермен қолдануы кеңінен таралды, ал қылқандыларда әлі де болса тозаңдану жаңбырмен жүреді.
ТҰҚЫМ. Ертедегі мегаспораның әртүрлі споралы өсімдіктері микроспораға тән ата – аналық спорофиттен босап шығатын болған. Тұқымдас өсімдіктерде мегаспоралар мегаспорангийде немесе тұқымтүйінде қала отырып, ата – аналық өсімдіктен бөлінбейді (2.45 суреті). Тұқымтүйінде аналық гамета болады. Аналық гаметаның ұрықтануынан кейін тұқымтүйін тұқым деп аталып кетеді. Осылайша, тұқым дегеніміз – тұқымтүйіннің ұрықтануы болып табылады. Тұқымтүйін мен тұқымның бар болуы тұқым өсімдігіне белгілі бір пайда әкеледі.
2.7.7. Құрғақ тіршілікке тұқымдас өсімдіктер бейімделгішінің қысқаша санап шығылуы
Тұқымдас өсімдіктердің негізгі пайдалы жақтары келесіде беріледі.
2.8.1. Эволюционды үрдістер
Жануарлар эакариот патшалығының төрттен бір бөлігін құрайды (2.4 суреті). Олардың бәрі көпжасушашылар, сондықтан жануарларға тән біржасушалы ағзалар Protoctista патшалығына тән болып келеді. Жануарлар өсімдіктерден автотрофты емес гетеротрофты болғанымен ерекшеленеді. Саңырауқұлақтардан және көпжасушалы гетеротрофты эукариоттардан жануарлар өнімді тауып алу қабілеттігімен ерекшеленеді. Саңыракқұлақтарды өнімді сіңіруші ағзалар деп сипаттауға болады, ал жануарларды жұтып алушы ағзалар деуге болады. Саңырауқұлақтар өнімді ағза сыртында қорытады және сосын барып қорытылудан кейінгі қоректі сіңіреді, ал жануарларда қорек әдетте жұтып алған дене ішінде соңғы қорытылған қорек болып енгізіледі (яғни, қоректер қабылдануы ағза ішінде). Барлық ұорытылмаған астар бөлінеді (денеден сыртұа шығарылады). Жануарлар эволюциясының үрдісінде қорек тәртібінің әртүрлі стратегиясының қатарлары іске асты, атап айтқанда жыртқыштық, өсімдік қоректенетіндік, талғаусыз қоректілік және паразитизм. Егер саңырауқұлақтар сөзінің дәлме дәл мағынасында өзінің қорегінде өсетін болса, онда жануарларға оларды жиі іздеуге жағдай туыындамайды. Бұл жағдайда олар бір жерден екінші басқа жерге көші үшін локомоцияға қабілеттілікті иелену керек, ал бұл өз кезегінде жүйке жүйесінің, сезу мен қозғалтқыш жүйке шеті ағзаларының жоғары дамуының бар болуын талап етеді. Одан басқа, ірі жануарлардың локомоциясы бұлшықеттер мен қаңқаның және тіректі функцияны орындайтынның бар болуын талап етеді.
Жануарлар оқуында барған сайын қиындай беретін күрделі дәрежедегі ұжымның дене құрылысын қамтамасыз ететін эволюция үрдістерін зерттеу керек. Тек қана бір жануар тобында ғана, яғни жөкеде нағыз тіндер түзіліп көрген жоқ (2.12 кестесі), бірақ басқа да барлық жануарларда тіндер түзіледі.
Тұқымдас өсімдіктерде тағы бір эволюциялықпайда бой көрсетті. Аталық гаметаларға бұдан әрі аналық гаметаларға қарай жүзі қажеті жоқ, соныдқтан тұқымдас өсімдіктерде тозаң түтікшелері пайда болды. Олар тозаң тостағандарынан дамиды және аналық гаметалар бағытында өседі. Бұл түиік арқылы аталық гаметалар аналық гаметаларға жетеді және оны ұрықтатады. Жүзгіш спермийлар бұдан әрі түзілмейді, ұрықтануға тек қана аталық ядролар қатысады.
Демек, өсімдіктерде суға тәуелсіз бір ұрықтану іске асты. Тұқымдас өсімдіктер жер дамуында басқа өсімдіктерден кәдімгідей асып түсті, бұл дегеніміз дәл осы себеп тағы да іске асты. Алғашқы тозаңдандыру тек қана жаңбыр көмегімен жүрді, бұл үрдіс айтарлықтай кенеттен болды, бұл көптеген тозаңдардың жоғалуымен сипатталады. Бірақ эволюцияның ерте кезеңдерінде шамамен 300 млн жыл бұрын тас көмір дәуірінде ұшатын жәндіктер пайда бола бастады, ал олардың көмегімен тозаңдануды тиімдірек қолдануға мүмкіндңк туды. Гүлді өсімдіктерде тозаңданудың жәндіктермен қолдануы кеңінен таралды, ал қылқандыларда әлі де болса тозаңдану жаңбырмен жүреді.
ТҰҚЫМ. Ертедегі мегаспораның әртүрлі споралы өсімдіктері микроспораға тән ата – аналық спорофиттен босап шығатын болған. Тұқымдас өсімдіктерде мегаспоралар мегаспорангийде немесе тұқымтүйінде қала отырып, ата – аналық өсімдіктен бөлінбейді (2.45 суреті). Тұқымтүйінде аналық гамета болады. Аналық гаметаның ұрықтануынан кейін тұқымтүйін тұқым деп аталып кетеді. Осылайша, тұқым дегеніміз – тұқымтүйіннің ұрықтануы болып табылады. Тұқымтүйін мен тұқымның бар болуы тұқым өсімдігіне белгілі бір пайда әкеледі.
- Аналық гаметофит тұқымтүйінмен қорғалған. Ол толығымен ата – аналық спорофитке тәуелді болады және еркін тіршілік етуші гаметофиттен айырмашылығы құрғатуды сезе алмайды.
- Тұқымға ұрықтанудан кейін бұрынғыша бөлінбеген ата – аналық өсімдіктен алынған гаметофиттен қоректік заттардың қоры құралады. Бұл қор тұқымның өсіп – өнуінен кейіннен дамушы зиготаға (келесі спорофит ұрпағы) қолданылады.
- Тұқым қолайсыз жағдайларды бастан кешіру үшін белгіленген және өсіп – өну үшін қолайлы жағдайларға дейін тыныш күйінде қалады.
- Тұқымда оның таралуын жеңілдететін әртүрлі қабілеттер дамуы мүмкін.
2.7.7. Құрғақ тіршілікке тұқымдас өсімдіктер бейімделгішінің қысқаша санап шығылуы
Тұқымдас өсімдіктердің негізгі пайдалы жақтары келесіде беріледі.
- Гаметофитті ұрпақ өте қатты керісінше дамиды және бұл толығымен әрқашан гаметофиті ішінде қорғалып тұратын спорофиттің құрғақтықтағы тіршілігінің қабілеттілігіне тәуелді болады. Басқа өсімдіктерде гаметофит өте оңай кебеді.
- Ұрықтану суға тәуелсіз жүреді. Аталық гаметалар қозғалмайтындар болып келеді және жаңбыр немесе жәндіктердің тозаң түйіршіктерінің ішінде орнығады.
- Ұрықтанған тұқымтүйіндер (тұқым) таралмай тұрып, өнім мен қорғаныш затын алатын ата – аналық спорофитте белгілі бір уақытқа қалады.
- Көптеген тұқымдас өсімдіктерде тіректі функцияны . алып жүретін көп санды сүректердің бөлінуімен екінші дамуымен байқалады. Мұндай өсімдіктер тартыста жарықпен өзге де қажеттіліктер үшін тиімді бәсекелесе алатын ағаштар мен бұталарда өседі.
- Нағыз тамырлар жерден ылғалды шығаруды қамтамасыз етеді.
- Өсімдіктер құрғақтықтан сақтанатын су іткізбейтін кутикуласы (немесе екінші дамудан кейін пайда болатын тығын) бар эпидермиспен қапталған.
- Эпидермис жер өсімдіктерінің бөлшегі, әсіресе жапырақтың, ол өсімдікпен атмосфера арасындағы газ алмасуды іске асыратын саңылау деп аталатын көптеген кішігірім тесіктермен тізбектелген.
- Өсімдіктерде тағы мамандырылған ыстық құрғақ жағдайдағы тіршілікке бейімделгіштер бар (19 және 20 бөлім).
2.8.1. Эволюционды үрдістер
Жануарлар эакариот патшалығының төрттен бір бөлігін құрайды (2.4 суреті). Олардың бәрі көпжасушашылар, сондықтан жануарларға тән біржасушалы ағзалар Protoctista патшалығына тән болып келеді. Жануарлар өсімдіктерден автотрофты емес гетеротрофты болғанымен ерекшеленеді. Саңырауқұлақтардан және көпжасушалы гетеротрофты эукариоттардан жануарлар өнімді тауып алу қабілеттігімен ерекшеленеді. Саңыракқұлақтарды өнімді сіңіруші ағзалар деп сипаттауға болады, ал жануарларды жұтып алушы ағзалар деуге болады. Саңырауқұлақтар өнімді ағза сыртында қорытады және сосын барып қорытылудан кейінгі қоректі сіңіреді, ал жануарларда қорек әдетте жұтып алған дене ішінде соңғы қорытылған қорек болып енгізіледі (яғни, қоректер қабылдануы ағза ішінде). Барлық ұорытылмаған астар бөлінеді (денеден сыртұа шығарылады). Жануарлар эволюциясының үрдісінде қорек тәртібінің әртүрлі стратегиясының қатарлары іске асты, атап айтқанда жыртқыштық, өсімдік қоректенетіндік, талғаусыз қоректілік және паразитизм. Егер саңырауқұлақтар сөзінің дәлме дәл мағынасында өзінің қорегінде өсетін болса, онда жануарларға оларды жиі іздеуге жағдай туыындамайды. Бұл жағдайда олар бір жерден екінші басқа жерге көші үшін локомоцияға қабілеттілікті иелену керек, ал бұл өз кезегінде жүйке жүйесінің, сезу мен қозғалтқыш жүйке шеті ағзаларының жоғары дамуының бар болуын талап етеді. Одан басқа, ірі жануарлардың локомоциясы бұлшықеттер мен қаңқаның және тіректі функцияны орындайтынның бар болуын талап етеді.
Жануарлар оқуында барған сайын қиындай беретін күрделі дәрежедегі ұжымның дене құрылысын қамтамасыз ететін эволюция үрдістерін зерттеу керек. Тек қана бір жануар тобында ғана, яғни жөкеде нағыз тіндер түзіліп көрген жоқ (2.12 кестесі), бірақ басқа да барлық жануарларда тіндер түзіледі.


Жасуша тобында тін деп құрылымы мен түзілуімен және белгілі бір орындаумен арнайы функция арқылы жасалатын жұмыстары ұқсас болып келетіндерді айтады. Бұдан өзінің ерекше функциясын орындайтын әртүрлі тіндерді байқауға болады. Өзінің ерекше функциясының тінінің жетістігі саралану деп аталады. Бұл қағида әртүрлі функцияны орындау үшін әртүрлі жасушалы органеллалар мамандандырылған субжасушалы деңгейде де іске асады.
Ең бөлінісі, қағида бойынша, жұмыстың тиімділігін көтереді. Ұжым жеігейі тінге қарағанда әл де болса да биігірек болып саналады. тінге қатысты бірнеше жұмыскерлер ағза құрайды, мысалы, қарын. Ағзаларға қатысты жұмыскерлер тобы жүйе құрайды, сонымен қоса, ас қорыту сияқты. Әртүрлі барлық ағза жүйесінің байланастырылуы ағзаны құрайды.
Сонымен қатар, тін шетінде жасуша белсенділігі біріктірілетініндей олардың жүйесінің ағзалар жұмысы біріктірілуі тиіс. Бұл үшін гормондар мен жүйке жүйесі жауап береді. Әрі қарай көретініміздей, эволюция үрдісінде күрделі тіндер, ағзалар және ағза жүйелерінің пайда болуы дене құрылымы жоспарына маңызды өзгерістер алып келді және ақыры соңында транспортты жүйелер қажеттілігі туындады, соның ішінде қан өткізгіштік. Қан – бұл түгел ағзаны қан өткізгіш тамырлар мен жүректің қысқартылған түрінің арқасында айналушы сұйық тін.
Төменде біз бұл жолдағы маңызы зор болып келетін жануарлар патшалығының негізгі түр өкілдерін мысалға алып қарастырамыз.
2.8.2. Cnidaria түрі
Cnidaria – ның систематикасы мен негізгі белгілері 2.13 кестесінде берілген. Книдарийларға медузалар, актиниялар және маржандар жатады.
Қос қабатты жануарлар
Дененің құрылым жоспары қарапайым. Дене жасушаның қос қабатымен құрылған: сыртқысы – эктодерма және ішкісі – энтодерма. Бұл ұжымның қосқабатты дәрежесі деп аталады. Эктодермалды жасушалар сыртқы ортамен байланысады, ал
Ең бөлінісі, қағида бойынша, жұмыстың тиімділігін көтереді. Ұжым жеігейі тінге қарағанда әл де болса да биігірек болып саналады. тінге қатысты бірнеше жұмыскерлер ағза құрайды, мысалы, қарын. Ағзаларға қатысты жұмыскерлер тобы жүйе құрайды, сонымен қоса, ас қорыту сияқты. Әртүрлі барлық ағза жүйесінің байланастырылуы ағзаны құрайды.
Сонымен қатар, тін шетінде жасуша белсенділігі біріктірілетініндей олардың жүйесінің ағзалар жұмысы біріктірілуі тиіс. Бұл үшін гормондар мен жүйке жүйесі жауап береді. Әрі қарай көретініміздей, эволюция үрдісінде күрделі тіндер, ағзалар және ағза жүйелерінің пайда болуы дене құрылымы жоспарына маңызды өзгерістер алып келді және ақыры соңында транспортты жүйелер қажеттілігі туындады, соның ішінде қан өткізгіштік. Қан – бұл түгел ағзаны қан өткізгіш тамырлар мен жүректің қысқартылған түрінің арқасында айналушы сұйық тін.
Төменде біз бұл жолдағы маңызы зор болып келетін жануарлар патшалығының негізгі түр өкілдерін мысалға алып қарастырамыз.
2.8.2. Cnidaria түрі
Cnidaria – ның систематикасы мен негізгі белгілері 2.13 кестесінде берілген. Книдарийларға медузалар, актиниялар және маржандар жатады.
Қос қабатты жануарлар
Дененің құрылым жоспары қарапайым. Дене жасушаның қос қабатымен құрылған: сыртқысы – эктодерма және ішкісі – энтодерма. Бұл ұжымның қосқабатты дәрежесі деп аталады. Эктодермалды жасушалар сыртқы ортамен байланысады, ал

энтодермалды жасушалар ішекті немесе ішек қуысын төсей отырып, ішкісімен байланысады. Бұл қуыс сыртқы ортадан бір ғана тесік («ауыз») арқылы хабарланады. Қоректену ауызмаңы тесік айналасында орналасқан және дене қуысына қоректік бөлшектерді қуатын қармауыш арқылы алынады. Өнімдерді жұта бастау және қорытылмаған қалдықтарды шығару бұл бір ғана тесік арқылы жүреді. Жасуашлар кейбір деңгейге дейін мамандандырылған, сондықтан бұр берілген жағдайда тін деңгейіндегі ұжымдардың бар екені туралы айтуға болады. Эктодермада, мысалы нематобласттар деп аталатын күйдіргіш жасушалы топтары болатын қармауышы бар. Бұл жасушалар 3 типтиі жіпті шығарып тастайды, оның бірі – құрбандыққа қадалу, басқасы – оны ұстап тұрады және үшіншісі – өлтіреді. Эктодермада, сонымен қатар, жүйке жасушаларымен байланысатын сезімтал жасушалар бар; жүйке жасушалары мезоглеяда орналасатын байланыс желісін орнатады. Бұлшықеттерге тән қысқартылған жасушалар, талшық, дененің қозғлыс белсенділігін және қармауыштарды, сонымен қатар еркін жүзгіш медуза локомоциясын да қамтамасыз етеді. Энтодермада өнімді қорытатын және өнімдік заттарды сіңіру үшін мамандандырылған жасушалар бар.
Радиал симметрия
Книдарий денесі радиал симметриямен ерекшеленеді. Басқа сөзбен айтқанда, егер книдарий денесін кез келген 2 диаметрге бөліп тастағанда, онда бөлініп кеткен екі жарты бір біріне ұқсас болып келеді. Радиал симметрия, әдеттегідей, локомоцияға қабілетсіз ағзалармен байланысады. (2.8.8. бөлімді қараңыздар). Жануарлардың көпшілігі дененің тек қана созылған және сүйір формасымен тоқтап қалмай, оның үлкен арнайы әртүрлі бөлшектерін қамтамасыз етеді.
Полиптер, медузалар және көп пішінділік
Книдарий үшін дене құрылымының екі негізгі түрін белгілейді, олар полип пен медуза (2.47 сурет). Полипте цилиндрлік формалы болып келеді және бекітілген тіршілігі болады. Мұндай ағзалар үстіне бекітілген болып қалады, мысалы, құзда ұзына бойы өзінің барлық өмірі біткенше. Қозғалыс қабілеті оларда өте шектеулі немесе жоқ десек те болады. Медуза – қолшатырға ұқсайтын еркінжүзгіш жануар. Белгілі бір нақты книдарийлердің тіршілік кезеңдерінде ұрпақтардың алмасуы байқалады және сонда медузалар жануарлардың жаңа жерге ауысуларын қамтамасыз етеді (төменнен қараңыздар). Бұл жағдайда полиптер медузаларды тармақтай отырып, жыныссыз жолмен көбейеді, ал медузалар полиптен дамитын балаңқұртты түзе отырып, жынысты жолмен көбейеді. Жалғыз полиптер бір колонияда форма бойынша ажыратылады. Мысалы, олардың біреуі өнімді ұстап алуға мамандандырылған болса, басқасы – жыныссыз көбеюге мамандандырылған (төменнен қараңыздар). Бір дара түрінің ішінде екі немесе одан да көп пішінде болатын болса, көп пішінділік деп аталады.
Радиал симметрия
Книдарий денесі радиал симметриямен ерекшеленеді. Басқа сөзбен айтқанда, егер книдарий денесін кез келген 2 диаметрге бөліп тастағанда, онда бөлініп кеткен екі жарты бір біріне ұқсас болып келеді. Радиал симметрия, әдеттегідей, локомоцияға қабілетсіз ағзалармен байланысады. (2.8.8. бөлімді қараңыздар). Жануарлардың көпшілігі дененің тек қана созылған және сүйір формасымен тоқтап қалмай, оның үлкен арнайы әртүрлі бөлшектерін қамтамасыз етеді.
Полиптер, медузалар және көп пішінділік
Книдарий үшін дене құрылымының екі негізгі түрін белгілейді, олар полип пен медуза (2.47 сурет). Полипте цилиндрлік формалы болып келеді және бекітілген тіршілігі болады. Мұндай ағзалар үстіне бекітілген болып қалады, мысалы, құзда ұзына бойы өзінің барлық өмірі біткенше. Қозғалыс қабілеті оларда өте шектеулі немесе жоқ десек те болады. Медуза – қолшатырға ұқсайтын еркінжүзгіш жануар. Белгілі бір нақты книдарийлердің тіршілік кезеңдерінде ұрпақтардың алмасуы байқалады және сонда медузалар жануарлардың жаңа жерге ауысуларын қамтамасыз етеді (төменнен қараңыздар). Бұл жағдайда полиптер медузаларды тармақтай отырып, жыныссыз жолмен көбейеді, ал медузалар полиптен дамитын балаңқұртты түзе отырып, жынысты жолмен көбейеді. Жалғыз полиптер бір колонияда форма бойынша ажыратылады. Мысалы, олардың біреуі өнімді ұстап алуға мамандандырылған болса, басқасы – жыныссыз көбеюге мамандандырылған (төменнен қараңыздар). Бір дара түрінің ішінде екі немесе одан да көп пішінде болатын болса, көп пішінділік деп аталады.
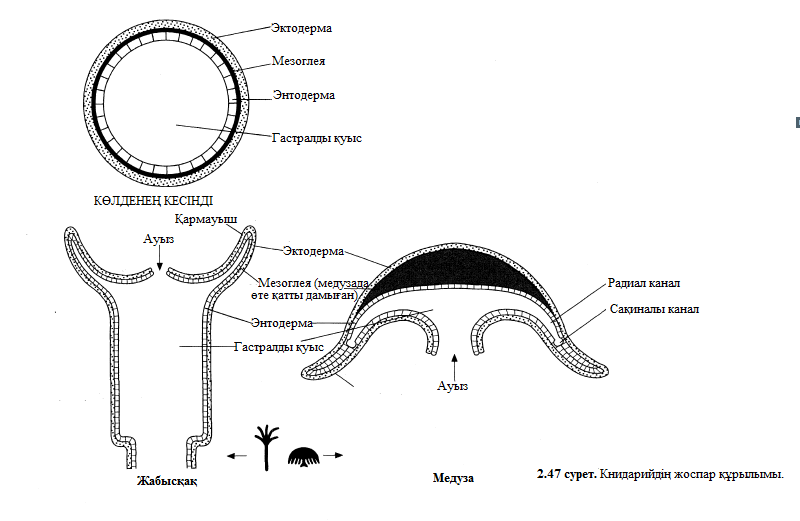
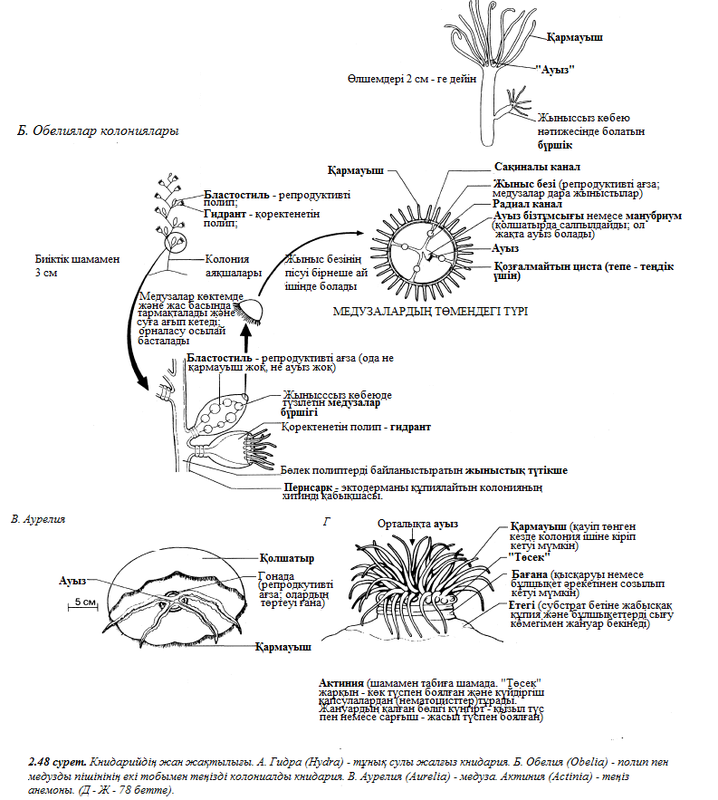
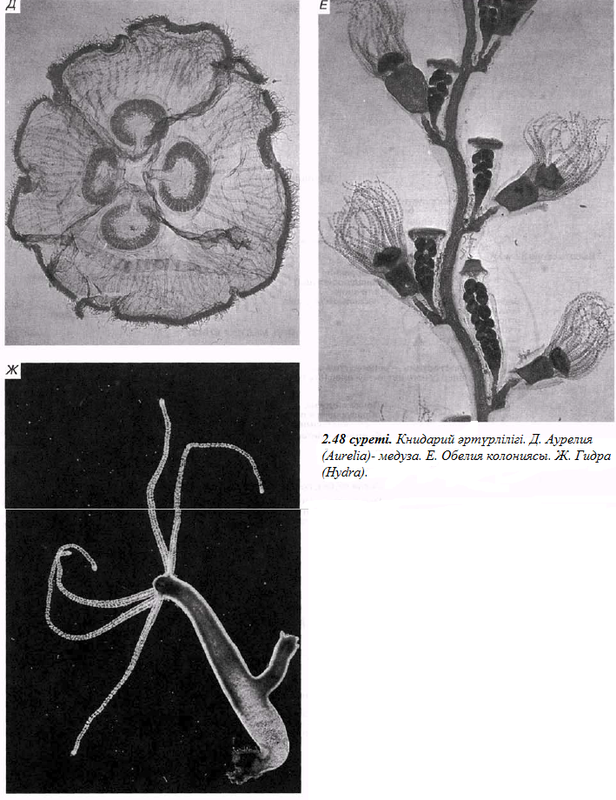
Өлшемдер
Книдарийлер кішігірім жануарлар. Аз ірі медузалар негізінде түзілуінде тірі жасушалар болмайтын мезоглеядан тұрады. Қорек заттары екі қабаттың ішінде, яғни энтодерма мен эктодермада тез диффуздалады. Онымен қоса, книдарий жасушалары диффузия жолымен болатын өте тиімді газ алмасу мүмкіндігін 2.48 суретінде әртүрлі книдарийлер берілген: суда мекендейтін обелия, аурелия мен актиния. Обелияның тіршілік кезеңінің мысалына полиптердің колониалды ұрпағы кіші медузалар ұрпақтарымен байланысатын көп пішінділікті оңай үйлестіруге болады. Олар әдетте құзға, қабыршақта, теңіз балдырларының жапырақтарнынажәне т.б –ларына бекіне отырып, таяз суда мекендейді. Актиния әдетте Ұлыбританияның жағалау жақтарында кездеседі, әсіресе, жылға жақтарда және құз жақтағы өзен жағалауларында.
2.8.3. Platyhelminthes түрі (жазық құрттар)
Platyhelminthes – тің системтикасы мен ерекше белігілері 2.14 кестесінде берілген.
Книдарийлер кішігірім жануарлар. Аз ірі медузалар негізінде түзілуінде тірі жасушалар болмайтын мезоглеядан тұрады. Қорек заттары екі қабаттың ішінде, яғни энтодерма мен эктодермада тез диффуздалады. Онымен қоса, книдарий жасушалары диффузия жолымен болатын өте тиімді газ алмасу мүмкіндігін 2.48 суретінде әртүрлі книдарийлер берілген: суда мекендейтін обелия, аурелия мен актиния. Обелияның тіршілік кезеңінің мысалына полиптердің колониалды ұрпағы кіші медузалар ұрпақтарымен байланысатын көп пішінділікті оңай үйлестіруге болады. Олар әдетте құзға, қабыршақта, теңіз балдырларының жапырақтарнынажәне т.б –ларына бекіне отырып, таяз суда мекендейді. Актиния әдетте Ұлыбританияның жағалау жақтарында кездеседі, әсіресе, жылға жақтарда және құз жақтағы өзен жағалауларында.
2.8.3. Platyhelminthes түрі (жазық құрттар)
Platyhelminthes – тің системтикасы мен ерекше белігілері 2.14 кестесінде берілген.
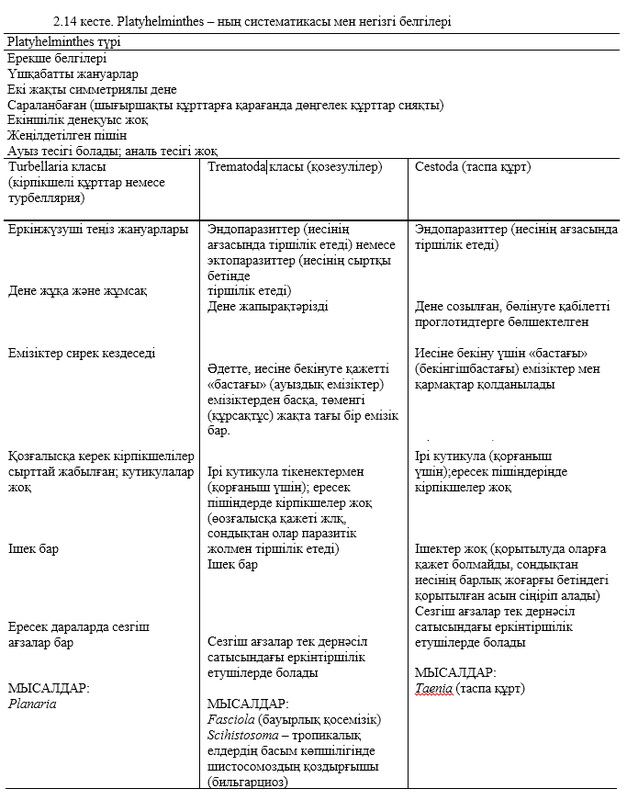
Жазық құрттың үшқабатты құрылымы экто – мен энтодерма арасында орналасқан жапырақтың үшінші ұрығы мезодерма ұрығының пайда болуымен қамтамасыз етіледі (2.55 суреті, А). Мезодерманың болуы бірнеше қарым – қатынастарда маңызды орын алады.
1. Дененің үшқабатты құрылымды өлшемінде кеңеюге болады. нәтижесінде жануар денесінің қабырғасынан қорытылмалы жол жойылған байқалады.
2. Мизодермадан әртүрлі ағзалар қалыптасады, келісілген жұмыс ұжымның жоғары дейгейіне дейін әкеледі – ағза жүйесінің дамуына. Мұндай жүйеге орталық жүйке жүйесі, қорыту ағзаларының жүйесі, бөліну және көбею мысалы бола алады.
3. Үшқабатты жануарларда бұлшықеттер жұмысы айтарлықтай жетілген болып саналады. бұл жерде керегі жануарлар өлшемдерінің үлкейтілуімен байланысты, сондықтан кірпікшелер мен жалған аяқтар енді локомоторлы функцияларды қамтамасыз ете алмайды.
Бірақ жануар өлшемдерінің кеңейтілуі эндодерма мен эктодерма қабаттарының арасындағы заттардың транспорт мәселесін тудырады. Кейбір жануарларда мезодерма эктодерма мен энтодерма арасындағы кеңістікті толтырып тастайды (ацеломикалық жануарлар; 2.55; А суреті). Бұл жағдайда көлік жағдайлары үлкен беттің ауданы оның көлемімен сақталуын қамтамасыз ететін дене жайпақтығымен шешіледі. Қарапайым диффузиялар нәтижесінде қоршаған ортамен жануарлар тіндерінің арасындағы заттардың ауысуының керек дәрежесін ұстап қалуы жеткілікті. Мезодермасында қуыс – жалпылай (жалпыламалық жануарлар) толығымен өсетін өзге жануарларда бір дене екінші денеге ауысатын заттардың тасымалын қамтамасыз ететін көлік жүйелері шықты.
Жазық құрттарда үшқабатты құрылым болып келеді. Бұл мезодермадан түзілетіналғашқы ағзалар мен ағзалар жүйесі болған өте ертедегі жануарлар тобы. Жазық құрттар – ацеломикалық жануарлар, зерттеу бойынша олардың атауы олардың денесіне қарай қойылған, яғни денесі – жазық болып келеді. Мезодерманың негізгі массасы денеде сараланбаған болып қалады және ішкі ағзадағы тіректі және қорғаныш функциясын орындайтын құрсақты тін – мезенхиманы түзеді.
Turbellaria класы
Планария (planaria) – еркін тіршілік етуші, тұщы өзендерде және тоғандарда кездесетін жазық құрт. Күндіз планария тас астарында тығылады, ал түнде аң аулайды. Ол қара түсті және ұзындығы 15 мм – ге жетеді. Планарияның денесі, яғни үлкен алдыңғы «бас» жәе өткір соңына байланысты ұзын және өте қатты жымырайған денесі бар. «Баста» сыртқы жақтан көз жұптары орналастыратын екіжақты симметрия тән (2.49 сурет).
Соңғы жағына жақын орналасқан құрсақтұс жағында ішектің сыртқы жағы ортамен байланыс жасайтын жалғыз тесігі ауыз орналасқан. Планария өнімі шаянтәрізді ұсақ құрттардан және ірілеу ағзалардың қалдықтарын шірітетіннен тұрады.
1. Дененің үшқабатты құрылымды өлшемінде кеңеюге болады. нәтижесінде жануар денесінің қабырғасынан қорытылмалы жол жойылған байқалады.
2. Мизодермадан әртүрлі ағзалар қалыптасады, келісілген жұмыс ұжымның жоғары дейгейіне дейін әкеледі – ағза жүйесінің дамуына. Мұндай жүйеге орталық жүйке жүйесі, қорыту ағзаларының жүйесі, бөліну және көбею мысалы бола алады.
3. Үшқабатты жануарларда бұлшықеттер жұмысы айтарлықтай жетілген болып саналады. бұл жерде керегі жануарлар өлшемдерінің үлкейтілуімен байланысты, сондықтан кірпікшелер мен жалған аяқтар енді локомоторлы функцияларды қамтамасыз ете алмайды.
Бірақ жануар өлшемдерінің кеңейтілуі эндодерма мен эктодерма қабаттарының арасындағы заттардың транспорт мәселесін тудырады. Кейбір жануарларда мезодерма эктодерма мен энтодерма арасындағы кеңістікті толтырып тастайды (ацеломикалық жануарлар; 2.55; А суреті). Бұл жағдайда көлік жағдайлары үлкен беттің ауданы оның көлемімен сақталуын қамтамасыз ететін дене жайпақтығымен шешіледі. Қарапайым диффузиялар нәтижесінде қоршаған ортамен жануарлар тіндерінің арасындағы заттардың ауысуының керек дәрежесін ұстап қалуы жеткілікті. Мезодермасында қуыс – жалпылай (жалпыламалық жануарлар) толығымен өсетін өзге жануарларда бір дене екінші денеге ауысатын заттардың тасымалын қамтамасыз ететін көлік жүйелері шықты.
Жазық құрттарда үшқабатты құрылым болып келеді. Бұл мезодермадан түзілетіналғашқы ағзалар мен ағзалар жүйесі болған өте ертедегі жануарлар тобы. Жазық құрттар – ацеломикалық жануарлар, зерттеу бойынша олардың атауы олардың денесіне қарай қойылған, яғни денесі – жазық болып келеді. Мезодерманың негізгі массасы денеде сараланбаған болып қалады және ішкі ағзадағы тіректі және қорғаныш функциясын орындайтын құрсақты тін – мезенхиманы түзеді.
Turbellaria класы
Планария (planaria) – еркін тіршілік етуші, тұщы өзендерде және тоғандарда кездесетін жазық құрт. Күндіз планария тас астарында тығылады, ал түнде аң аулайды. Ол қара түсті және ұзындығы 15 мм – ге жетеді. Планарияның денесі, яғни үлкен алдыңғы «бас» жәе өткір соңына байланысты ұзын және өте қатты жымырайған денесі бар. «Баста» сыртқы жақтан көз жұптары орналастыратын екіжақты симметрия тән (2.49 сурет).
Соңғы жағына жақын орналасқан құрсақтұс жағында ішектің сыртқы жағы ортамен байланыс жасайтын жалғыз тесігі ауыз орналасқан. Планария өнімі шаянтәрізді ұсақ құрттардан және ірілеу ағзалардың қалдықтарын шірітетіннен тұрады.
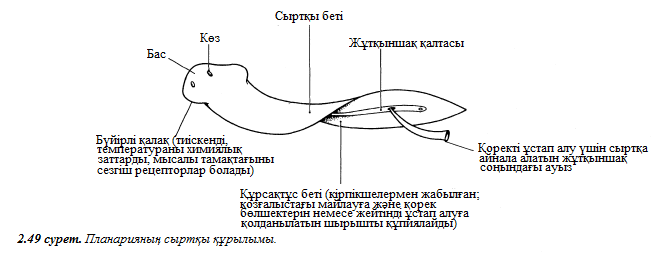
|
Trematodа класы
Бауырсорғыш (Fasicola hepatica); (2. 50 сурет), жануарлар патшалығында негізгі паразиттік топтарды құрайтын Trematodа класына жатады. Бауырсорғыш – эндопаразит, яғни ол иесінің ағзасында тіршілік етеді. Оның қоректену жері болып иесінің алғашқы не болмаса соңғы қойының өт ағыны қызмет етеді. Алғашқы иесі ретінде ірі мүйізді мал жатуы мүмкін, ал кейде адам қос емізік пен еркін тіршілік етуші планарияның арасында көп айырмашылық бар. Бұл айрымашылықтар, эндопаразит сияқты тіршілік етуге қажетті бейімдеушіліктердің дамуымен қамтамасыз етілуі мүмкін. Паразиттік тіршілік өмірі ұрпақ санын тіршілік кезеңінде көбейтуге қабілетті 3 дернәсіл сатыларынан (мирцидий, редия, церкария) тұратын күрделі ьіршілік кезеңімен байланысты. Көпсанды ұрпақ иесінің болмай қалатын ауысымында өлімнің үлкен дәрежесін жауып тастауға әкеледі. Бауырсорғыштардың тіршілік кезеңінің бөлігі екіншілік иесі бұл паразиттің дернәсілдік сатыларында өсіп және көбеюлеріне болатын белгілі бір уақыттарда (аралық) тұщы сулы үйрек Limnaea ағзасына қарай ығысады. Қос емізіктердің тіршілік кезеңінің барлық сатыларындапаразиттік тіршілік өміріне морфологиялық, физиологиялық және репродуктивті бейімделгіштерді белгілеуге болады. Олардың кейбіреулері 2.50 суретінде берілген. Бауырсорғыштардың тіршілік кезеңі 2.50 суретте көрсетілген. Ересек қосезулілер. Қосезулілер денесіжеңілдетілген және ішігірім өт ағысы тіршілігіне бейімделген. Дене қабырғасы қосезуліліні иесінің ферменттер әсерелерінен қорғайды, ал безді жасушалардың қабырғада орналасу құпиясы иесін анттитоксин әсерінен қорғайды. |
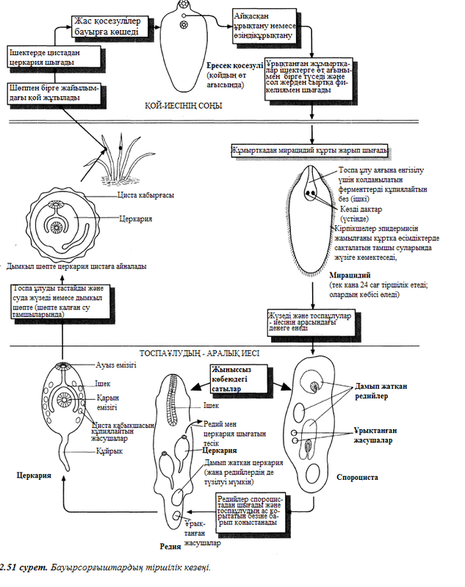
|
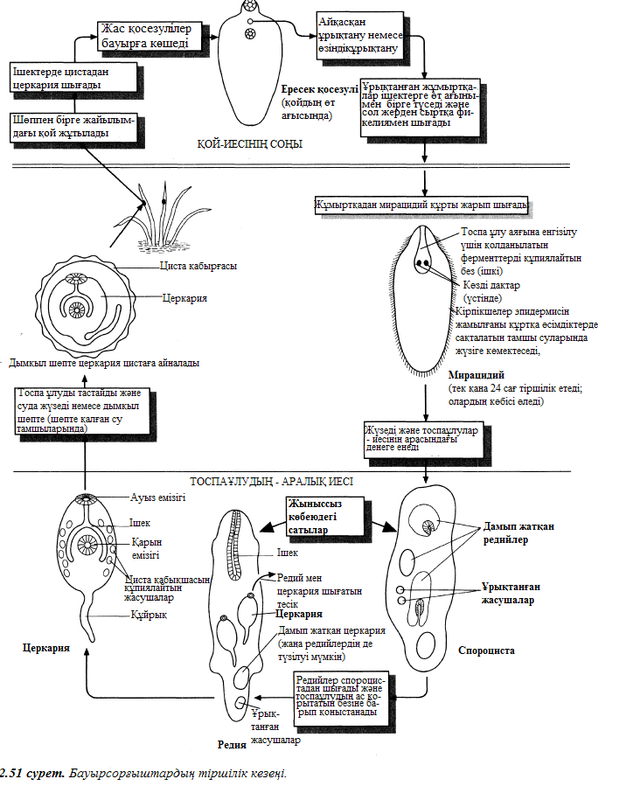
Қос жынысты жүйесі ағзаларының көбеюі (сол және басқа да ағзада аталық және аналық ағзаларының болуы) өзіндік ұрықтану мүмкіндігін қамтамасыз етеді және айқасқанұрықтануд да. Бауырсорғыш оттек жетіспесе де тіршілік ете беруге де қабілетті.
Мирацидий. Мирацидий – бұл қос езулілердің даму кезеңіндегі алғашқы дернәсіл сатысы (2.51 суреті). Мирацидияның негізгі функциясы – қозғалысқа қабілетті сезім ағзаларының бар болуы қажет етілетін аралық иесінің іздеуі. Одан басқа, мирацидий жаңа дернәсілдерді (спорацистерді, төменде қраңыздар) туындатады. Мирацидия эпидермисі кірпікшелермен жабылғаны бұл дернәсілге суда немесе өсімдіктерде қалып қойған дым тамшыларынжа мүмкіндік береді.
Белгілі химиялық заттардың әсерінен мирацидийлер өзінің аралық иесі – тоспа ұлуларға жүзеді. (Хемотаксис деп аталатын химиялық қоздырғыш жаққа ауысу қабілеттілігі). Мирацидий бүршіктің жоғарғы жағы көмегімен тоспаұлу аяғына бекінеді, ал аяқ бетіндегі үстіңгі безді құпиялайтын протеолитикалық ферменттер иесінің тіндеріне мирациядияның таралуын жеңілдетеді. Әрі қарай енуі тоспаұлудың денесі кіріп кетуді жеңілдететін бұлшықет жасушалары көмегімен жүреді. Осылайша, мирациядияның миграциясы тоспаұлудың асқорыту өтінде жүреді. Мирацийде келесі дернәсіл пішіндері дамитын арнайы ұрпақ жасушалары болады.
Спороцист. Бұл сатының рөлі өзіне таппаған дернәсіл бөлігімен байланысты жоғалудың орнын толтыру үшін дернәсіл сатысы – редиядан дамитын ұрпақ жасушалары бар қозғалмайтын бекітуші қап.
Редия. Бұл – көбею мен қорек сатысы. Бұл сатыдағы дернәсілдерде иесінің тіндерінен сұйықтарды сіңіретін бұлшықетті жұтқыншақ болады. Редийдің қозғалыстары бұлшықет жасушаларының көмегімен болады. Ұрықтанған жасушалар редиялар мен церкарияларды туындатады. Редия мен церкарийдің жаңа ұрпағы арнайы тесік арқылы сыртқа шығады.
Церкария. Бұл көбінесе ересек қосезулілерге ұқсайтын дернәсіл. Сол, атап айтқанда, ода сәйкес субстратқа, мысалы, шөпке бекінетін ауызжәне құрсақ емізіктері бар. Бірақ одан басқа, ода суда және өсімдік суларының тамшыларында қозғалуға көмектесетін құйрығы бар. Бұл дернәсілдерде циста қабатын құпиялайтын бездер болады (2.51 сурет). Церкария цистаға айналмай қойға жұтылып қоймаса, оның әрі қарай дамуы жүрмейді. Цистаға айналған церкариялар төмен температураға ұлкен тұрақтылықты иемденеді, біраз құрғақшылыққа сезімтал.
Limnаea – қосмекенді тіршілік ететін және тоғандарда, өзен лайлы жерлерінде және шығанақ көгалдарында мекендейтін тоспаұлу. Ол жайсыз жағдайлардың тасымалдауға қабілетті, сондықтан оның денесінде дамитын спорацистер мен редиялар қос езулілердің дернәсіл пішіндері мұндай жайсыз жағдайларда қорғанышты бола алады. Және шынымен төмен температураларда редиялар церкария емес, еншілес редийлерді түзеді. Редия тоспаұлу денесінде қалады және сонда қыстап шығуына болады. Жылы ауа – райы басталған кезде көктемде ғана церкарийлер дамып бастайды. Тоспаұлу да өте тез көбейеді. 12 аптада бір ұлу 160 000 дараға жететін ұрпақ беретіні саналған. Егер осы барлық ұрпақ қосезулілер сатысында дамитындарға жұқтыратын болса, онда жаңа біткен иелерді бүлдіруге қабілетті церкарийдің шығу ықтималдылығы сыртқы орта бойынша кәдімгідей өседі. Limnаea – ның қосмекендік тіршілігі олар ммекендіріле алатын церкарийдің ылғалды ортада түсуіне кепілдік береді.
Жас қос езулілердің мегацеркарийден босап шығуы қой немесе сиырлар ішегінде болады. Бұл үрдіс көміртек диоксиді мен үлкен (39 ) температураның үлкен ұстанымымен асқазанда бастама көрсетеді. Мұндай жағдайларда паразит циста тесігінде оның қабырғасын қорытатын протеин ыдыратушы ферменттерді бөліп шығарады. Жас қосезулілердің цистадан шығуы кішкентай ішектің қорытатын шырындарында өттің бар болуымен бастамашылық етеді.
Босап шыққан жас қоз езулілер ішек қабырғасы арқылы бауырға ауысады. Бір уақыттарда олар бауыр тінімен қоректеп, бірақ сосын, шамамен зақымданудан кейін 6 апта өткен соң мәңгілікке өт ағынына ауысады.
Қосезулілердің зақымдану әсері тіршілік иесіне өте әртүрлі. Қосезулілердің ауыр жағдайларда зақымдануы иесіне бүліншік шақыруы мүмкін. Церкарийдің бауыр арқылы жүрген миграциясы бұл ағзадағы заттардың алмасуына бұзылу әкеледі. Нәтижесінде, бауыр жасушаларында бұзылу болады және тиісінше, өт ағындарында бітеліп қалады, бауырдың (бауыр циррозы) ірі жарасы ішбөсірді дамуына әкеледі. Өттің жоқтығы толық немесе жеткіліксіз саны ішектерде асқорытылуды бұзады, ал қосезулілерде бөлініп шыққан өнімдер иесіне таксиндік әрекеттерді білдіртуі мүмкін.
Қосезулілер күресіне әртүрлі әдістер қолданылады. Жайылымдық жерлердің құрғатылуы және өрісте ұлуларды (күрестің биологиялың әдісі) қаз бен үйректің жайылуы аралық иесі – тоспаұлудың (Limnаea) құртылып кетуіне бағытталған. Тиімді тәсіл ретінде жердің әктігін алуға да болады, сондықтан сілтілі ортада (рН 7,5) паразит жұмыртқасы дами алмайды. Зарарлы қойды бауырсорғыштардан арылту үшін жануарларға бауырда қосезулілерді тіке өлтіретін тетрахлоридті көміртек жүргізіледі.
2.8.4. Nematoda типі (нематодтар немесе жұмыр құрттар)
Нематодтар – бұл кішігірім цилиндрлі дене және ұштары өткір болып келетін ең төмен құрттар. Жазық құрттар, олап үшқыбытты құрлым және қан жүретін жүйесінен айырылған болып келеді. Оларды не целомикалық, не ацеломикалық жануарлармен санауға болмайды (2.8.5 бөлім); олардың ішкі құрылым өте қарапайым және мұнда оларды қарастырудың мәні жоқ. Нематодтар кезбе болып қалды: олар суда, жерде (100 000 млн. гектарға дейін) және ең әртүрлі басқа да мекендейтін жерлерде кездеседі. Бірақ, олардың көбісінде өзінде микроскопиялық немесе өте аз, себебі оларды кенеттен оңай көре алмайсыз. Әрдайым олардың жаңа түрлері шығып жатады; олардың бірі, Германиядағы сыра өндірісінде қолданылатын сыра ашытқысының бірден – бірі болып табылады. Жұмыр құрттар, трематодтар және таспақұрттар паразит жануарларының ірі 3 тобын түзеді. 2.15 кестесінде жұмыр құрттардың ерекше белгілері берілген. Нематод мысалы болып аскарид бола алады (Ascaris; 2.52 сурет).
Мирацидий. Мирацидий – бұл қос езулілердің даму кезеңіндегі алғашқы дернәсіл сатысы (2.51 суреті). Мирацидияның негізгі функциясы – қозғалысқа қабілетті сезім ағзаларының бар болуы қажет етілетін аралық иесінің іздеуі. Одан басқа, мирацидий жаңа дернәсілдерді (спорацистерді, төменде қраңыздар) туындатады. Мирацидия эпидермисі кірпікшелермен жабылғаны бұл дернәсілге суда немесе өсімдіктерде қалып қойған дым тамшыларынжа мүмкіндік береді.
Белгілі химиялық заттардың әсерінен мирацидийлер өзінің аралық иесі – тоспа ұлуларға жүзеді. (Хемотаксис деп аталатын химиялық қоздырғыш жаққа ауысу қабілеттілігі). Мирацидий бүршіктің жоғарғы жағы көмегімен тоспаұлу аяғына бекінеді, ал аяқ бетіндегі үстіңгі безді құпиялайтын протеолитикалық ферменттер иесінің тіндеріне мирациядияның таралуын жеңілдетеді. Әрі қарай енуі тоспаұлудың денесі кіріп кетуді жеңілдететін бұлшықет жасушалары көмегімен жүреді. Осылайша, мирациядияның миграциясы тоспаұлудың асқорыту өтінде жүреді. Мирацийде келесі дернәсіл пішіндері дамитын арнайы ұрпақ жасушалары болады.
Спороцист. Бұл сатының рөлі өзіне таппаған дернәсіл бөлігімен байланысты жоғалудың орнын толтыру үшін дернәсіл сатысы – редиядан дамитын ұрпақ жасушалары бар қозғалмайтын бекітуші қап.
Редия. Бұл – көбею мен қорек сатысы. Бұл сатыдағы дернәсілдерде иесінің тіндерінен сұйықтарды сіңіретін бұлшықетті жұтқыншақ болады. Редийдің қозғалыстары бұлшықет жасушаларының көмегімен болады. Ұрықтанған жасушалар редиялар мен церкарияларды туындатады. Редия мен церкарийдің жаңа ұрпағы арнайы тесік арқылы сыртқа шығады.
Церкария. Бұл көбінесе ересек қосезулілерге ұқсайтын дернәсіл. Сол, атап айтқанда, ода сәйкес субстратқа, мысалы, шөпке бекінетін ауызжәне құрсақ емізіктері бар. Бірақ одан басқа, ода суда және өсімдік суларының тамшыларында қозғалуға көмектесетін құйрығы бар. Бұл дернәсілдерде циста қабатын құпиялайтын бездер болады (2.51 сурет). Церкария цистаға айналмай қойға жұтылып қоймаса, оның әрі қарай дамуы жүрмейді. Цистаға айналған церкариялар төмен температураға ұлкен тұрақтылықты иемденеді, біраз құрғақшылыққа сезімтал.
Limnаea – қосмекенді тіршілік ететін және тоғандарда, өзен лайлы жерлерінде және шығанақ көгалдарында мекендейтін тоспаұлу. Ол жайсыз жағдайлардың тасымалдауға қабілетті, сондықтан оның денесінде дамитын спорацистер мен редиялар қос езулілердің дернәсіл пішіндері мұндай жайсыз жағдайларда қорғанышты бола алады. Және шынымен төмен температураларда редиялар церкария емес, еншілес редийлерді түзеді. Редия тоспаұлу денесінде қалады және сонда қыстап шығуына болады. Жылы ауа – райы басталған кезде көктемде ғана церкарийлер дамып бастайды. Тоспаұлу да өте тез көбейеді. 12 аптада бір ұлу 160 000 дараға жететін ұрпақ беретіні саналған. Егер осы барлық ұрпақ қосезулілер сатысында дамитындарға жұқтыратын болса, онда жаңа біткен иелерді бүлдіруге қабілетті церкарийдің шығу ықтималдылығы сыртқы орта бойынша кәдімгідей өседі. Limnаea – ның қосмекендік тіршілігі олар ммекендіріле алатын церкарийдің ылғалды ортада түсуіне кепілдік береді.
Жас қос езулілердің мегацеркарийден босап шығуы қой немесе сиырлар ішегінде болады. Бұл үрдіс көміртек диоксиді мен үлкен (39 ) температураның үлкен ұстанымымен асқазанда бастама көрсетеді. Мұндай жағдайларда паразит циста тесігінде оның қабырғасын қорытатын протеин ыдыратушы ферменттерді бөліп шығарады. Жас қосезулілердің цистадан шығуы кішкентай ішектің қорытатын шырындарында өттің бар болуымен бастамашылық етеді.
Босап шыққан жас қоз езулілер ішек қабырғасы арқылы бауырға ауысады. Бір уақыттарда олар бауыр тінімен қоректеп, бірақ сосын, шамамен зақымданудан кейін 6 апта өткен соң мәңгілікке өт ағынына ауысады.
Қосезулілердің зақымдану әсері тіршілік иесіне өте әртүрлі. Қосезулілердің ауыр жағдайларда зақымдануы иесіне бүліншік шақыруы мүмкін. Церкарийдің бауыр арқылы жүрген миграциясы бұл ағзадағы заттардың алмасуына бұзылу әкеледі. Нәтижесінде, бауыр жасушаларында бұзылу болады және тиісінше, өт ағындарында бітеліп қалады, бауырдың (бауыр циррозы) ірі жарасы ішбөсірді дамуына әкеледі. Өттің жоқтығы толық немесе жеткіліксіз саны ішектерде асқорытылуды бұзады, ал қосезулілерде бөлініп шыққан өнімдер иесіне таксиндік әрекеттерді білдіртуі мүмкін.
Қосезулілер күресіне әртүрлі әдістер қолданылады. Жайылымдық жерлердің құрғатылуы және өрісте ұлуларды (күрестің биологиялың әдісі) қаз бен үйректің жайылуы аралық иесі – тоспаұлудың (Limnаea) құртылып кетуіне бағытталған. Тиімді тәсіл ретінде жердің әктігін алуға да болады, сондықтан сілтілі ортада (рН 7,5) паразит жұмыртқасы дами алмайды. Зарарлы қойды бауырсорғыштардан арылту үшін жануарларға бауырда қосезулілерді тіке өлтіретін тетрахлоридті көміртек жүргізіледі.
2.8.4. Nematoda типі (нематодтар немесе жұмыр құрттар)
Нематодтар – бұл кішігірім цилиндрлі дене және ұштары өткір болып келетін ең төмен құрттар. Жазық құрттар, олап үшқыбытты құрлым және қан жүретін жүйесінен айырылған болып келеді. Оларды не целомикалық, не ацеломикалық жануарлармен санауға болмайды (2.8.5 бөлім); олардың ішкі құрылым өте қарапайым және мұнда оларды қарастырудың мәні жоқ. Нематодтар кезбе болып қалды: олар суда, жерде (100 000 млн. гектарға дейін) және ең әртүрлі басқа да мекендейтін жерлерде кездеседі. Бірақ, олардың көбісінде өзінде микроскопиялық немесе өте аз, себебі оларды кенеттен оңай көре алмайсыз. Әрдайым олардың жаңа түрлері шығып жатады; олардың бірі, Германиядағы сыра өндірісінде қолданылатын сыра ашытқысының бірден – бірі болып табылады. Жұмыр құрттар, трематодтар және таспақұрттар паразит жануарларының ірі 3 тобын түзеді. 2.15 кестесінде жұмыр құрттардың ерекше белгілері берілген. Нематод мысалы болып аскарид бола алады (Ascaris; 2.52 сурет).

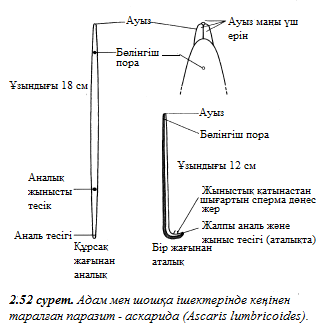
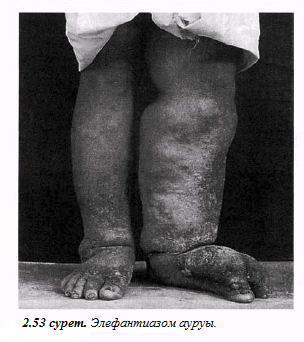
Чтобы изменить, нажмите здесь.
Түрдің бір түрі, Ascaris lumbricoides, адам мен шошқаның әдеттегі паразиті болып табылады. бұл өте ірі, шамамен жұмыр құрт сияқты өлшемде (ұзындығы 20 – см – ге дейін), нематодта ақсары – ақ түс болады. Ауыр жағдайларда аскаридтердің (аскаридоз) зақымдалуы ішек бітелуіне әкелуі мүмкін.
Фекалиямен бірге сыртқа бөлініп шығатын аскарид жұмыртқалары өте тіректі және жылдар бойы тіршілік қабілеттіктерін жоғалтпайды. Аскарид аталықтары аналықтардан кішілеу және соңғы жағы бұралған. Басқа жақсы белгілі паразиттегіш нематод – Wuchereria bancroftii – шораяқты (элефантиаз) шақырта отырып, лимфатикалық жүйеге зақым келтіреді (2.53 сурет). Бұл жағдайда аяқтары пілге ұқсай отырып шектен тыс үлкейеді. Нематодтар әртүрлі астықтарды қоса отырып, өсімдіктерді де зақымдауы мүмкін.
2.8.5. Annelida типі (анелидтер немесе сақина тәрізді құрттар)
2.16 кестесінде сақина тәрізді құрттардыңсистематикасы мен ерекше белгілері 2.54 суретінде әртүрлі аналид мысалдары берілген. Оның екеуі, Nereis және Lumbricus астында толығырақ қаралады. Аннелидтер – целомикалық жануарлар.
Фекалиямен бірге сыртқа бөлініп шығатын аскарид жұмыртқалары өте тіректі және жылдар бойы тіршілік қабілеттіктерін жоғалтпайды. Аскарид аталықтары аналықтардан кішілеу және соңғы жағы бұралған. Басқа жақсы белгілі паразиттегіш нематод – Wuchereria bancroftii – шораяқты (элефантиаз) шақырта отырып, лимфатикалық жүйеге зақым келтіреді (2.53 сурет). Бұл жағдайда аяқтары пілге ұқсай отырып шектен тыс үлкейеді. Нематодтар әртүрлі астықтарды қоса отырып, өсімдіктерді де зақымдауы мүмкін.
2.8.5. Annelida типі (анелидтер немесе сақина тәрізді құрттар)
2.16 кестесінде сақина тәрізді құрттардыңсистематикасы мен ерекше белгілері 2.54 суретінде әртүрлі аналид мысалдары берілген. Оның екеуі, Nereis және Lumbricus астында толығырақ қаралады. Аннелидтер – целомикалық жануарлар.
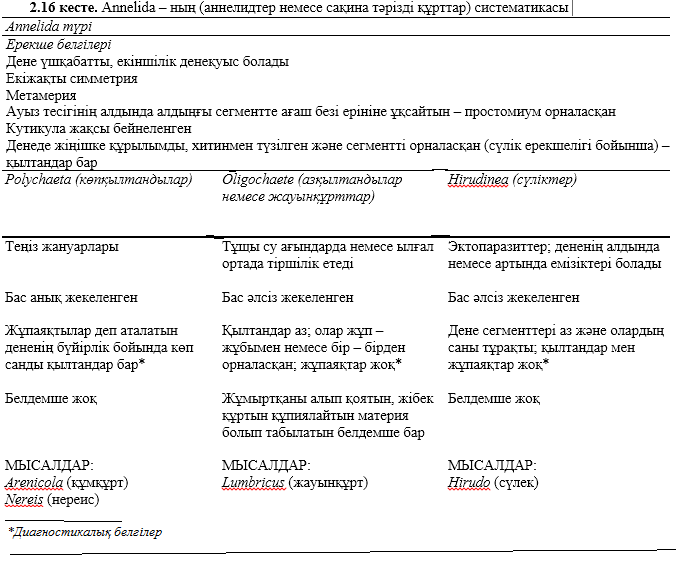
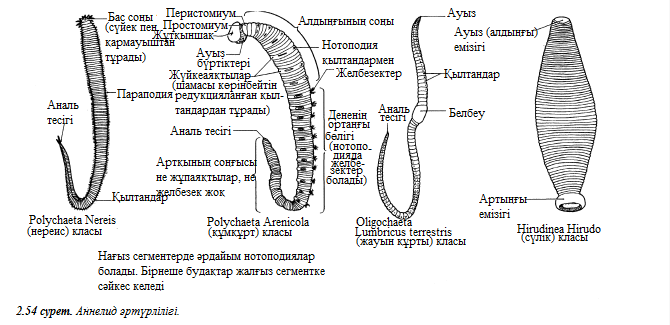
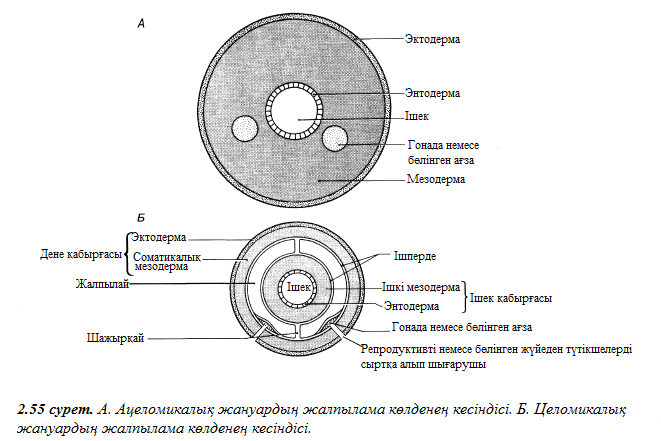
Дене құрылымының целомикалық типі
Өзіміз көргеніміздей, жазық құрттарда мезодерма тығыз орташа қабықты құрай отырып, эктодерма мен энтодерма арасындағы жапырақта кеңістікті толығымен толтырады. Дене құрылымында екіншілік денеқуыс жоқ болатын болса, онда ацеломикалық деп аталады (2.55 суреті, А).
Аннелидте жоғары бірлескен жануарлар тобында екіншілік қуыс деп аталатын дене қуысы құрылады. Ол мезодермада эмбриондық үрдіс кезінде саңылау сияқты пайда болады. Мұндайдан түзілген қуыс мезодерманы екі жапыраққа бөле отырып, сыртқа қаралғаны – соматикалық, ішке қарағаны – ішкілік (2.55 сурет, Б) деп аталатын целомикалық сұйықтықты толтырады. Эктодермамен байланысатын соматикалық мезодерма дене қабырғасын түзеді. Энтодермамен байланысатын ішкілік мезодерма ішектің бұлшықетті қабырғасын түзеді. Осылайша, екіншілік денеқуыс ішек қабырғасынан дене қабырғасын бөледі.
Мезодерманың екіншілік денеқуыспен төселген негізгі бөлігі бұлшықеттерде дамиды; дене қабырғасын құрайтын бұлшықеттер жануардың локомоциясын қамтамасыз етеді, ал ішек қабырғасының бұлшықеттері соңғысының толқып жиырылғанын қамтамасыз етеді. Дене қабырғасынан ішек қабырғасына дейінгі Дене құрылымының целомикалық типі
Өзіміз көргеніміздей, жазық құрттарда мезодерма тығыз орташа қабықты құрай отырып, эктодерма мен энтодерма арасындағы жапырақта кеңістікті толығымен толтырады. Дене құрылымында екіншілік денеқуыс жоқ болатын болса, онда ацеломикалық деп аталады (2.55 суреті, А).
Екіншілік денеқуыстың биологиялық мәні
Целомикалық жануарлардың дамуындағы тағы бір эволюционды артықшылық болып табылатын метамерия немесе сегментация пайда болды, - жануар денесіндегі көлденең кеңістіктердің бірнеше бірдей бөлшектерге немесе сегменттерге бөлінген ұжым деңгейі. Басқа сөздермен айтқанда, жануар денесі өзіндік бірдей сегментте бір – бірімен орналасқан ұзын қатарлы болады. Сегментация мезодермадан бастау алады, бірақ әдетте мизодермалды және эктодермалды дененің бөліктерін қамтиды.
Анығырақ метамерия бұл бөлініс сырттан жақсы көрінетін (дененің барлық ұзына бойы көшірілуі) сақина тәрізді құрттарда пайда болады (аннелид). Ішкі сегменттер бір бірінен екіншілік денеқуыс арқылы жүретін кеңістіктермен бөлінеді. Әрбір сегментте өзінің өзіндік бұлшықеттер жинағы, қан жүретін тамырлары, жүйке жасушалары, ал кейбіреулерінде репродуктивті ағзалардың топтары болады. Бірақ тіпті анналидтің өзінде сегменттер бір бірінен толығымен тәуелсіз емес, сондықтан жүйкелі, әсіресе қан жүретін жүйелер жануарлар денесінің барлық бойынан өтіп шығуы керек.
Әрбір сегменттің сегментациясы мен құрылымының негізгі жоспары эволюционды бекітілген бойда, әрі қарайғы бөлек сегменттердің немесе олардың кішігірім топтары бойынша, сонымен қатар, тағы бір мүмкіндік енсіз мамандану мен әртүрлі дене бөлшектерінің еңбек бөлінісі сияқты эволюционды өзгерістердің мүмкіндігі пайда болды. Бұл бірнеше жол арқылы болады. Әртүрлі функциялар әртүрлі сегменттермен орындалуы мүмкін; басты қалыптастыру кезінде бірнеше сегменттер қосылғандағы цефализация болғанда, сегменттердің қосылуы да мүмкін болды; бауыраяқтыларда кейбір сегменттердің жоғалуы да байқалады. Бірақ, бунақаяқтылардың ішкі сегментациясы аннелид сияқты анық айтылған. Желілілерде сыртқы сегментациясы құртылған, бірақ кейбір жүйелер әлі де болсын нақты сегментацияны сақтайды, мысалы, ұрпақ миотомдары (бұлшықет бөлімдері) және арқатұс жұйкелері.
Өлшемдер және беттің ауда: көлем арақатынастары
Жоғарғы аудан беиті : көлемге қатысты ағзалар өзінің көлік қажеттіліктерін қамтамасыз еті үшін диффузияға сүйенеді. Оттек, қорек заттары және көміртек диоксиді сияқты ауысым өнімдері ағзаның тіршілік етуін қамтамасыз ету үшін арнайы көлік жүйелерісіз айтарлықтай тез араластырады. Бірақ, диффузия, заттар тасымалын қысқа арақашықтықтарда ғана қамтамасыз ете алады.
Өзіміз көргеніміздей, жазық құрттарда мезодерма тығыз орташа қабықты құрай отырып, эктодерма мен энтодерма арасындағы жапырақта кеңістікті толығымен толтырады. Дене құрылымында екіншілік денеқуыс жоқ болатын болса, онда ацеломикалық деп аталады (2.55 суреті, А).
Аннелидте жоғары бірлескен жануарлар тобында екіншілік қуыс деп аталатын дене қуысы құрылады. Ол мезодермада эмбриондық үрдіс кезінде саңылау сияқты пайда болады. Мұндайдан түзілген қуыс мезодерманы екі жапыраққа бөле отырып, сыртқа қаралғаны – соматикалық, ішке қарағаны – ішкілік (2.55 сурет, Б) деп аталатын целомикалық сұйықтықты толтырады. Эктодермамен байланысатын соматикалық мезодерма дене қабырғасын түзеді. Энтодермамен байланысатын ішкілік мезодерма ішектің бұлшықетті қабырғасын түзеді. Осылайша, екіншілік денеқуыс ішек қабырғасынан дене қабырғасын бөледі.
Мезодерманың екіншілік денеқуыспен төселген негізгі бөлігі бұлшықеттерде дамиды; дене қабырғасын құрайтын бұлшықеттер жануардың локомоциясын қамтамасыз етеді, ал ішек қабырғасының бұлшықеттері соңғысының толқып жиырылғанын қамтамасыз етеді. Дене қабырғасынан ішек қабырғасына дейінгі Дене құрылымының целомикалық типі
Өзіміз көргеніміздей, жазық құрттарда мезодерма тығыз орташа қабықты құрай отырып, эктодерма мен энтодерма арасындағы жапырақта кеңістікті толығымен толтырады. Дене құрылымында екіншілік денеқуыс жоқ болатын болса, онда ацеломикалық деп аталады (2.55 суреті, А).
Екіншілік денеқуыстың биологиялық мәні
- Сондықтан екіншілік денеқуыс ішекті дене қабырғасынан бөлетіні, дене қабырғасының бұлшықеттер қозғалысы, жануарлардың локомоциямен ұқсастықтары, ішек арқылы өнімді итеру және айналдыруға қабілетті ішек қабырғаларына тәуелсіз болуға қабілетті. Нәтижесінде, жануарлар локомоциясының қарқындылығы өседі және әртүрлі функцияларды орындауға қабілетті ішектің әртүрлі бөлшектерінің саралануы басталады. Мысалы, асқазанда өз кезегінде диетаны әртүрлі қылатын мүмкіндік беріп, өнімнің араластырылуы жүреді.
- Екіншілік денеқуыс – сол немесе басқа да ағзалар өсе алатын, дами алатын және бір – біріне тәуелсіз жұмыс істей алатын қуыс.
- Екіншілік денеқуыстың бар болуы дененің үлкендеуіне және оның күрделі құрылымына, яғни көлік пен үйлестік үрдістерімен байланысты қосымша мәселелерді туындатады. Мысалы, өнім ішекте қорытылады, бірақ дене жасушалары ішектің екіншілік денеқуысынан бөлек. Сол сияқты, ішек газ алмасу жүретін дене бетінен кейбір алшақтықта жүреді. Дене қуысының өлшемдерінің үлкеюлеріне қарай 2.8.3 бөлімінде қаралып кеткен мәселелерде көбейеді, нәтижесінде, көлік жүйесінің қажеттілігі туындайды. Сондықтан барлық целомикалық жануарларда түтіктер жүйесінен тұратын тамырлы қан жүретін жүйе пайда болды. Қан – барлық жануарлар денесіне қан жүретін тамырлар қабырғасында орналасатын бұлшықетті талшықтардың қысқару есебінен немесе жүрек жұмысының есебінен таралатын сұйық тін. Тамыр мен жүректе қан қақпақшаларының бар болуы арқасында тек қана бір бағытпен ағады. Өте үлкен күрделі және өте үлкен үйлесімділік керек, демек, үлкен жүйке жүйесі де керек деген сөз. Бұл бағыт эволюциясының бір салдарында цефализация пайда болған, яғни, бастың дамуы (төменде сонымен қатар, дене өлшемдері мен ауданның бетке қатынасына: көлем мән беріңдер).
- Аннелидте екіншілік денеқуыс тағы да қосымша мамандандырылған функция – гидростатикалық функциясын, яғни, сұйық қаңқаны алып жүреді. Қаңқа үш негізгі функцияны орындайды: тірек, қорғаныш және локомоция. Сондықтан целомикалық сұйықтық әрбір сұйықтық сияқты қысылмайды, қысқарған бұлшықет құрт пішінінің өзгерісіне әкелуі мүмкін, бірақ оның көлемін емес. локомоцияда дененің белгілі бір бөліктері уақыт өте келе не ұзын, не кіші, не қысқа, не ірі болады, ол целомикалық сұйықтыққа бұлшықеттің қандай топтары қысым көрсететініне тәуелді болады. Екіншілік қуыстың қорғаныштық рөлі сұйықтықтықтардың барлық бағыт бойынша сыртқы қысымды тең және тез бөлінуіне қабілеттілігімен қамтамасыз етеді.
- Целомикалық сұйықтық қоректік заттар, өнім алмасу және дем алатын көліктерінде қатыса алады, әйтсе де, бұл функцияларды орындаудың негізгі рөлінде қан жүретін тамырлардың жүйесі ойнайды.
Целомикалық жануарлардың дамуындағы тағы бір эволюционды артықшылық болып табылатын метамерия немесе сегментация пайда болды, - жануар денесіндегі көлденең кеңістіктердің бірнеше бірдей бөлшектерге немесе сегменттерге бөлінген ұжым деңгейі. Басқа сөздермен айтқанда, жануар денесі өзіндік бірдей сегментте бір – бірімен орналасқан ұзын қатарлы болады. Сегментация мезодермадан бастау алады, бірақ әдетте мизодермалды және эктодермалды дененің бөліктерін қамтиды.
Анығырақ метамерия бұл бөлініс сырттан жақсы көрінетін (дененің барлық ұзына бойы көшірілуі) сақина тәрізді құрттарда пайда болады (аннелид). Ішкі сегменттер бір бірінен екіншілік денеқуыс арқылы жүретін кеңістіктермен бөлінеді. Әрбір сегментте өзінің өзіндік бұлшықеттер жинағы, қан жүретін тамырлары, жүйке жасушалары, ал кейбіреулерінде репродуктивті ағзалардың топтары болады. Бірақ тіпті анналидтің өзінде сегменттер бір бірінен толығымен тәуелсіз емес, сондықтан жүйкелі, әсіресе қан жүретін жүйелер жануарлар денесінің барлық бойынан өтіп шығуы керек.
Әрбір сегменттің сегментациясы мен құрылымының негізгі жоспары эволюционды бекітілген бойда, әрі қарайғы бөлек сегменттердің немесе олардың кішігірім топтары бойынша, сонымен қатар, тағы бір мүмкіндік енсіз мамандану мен әртүрлі дене бөлшектерінің еңбек бөлінісі сияқты эволюционды өзгерістердің мүмкіндігі пайда болды. Бұл бірнеше жол арқылы болады. Әртүрлі функциялар әртүрлі сегменттермен орындалуы мүмкін; басты қалыптастыру кезінде бірнеше сегменттер қосылғандағы цефализация болғанда, сегменттердің қосылуы да мүмкін болды; бауыраяқтыларда кейбір сегменттердің жоғалуы да байқалады. Бірақ, бунақаяқтылардың ішкі сегментациясы аннелид сияқты анық айтылған. Желілілерде сыртқы сегментациясы құртылған, бірақ кейбір жүйелер әлі де болсын нақты сегментацияны сақтайды, мысалы, ұрпақ миотомдары (бұлшықет бөлімдері) және арқатұс жұйкелері.
Өлшемдер және беттің ауда: көлем арақатынастары
Жоғарғы аудан беиті : көлемге қатысты ағзалар өзінің көлік қажеттіліктерін қамтамасыз еті үшін диффузияға сүйенеді. Оттек, қорек заттары және көміртек диоксиді сияқты ауысым өнімдері ағзаның тіршілік етуін қамтамасыз ету үшін арнайы көлік жүйелерісіз айтарлықтай тез араластырады. Бірақ, диффузия, заттар тасымалын қысқа арақашықтықтарда ғана қамтамасыз ете алады.
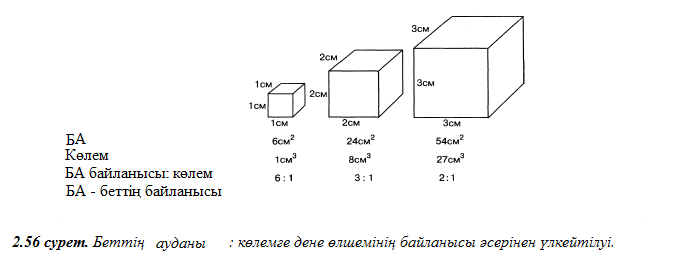
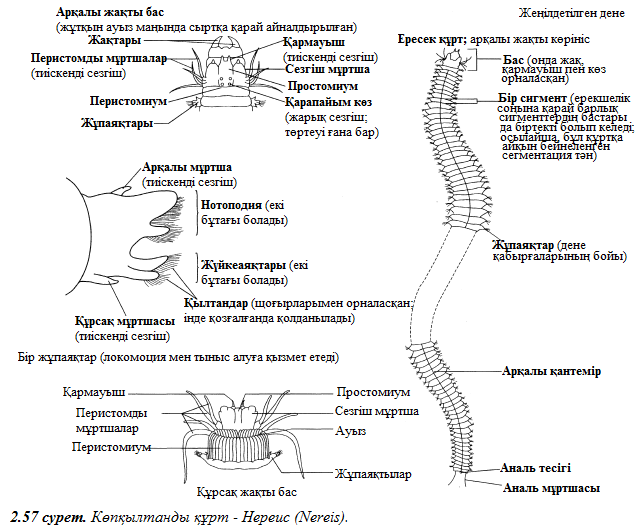
Бұл ретте өлшемдердің кеңейтілу салдарынан қандай да бір дене көлемі оның беткі ауданына қарағанда тезірек кеңейеді және нәтижесінде, беттің ауданы: көлем кішірейеді. Бұны текше мысалындарымен дәлелдеуге болады (2.56 сурет), бірақ сфералық дене, жасуша және барлық тірі ағзалар жағдайларында да сол қағида әсер етеді. Жазық тірі ағзаларда кеңеюде беттің ауданы көлему жазық құрттарғы тән болып, ұзын болып қала береді. Және бұл ағзаларда диффузия толығымен барлық көлік қажеттіліктерін қамтамасыз ете алады. Бірақ, целомикалық жануарларға газалмасу мен көлікті жүйелер үшін мамандандырылған жүйелер қажет.
Polychaeta класы
Құмқұрт (Arenicola) құмда індерді немесе жұмсақ лайлы топырақты бұза отырып, жоғары – төмен зонасында мекен етеді. Нереис (Nereis) (2.54 сурет) не тас астында, не лайлы немесе лайлы – топырақты жерде сукүзенділер қазып эстуарийда мекен етеді.
Nereis сегментациясы сырттан анық көрінеді. ең алдысы мен арттағысын қоспағандағы барлық сегменттер бірдей. Әрбір сегмент бүйірінде жұпаяқтылар деп аталатын өсінділер орналасады. Жұпаяқтылардың жоғарғы тармағы нотоподия, астыңғысы – жүйкеаяқтылар деп аталады (2.57 сурет). Бұл екі құрылымнан (нотоподия мен жүйкеаяқтар) түкшелі қылтандардың екі тобы шығады. Әрбір жұпаяқтыларда қосымша өскіндер – арқалы деп немесе қарын мұртшалары деп аталатын инетәрізді құрылымдар болады. Жұпаяқтылар беті арқылы мол қамтылған қанмен газалмасу жүзеге асырылады. Nereis ескеаяқтар секілді жұпаяқтарын қолданып жорғалайды. Бұл құрт тағы жұпаяқтылар мен бүгіле алатын тері бұлшықетті қабы бұлшықеттерінің келісілген жұмысының арқасында жүзе алады. Nereis денесі кішігірім кутикаламен қоршалған. ас қорытатын арық барлық дене бойына – ауыз тесігінен аналь тесігіне дейін таралады. Nereis жұтқыншағы жылжымалы. Құрт асын жұтқан бойда, жұтқыншақ қайтадан жұтылады.
Nereis – те безендірілген бас болады (2.57 сурет); сондай анық кескінделген цефализация көпқылтанды құрттарға типті, бірақ өзге аналидтерге емес. бас алғашқы және соңғы бөлімдерден тұрады. Алғашқы бөлім, бірінші сегментпен түзілген простомиум, ал соңысы екінші сегментпен түзілген перистомиум. Простомиумда сезгіш қармауыштардың (сыртқы жақта) екі жұбы және екі жұп көздері орналасқан, ал төменгі – бүйір қатысушыларынан сезі сезімімен үлестірілген етті «сезгіш мұртшалар» аулақ кетеді. Ауыз бастың екі бөлімі арасында орналасқан. Оның бірінен – перистомиумнан – ұзын иілгіш мұртшалардың төрт жұбы аулақ кетеді, сезгіш сезімінің үлестірілгендері мен алып жүретіндері, одан басқа, әртүрлі химикалық заттарды қабылдайтын және жануарларға иіс сезі мен дәм сезу ағзалары алмастырушы хеморецепторлар.
Oligochaeta класы
Жауын құрты (Lumbricus terrestris) – 12 – 18 см – ге жетіп қалатын ұзындыққа жететін, ұзын цилиндр денелі жануар (2.58 сурет). Алдыңғының соңғысы конус тәрізді, ал соңғысы сыртқы – құрсақтұс бағытында сопайтылған. Жауынқұрт құрғақтықта тіршілік етеді, ал егер жердегі тіршілік өмірінде тұрса ол барлық мәселелер шешімдерін білмес еді. Құрғақтықтан құтқарыла отыры, ол ылғалды жерде інді бұза отырып, жер бетінде өмір сүреді және тек түнде ғана қорек пен немесе жыныстық серіктестікті табу мақсатында ғана тастап кетеді.
Nereis денесінің сыртқы құрылымы мен жауын құртының айырмашылығы соңғысының жердегі тіршілік өміріне қабілеттілігімен қанағаттандырылады.
Polychaeta класы
Құмқұрт (Arenicola) құмда індерді немесе жұмсақ лайлы топырақты бұза отырып, жоғары – төмен зонасында мекен етеді. Нереис (Nereis) (2.54 сурет) не тас астында, не лайлы немесе лайлы – топырақты жерде сукүзенділер қазып эстуарийда мекен етеді.
Nereis сегментациясы сырттан анық көрінеді. ең алдысы мен арттағысын қоспағандағы барлық сегменттер бірдей. Әрбір сегмент бүйірінде жұпаяқтылар деп аталатын өсінділер орналасады. Жұпаяқтылардың жоғарғы тармағы нотоподия, астыңғысы – жүйкеаяқтылар деп аталады (2.57 сурет). Бұл екі құрылымнан (нотоподия мен жүйкеаяқтар) түкшелі қылтандардың екі тобы шығады. Әрбір жұпаяқтыларда қосымша өскіндер – арқалы деп немесе қарын мұртшалары деп аталатын инетәрізді құрылымдар болады. Жұпаяқтылар беті арқылы мол қамтылған қанмен газалмасу жүзеге асырылады. Nereis ескеаяқтар секілді жұпаяқтарын қолданып жорғалайды. Бұл құрт тағы жұпаяқтылар мен бүгіле алатын тері бұлшықетті қабы бұлшықеттерінің келісілген жұмысының арқасында жүзе алады. Nereis денесі кішігірім кутикаламен қоршалған. ас қорытатын арық барлық дене бойына – ауыз тесігінен аналь тесігіне дейін таралады. Nereis жұтқыншағы жылжымалы. Құрт асын жұтқан бойда, жұтқыншақ қайтадан жұтылады.
Nereis – те безендірілген бас болады (2.57 сурет); сондай анық кескінделген цефализация көпқылтанды құрттарға типті, бірақ өзге аналидтерге емес. бас алғашқы және соңғы бөлімдерден тұрады. Алғашқы бөлім, бірінші сегментпен түзілген простомиум, ал соңысы екінші сегментпен түзілген перистомиум. Простомиумда сезгіш қармауыштардың (сыртқы жақта) екі жұбы және екі жұп көздері орналасқан, ал төменгі – бүйір қатысушыларынан сезі сезімімен үлестірілген етті «сезгіш мұртшалар» аулақ кетеді. Ауыз бастың екі бөлімі арасында орналасқан. Оның бірінен – перистомиумнан – ұзын иілгіш мұртшалардың төрт жұбы аулақ кетеді, сезгіш сезімінің үлестірілгендері мен алып жүретіндері, одан басқа, әртүрлі химикалық заттарды қабылдайтын және жануарларға иіс сезі мен дәм сезу ағзалары алмастырушы хеморецепторлар.
Oligochaeta класы
Жауын құрты (Lumbricus terrestris) – 12 – 18 см – ге жетіп қалатын ұзындыққа жететін, ұзын цилиндр денелі жануар (2.58 сурет). Алдыңғының соңғысы конус тәрізді, ал соңғысы сыртқы – құрсақтұс бағытында сопайтылған. Жауынқұрт құрғақтықта тіршілік етеді, ал егер жердегі тіршілік өмірінде тұрса ол барлық мәселелер шешімдерін білмес еді. Құрғақтықтан құтқарыла отыры, ол ылғалды жерде інді бұза отырып, жер бетінде өмір сүреді және тек түнде ғана қорек пен немесе жыныстық серіктестікті табу мақсатында ғана тастап кетеді.
Nereis денесінің сыртқы құрылымы мен жауын құртының айырмашылығы соңғысының жердегі тіршілік өміріне қабілеттілігімен қанағаттандырылады.
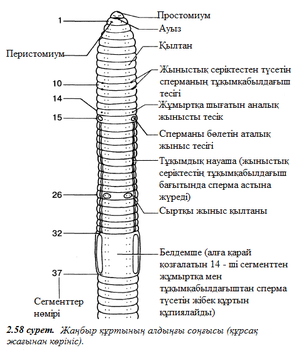
|
Жауын құрт денесі ағынды пішін болады және жердегі еркін қозғалатын құрттарға қарсы шыға алатын қандай да бір өскіндер құртылып кеткен. Ауыз астында орналасқан простомиум кішігірім және дөңгелек, онда сезгіш өсінділер жоқ. Алғашқы және соңғы сегменттерінен басқа, барлық сегменттерде қылттанның төрт жұбы болады; оның екеуі құрсақтұс жақта (құрсақ жақта), ал екеуі сыртта орналасқан. Қылтандар қылтанды қап денесінде орналасқаннан шығады. Олар ішке кіре алады және сыртқа арнайы бұлшықеттер әрекетінен шығарылады. Қылтанда құрттардың жердегі қозғалыстарын жеңілдетеді. Қылтандар тағы қорғаныш үшін қолданылады: олардың ін қабырғасына тағыныла отырып, ол онда берік ұсталады. сегментте орналасқан ең ұзын қылтандар 10 – 15, 26 және 32 – 37, жыныстық қатынаста қолданылады.
|
Жыныстық қатынаста қолданылатын тағы бір құрылым, 32 – 37 сегменттерде орналасқан – белдемше. Белдемше эпидермисі арқалы және қарын беттерінде кейбір жуандатылуы ет – тоқымға ұқсап кететін безді жасушалардың басым көпшілігін ұстайды. Белдемше жыныстық қатынас және жібек құрт қалыптастыру үрдістеріне қатысады.
Құрттардың ауыз және аналь тесігі дененің қарама – қарсы аяқ - қолдарында орналасқан. Жауынқұрт құрт топырақпен бірге сіңіретін дитритпен (органикалық материалды іріп – шірітітен фрагменттер) қоректенеді. Сіңірілген топырақтың басым бөлігі ішек арқылы келеді және жер бетіне ерекше ұоғырлармен лақтырылады.
Целомикалық сұйықтықтың арқалы поралары арқылы бөлінетін және эпидермистің безді жасушаларын құпиялайтын шырыш үнемі кіші кутикуланы ылғалдатады. Нәтижесінде, кутикула құрғатылуына жол берілмейді, газ алмасу жеңілдетіледі және өз тұқымынан құрттардың жерде қозғалу қозғалысы жеңілдететін жағармайды шығарады. Дәл кутикула арқылы газ алмасу тармақталған капиллярлар байланыстарының эпидермалды қабатиың бар болуып келуіне тәуелдіүрді диффузия жолымен болады.
Үш алғашқы және бір соңғыдан басқа, барлық сегменттер осморегуляция мен бөлініп шығу функцияларын орныдайтын нефрид арықтары жұр – жұптан болады. Ол дене бетінде жұп болып ашылады.
Жауын құрттың репродуктивті жүйесі, көбею тәртібі сияқты, жердегі тіршілік өмірімен және гаметалар мен ұрықтанған жұмырқаларды құрғап қалудан сақтануы керектігімен түсіндірілетін үлкен күрделілікпен ажыратылады. Lumbricus қосжынысты (яғни,ірбір дарада аталық пен аналық репродуктивті ағзалар болады). Жауынқұрттар бір – бірімен байланысы өте сирек болады, бірақ, бұл мұндай болған жағдайда, екі түрлі даралар шағылыстырыла алады, сондықтан екеуі де қос жыныстылар. Нәтижесінде, сперамалардың өзара алмасуы жүреді және екі құрт та ұрықтанады.
Жауын құрттың жыныс ағзалары дененің алдыңғы соңында шоғырланады. Жыныстық көбеюмен байланысты сыртқы белгілері 2.58 суретте көрсетілген. Шағылыстырылу мен соңғы ұрықтанған жұмыртқалардың алып қойылуы – өте күрделі үрдіс. Шағылыстырылған даралар өзара қарама – қарсы бағыттарда бір – бірінің қасына орналасады, сондықтан серіктестің буреуінің басы басқасының құйрық бөлігіне бағытталғаны көрінеді және керісінше.
2.8.6. Arthropoda типі (буын аяқтылар)
Буын аяқтылардың систематикасы мен ерекше белгілері 2.17 кестесінде жинақталған. Arthropoda типі саны бойынша өзгелердің арасында ең көпсандысы. Барлық белгі түрлердің жалпы санының төртінен үштен астамы – осы топтың өкілдері. Тек жәндіктердің біреуінің үлесіне барлық белгілі түрлердің жартысынан көбі келеді. Буынаяқтылар құрғақтықтағы мен жердегі барлық мекер ортасын игеріп алды.
Буынаяқтылар дене құрылысының негізгі жоспары шектен тыс сәтті болып байқалды, бейімделме радиция деп аталатын үрдіс нәтижесі белгілі бір сәтті арғы тектің эволюция жолымен бірте – бірте дамыған пішіндерінен әртүрлі экологиялық тауашалардың көбісімен толтырылатын жан – жақты түрлер пайда болды (26 бөлім). Мысалы, жәндіктерде, бейімделме радияциялардың нәтижесінде ұшқыштыққа, індегі тіршілікке, суға, паразитікке қабілетті түрлер пайда болды.
Жәндіктер денесінің құрылым жоспарын сегменттік аннелид денесінің эволюция жолымен бірте – бірте дамитын құрылым жоспары ретінде қарауға болады. Бұл мысалда, метамерлі сегментацияны қалай қолданылуға болатыны анық көрсетулі.бұрынғы буынаяқтыларла дененің ұзына бойына газалмасу, қорек табу, локомоция және әртүрлі белгілерді тану сияқты маңызды функцияларды орындайтын қарапайым аяқ – қолдар орын алған болатын. Қазіргі буынаяқтыларда аннелидпен салыстырып қарағанда үрдіс кішілеу болып келеді, мамандану одан да күрделі және бөліну еңбегінің көрсетілгеніндей одан да көп мамандандырылған аяқ – қолдарды әкелді. Сегментацияның сыртқы құрылымы әлі де болсын бақыланады, бірақ сегменттер саны аннелидке қарағанда кішірейе береді.
Біз төменде басқа да буынаяқтылардың негізгі ерекшеліктерін қарастырамыз. Олар сегментацияның жоғарыда айтылған эволюциясы байланысында олардың өркендеуін жақсартады.
ЭКЗОҚАҢҚА НЕМЕСЕ КУТИКУЛА. Кутикула эпидермис жасушаларымен құпияланады. Кутикула құрамына өсімдіктердің жасушалық қабырғасына тірек материалы болып қызмет ететін, целлюлозаға аумайтын азотты полисахарид – хитин жатады. Хитин жарылыста жоғарғы беріктілігімен ажыратылады (оны екі соңымен жырту қиын). Хитиннің басқа да химиялық байланыстармен байланысуы экзоқаңқа сипатына өзгеріс әкелуі мүмкін. Мысалы, минералды тұздар қоссақ (әсіресе кальций тұздары), экзоқаңқа шаянтәрізділерге қарағанда беруктеу болуы мүмкін. Мұндай әсер ақуызда да болады. бұл экзоқаңқалар үшін беріктік, иілгіштік және қаттылық жағынанүлкен жан – жақтылықты ашады. Кутикула иілгіштігі буындарда маңызды рөл ойнайды. Экзоқаңқаның бар болуы келесі артықшылықтарды құрайды:
Құрттардың ауыз және аналь тесігі дененің қарама – қарсы аяқ - қолдарында орналасқан. Жауынқұрт құрт топырақпен бірге сіңіретін дитритпен (органикалық материалды іріп – шірітітен фрагменттер) қоректенеді. Сіңірілген топырақтың басым бөлігі ішек арқылы келеді және жер бетіне ерекше ұоғырлармен лақтырылады.
Целомикалық сұйықтықтың арқалы поралары арқылы бөлінетін және эпидермистің безді жасушаларын құпиялайтын шырыш үнемі кіші кутикуланы ылғалдатады. Нәтижесінде, кутикула құрғатылуына жол берілмейді, газ алмасу жеңілдетіледі және өз тұқымынан құрттардың жерде қозғалу қозғалысы жеңілдететін жағармайды шығарады. Дәл кутикула арқылы газ алмасу тармақталған капиллярлар байланыстарының эпидермалды қабатиың бар болуып келуіне тәуелдіүрді диффузия жолымен болады.
Үш алғашқы және бір соңғыдан басқа, барлық сегменттер осморегуляция мен бөлініп шығу функцияларын орныдайтын нефрид арықтары жұр – жұптан болады. Ол дене бетінде жұп болып ашылады.
Жауын құрттың репродуктивті жүйесі, көбею тәртібі сияқты, жердегі тіршілік өмірімен және гаметалар мен ұрықтанған жұмырқаларды құрғап қалудан сақтануы керектігімен түсіндірілетін үлкен күрделілікпен ажыратылады. Lumbricus қосжынысты (яғни,ірбір дарада аталық пен аналық репродуктивті ағзалар болады). Жауынқұрттар бір – бірімен байланысы өте сирек болады, бірақ, бұл мұндай болған жағдайда, екі түрлі даралар шағылыстырыла алады, сондықтан екеуі де қос жыныстылар. Нәтижесінде, сперамалардың өзара алмасуы жүреді және екі құрт та ұрықтанады.
Жауын құрттың жыныс ағзалары дененің алдыңғы соңында шоғырланады. Жыныстық көбеюмен байланысты сыртқы белгілері 2.58 суретте көрсетілген. Шағылыстырылу мен соңғы ұрықтанған жұмыртқалардың алып қойылуы – өте күрделі үрдіс. Шағылыстырылған даралар өзара қарама – қарсы бағыттарда бір – бірінің қасына орналасады, сондықтан серіктестің буреуінің басы басқасының құйрық бөлігіне бағытталғаны көрінеді және керісінше.
2.8.6. Arthropoda типі (буын аяқтылар)
Буын аяқтылардың систематикасы мен ерекше белгілері 2.17 кестесінде жинақталған. Arthropoda типі саны бойынша өзгелердің арасында ең көпсандысы. Барлық белгі түрлердің жалпы санының төртінен үштен астамы – осы топтың өкілдері. Тек жәндіктердің біреуінің үлесіне барлық белгілі түрлердің жартысынан көбі келеді. Буынаяқтылар құрғақтықтағы мен жердегі барлық мекер ортасын игеріп алды.
Буынаяқтылар дене құрылысының негізгі жоспары шектен тыс сәтті болып байқалды, бейімделме радиция деп аталатын үрдіс нәтижесі белгілі бір сәтті арғы тектің эволюция жолымен бірте – бірте дамыған пішіндерінен әртүрлі экологиялық тауашалардың көбісімен толтырылатын жан – жақты түрлер пайда болды (26 бөлім). Мысалы, жәндіктерде, бейімделме радияциялардың нәтижесінде ұшқыштыққа, індегі тіршілікке, суға, паразитікке қабілетті түрлер пайда болды.
Жәндіктер денесінің құрылым жоспарын сегменттік аннелид денесінің эволюция жолымен бірте – бірте дамитын құрылым жоспары ретінде қарауға болады. Бұл мысалда, метамерлі сегментацияны қалай қолданылуға болатыны анық көрсетулі.бұрынғы буынаяқтыларла дененің ұзына бойына газалмасу, қорек табу, локомоция және әртүрлі белгілерді тану сияқты маңызды функцияларды орындайтын қарапайым аяқ – қолдар орын алған болатын. Қазіргі буынаяқтыларда аннелидпен салыстырып қарағанда үрдіс кішілеу болып келеді, мамандану одан да күрделі және бөліну еңбегінің көрсетілгеніндей одан да көп мамандандырылған аяқ – қолдарды әкелді. Сегментацияның сыртқы құрылымы әлі де болсын бақыланады, бірақ сегменттер саны аннелидке қарағанда кішірейе береді.
Біз төменде басқа да буынаяқтылардың негізгі ерекшеліктерін қарастырамыз. Олар сегментацияның жоғарыда айтылған эволюциясы байланысында олардың өркендеуін жақсартады.
ЭКЗОҚАҢҚА НЕМЕСЕ КУТИКУЛА. Кутикула эпидермис жасушаларымен құпияланады. Кутикула құрамына өсімдіктердің жасушалық қабырғасына тірек материалы болып қызмет ететін, целлюлозаға аумайтын азотты полисахарид – хитин жатады. Хитин жарылыста жоғарғы беріктілігімен ажыратылады (оны екі соңымен жырту қиын). Хитиннің басқа да химиялық байланыстармен байланысуы экзоқаңқа сипатына өзгеріс әкелуі мүмкін. Мысалы, минералды тұздар қоссақ (әсіресе кальций тұздары), экзоқаңқа шаянтәрізділерге қарағанда беруктеу болуы мүмкін. Мұндай әсер ақуызда да болады. бұл экзоқаңқалар үшін беріктік, иілгіштік және қаттылық жағынанүлкен жан – жақтылықты ашады. Кутикула иілгіштігі буындарда маңызды рөл ойнайды. Экзоқаңқаның бар болуы келесі артықшылықтарды құрайды:
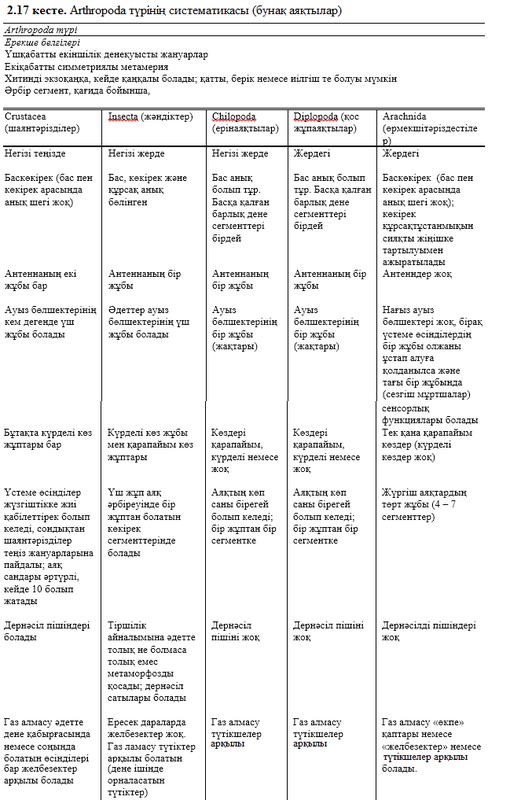

ҚАНДЫҚУЫС. Буынаяқтылар мен моллюскаларда даму кезінде екіншілік денеқуыс толығымен дерлік қандықуыс деп аталатын басқа бір қуысқа айналады (2.60сурет). Қандықуыс тамырлы қан жүретін жүйе қуысынан дамиды және сондықтан қанға толыққан. Ағзаның әрбір бөлігі тікелей қанмен малынады. Екіншілік денеқуыс болады, бірақ ол өте аз және репродуктивті жолдар мен бөлініп шығушы ағзалар қуытармен шектелген. Қан көлемінің буын аяқтылардың дене көлеміне деген үлкен байланысы өзінің кезегінде бұл жануарлардың ең үлкен белсенділігін қамтамасыз ету үшін оларға заттар алмасуының үлкен дәрежесін қолдауға ерік беріледі. Бірақ зақымдануда қан жоғалту өте жоғарғы қауіпке әкеледі.
ДЕНЕ БӨЛШЕКТЕРІНІҢ МАМАНДАНУЫ. Буынаяқтыларда айтылған еңбек бөлінісі айтарлықтай мықтырақ, ол анық дайын болған дене бөлшектеріне жәрдем берді, анығырақ айтқанда, бас және көп жағдайда көкірек пен құйрыққа. Баста көз және антенналар сияқты сенсорлы рецепторлар орналасқан, сонымен қатар, ас табуға көмектесетін үстеме өсінділер. Жануарларда екіжақты симметриялы алдыңғы соңы (бас) дененің барлық бөлшектерінен алғашқы болып, жаңа ортамен байланысқа келеді.
- Құрғақа тірек болып қызмет етеді;
- Экзоқаңқаның ішкі бетіне бұлшықеттер бекінеді, атап айтқанда, локомоцияға қатысатындар, мысалы ұшу;
- Физикалық қауіптерден қорғаныш болып қызмет етеді;
- Кутикуланы арнайы безбен істеп шығарылатын балауызды қабатпен жабушы жердеге мекен ет орталарын құрғап қалудан сақтайды;
- Жәндіктердің ұшуға қабілеттілігі, бүрге мен шегірткені ң секіргені сияқты экзоқаңқада өте иілгіш нәруыздың бар болуына тәуелді болады;
- Экзоқаңқа ұшқын жануарларға өте маңызды болып келетін төмен тығыздығы болады;
- Кутикулалардың бар болуы сегменттердің арасында иілгіш буындардың туындатуына мүмкіндік тудырады;
- Экзоқаңқа тістей латын, уақтайтын, соратын (2.61 суреті, Б) немесе өнімді ұрықтататын қатты иықтарды түзе отырып, түр өзгерте алады.
- Экзоқақңқа орындары, көзде жарықтың пайда болуын және суда бүркену мүмкіндігін қамтамасыз ететін әйнекті жерлер болып келеді.
- Дененің соңғы өлшемдері шектеулі, сондықтан, өткен бөлімде айтқандай, дене өлшемінің үлкейтілуіне қарай беттің ауданы : көлем қатынасы төмен түседі. Экзоқаңқа өлшемдері беттің ауданына тәуелді болады, сонда жануар массасы ретінде көлеміне тәуелді болады. Пілмен буынаяқтылар өлшемі не өзіндік массасын ұстай алмады, не болмаса экзоқаңқа жануар орнынан қозғалып кетпес үшін соншалықты массивті болуы керек еді. (Жәндіктер өлшемдеріне жатқызылатын өзге бір маңызды шектеу, негізінде оттек диффузиясының жолымен трахея деп аталатын арнайы түтікшелер арқылы жүзеге асатын тыныс алумен байланысты. Қазіргі жәндіктерден ең ірілері – ұзындығы 30 см – ге жеттеін талшықтылар, ал қоңыздардың геркулес қоңызы сияқты ең ірілерінде массасы 100 г – ға жетеді, яғни тышқан массасы).
- Экзоқаңқа жануарлар дамуын шектейді. Мұның мәселесі периодтық түлеулермен (экдизистер) иемделе алады. Бірақ, кутикула беріктенбегенше, жануар түгелімен жыртқыштарға осалды болып келеді, сондықтан түлеулер біткенше жәндіктер әдетте паналарға жасырынылады.
ҚАНДЫҚУЫС. Буынаяқтылар мен моллюскаларда даму кезінде екіншілік денеқуыс толығымен дерлік қандықуыс деп аталатын басқа бір қуысқа айналады (2.60сурет). Қандықуыс тамырлы қан жүретін жүйе қуысынан дамиды және сондықтан қанға толыққан. Ағзаның әрбір бөлігі тікелей қанмен малынады. Екіншілік денеқуыс болады, бірақ ол өте аз және репродуктивті жолдар мен бөлініп шығушы ағзалар қуытармен шектелген. Қан көлемінің буын аяқтылардың дене көлеміне деген үлкен байланысы өзінің кезегінде бұл жануарлардың ең үлкен белсенділігін қамтамасыз ету үшін оларға заттар алмасуының үлкен дәрежесін қолдауға ерік беріледі. Бірақ зақымдануда қан жоғалту өте жоғарғы қауіпке әкеледі.
ДЕНЕ БӨЛШЕКТЕРІНІҢ МАМАНДАНУЫ. Буынаяқтыларда айтылған еңбек бөлінісі айтарлықтай мықтырақ, ол анық дайын болған дене бөлшектеріне жәрдем берді, анығырақ айтқанда, бас және көп жағдайда көкірек пен құйрыққа. Баста көз және антенналар сияқты сенсорлы рецепторлар орналасқан, сонымен қатар, ас табуға көмектесетін үстеме өсінділер. Жануарларда екіжақты симметриялы алдыңғы соңы (бас) дененің барлық бөлшектерінен алғашқы болып, жаңа ортамен байланысқа келеді.
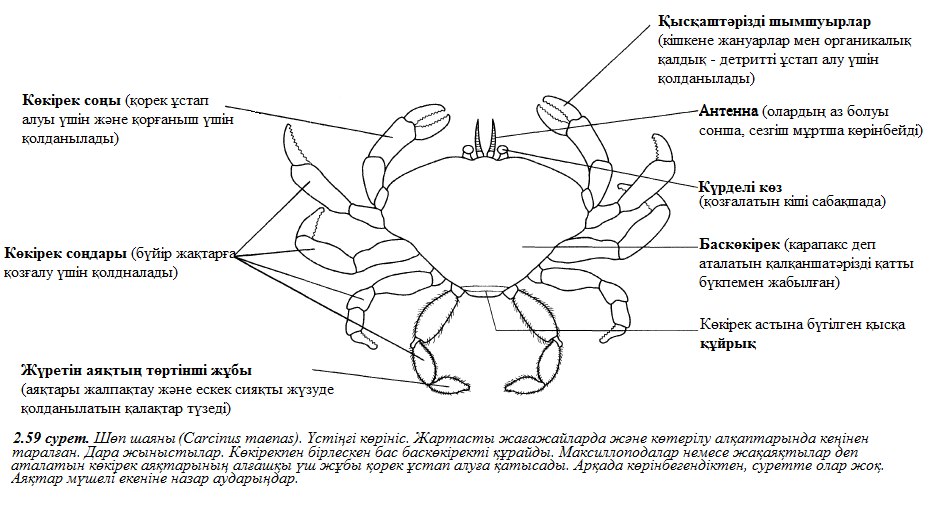
Дәл осы себеппен алдыңғы соңы мамандануға шыдаған, буынаяқтыларда ми аннелидке қарағанда айтарлықтай ірілеу және цефализация да айтарлықтай мықтырақ деп айтылған.
ҰШУ. Жәндіктер эволюция кезінде ұшуға қабілеттіктері болған, және олардың тамақ табуға және дұшпандарынан қашуға мүмкідік туды (2.61 сурет).
Жәндіктердің тіршілік кезеңдері. Жәндіктердің тіршілік кезеңдері өте жан – жақты және жиі орасан күрделі болып келеді. Көп жәндіктерге метаморфозбен даму тән (грек. metamόrphōsis - айналдыру). Метаморфоз – оның тіршілік кезеңінің бойынан пайда болатын жануарлардың құрылымы мен пішіндерінің терең өзгерістері. Дернәсіл кезеңдерінің жәндіктердің қарапайымдылау топтары ересек жәндіктерге (имаго) жиі ұқсайды. Қағида бойынша, әрбір соңғы кезең нимфа немесе жас) ересек жәндікке ұқсай бастайды. Дамудың мұндай типі бірте – бірте немесе толық емес метаморфоз деп аталады. Жәндіктің тіршілік кезеңінің мысалы болып шегіртке типі қызмет етеді.
ҰШУ. Жәндіктер эволюция кезінде ұшуға қабілеттіктері болған, және олардың тамақ табуға және дұшпандарынан қашуға мүмкідік туды (2.61 сурет).
Жәндіктердің тіршілік кезеңдері. Жәндіктердің тіршілік кезеңдері өте жан – жақты және жиі орасан күрделі болып келеді. Көп жәндіктерге метаморфозбен даму тән (грек. metamόrphōsis - айналдыру). Метаморфоз – оның тіршілік кезеңінің бойынан пайда болатын жануарлардың құрылымы мен пішіндерінің терең өзгерістері. Дернәсіл кезеңдерінің жәндіктердің қарапайымдылау топтары ересек жәндіктерге (имаго) жиі ұқсайды. Қағида бойынша, әрбір соңғы кезең нимфа немесе жас) ересек жәндікке ұқсай бастайды. Дамудың мұндай типі бірте – бірте немесе толық емес метаморфоз деп аталады. Жәндіктің тіршілік кезеңінің мысалы болып шегіртке типі қызмет етеді.
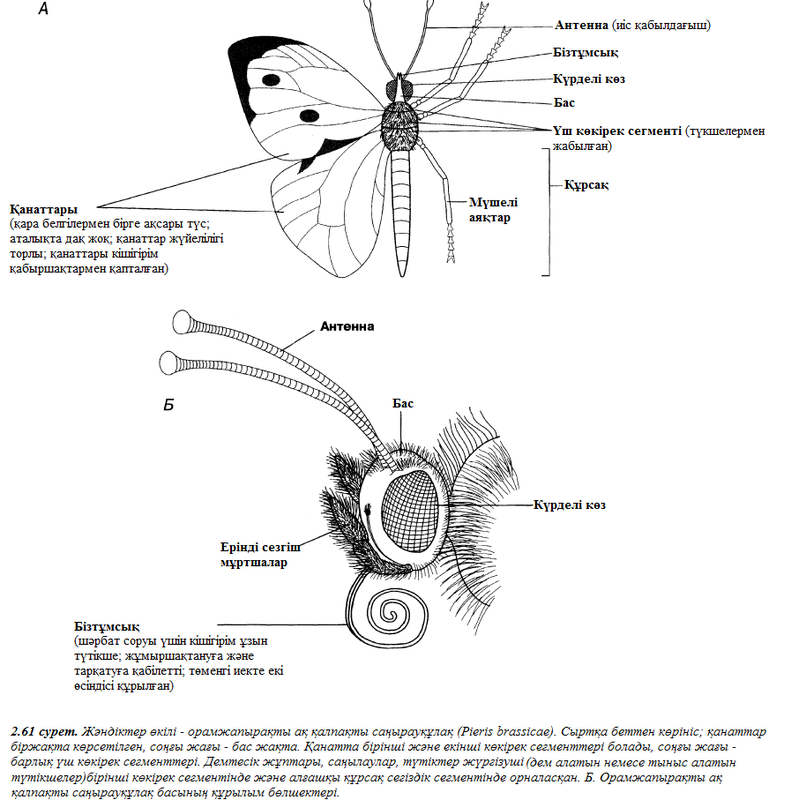
Кешірек эволюция кезеңдеріндегі пайда болған топтарда дернәсіл кезеңдері ересек жәндіктерден бірден айырылады. Соңғы түлеу ішінде метаморфоздың дернәсіл тіндерін бұзатын компоненттер есебінен ересек тіндердің түзілуімен жүретін қуыршақтың оларда түзілуіне әкеледі. Айналымның мұндай типі голометаболиялық немесе толық метаморфоз деп аталады. Мұндай типтің тіршілік кезеңінің мысалы 2.62 суретте келтірілген. Балауса немесе ересек пішіндер метаморфоздың арқасында әртүрлі жерлерде тіршілік етуге және әртүрлі өнім ресурстарын қолдануға, яғни бұл формалар арасындағы жыныстық қатынасты түсіру үшін экологиялық қуыстарда орын алуға мүмкіндік береді. Мысалы, инелік нимфалары суда тіршілік етеді, су жәндіктерімен қоректенеді, ал оларда газалмасу желбезек арқылы жүзеге асады, она ересек инеліктер сияқты ауа ортасында мекендейтіндер жердегі жәндіктермен қоректенеді, ал оларда газ алмасу түтікшелер арқылы болады. Сол сияқты, қабыраққанаттылардың (түңгі және күндізгі көбелектер) жұлдызқұрттарында негізінде жапырақтармен қоректенеді және олардың ауыз бөліктері таласушы типтен болады, ересек көбелектер шәрбатпен қоректенеді және олардың ауыз бөлшектері емуші типтен болады. Егер жәндіктерде толығымен қанат болатын болса, оларда түлеу мүмкін емесе болады да, олар өсуді тоқтатады.
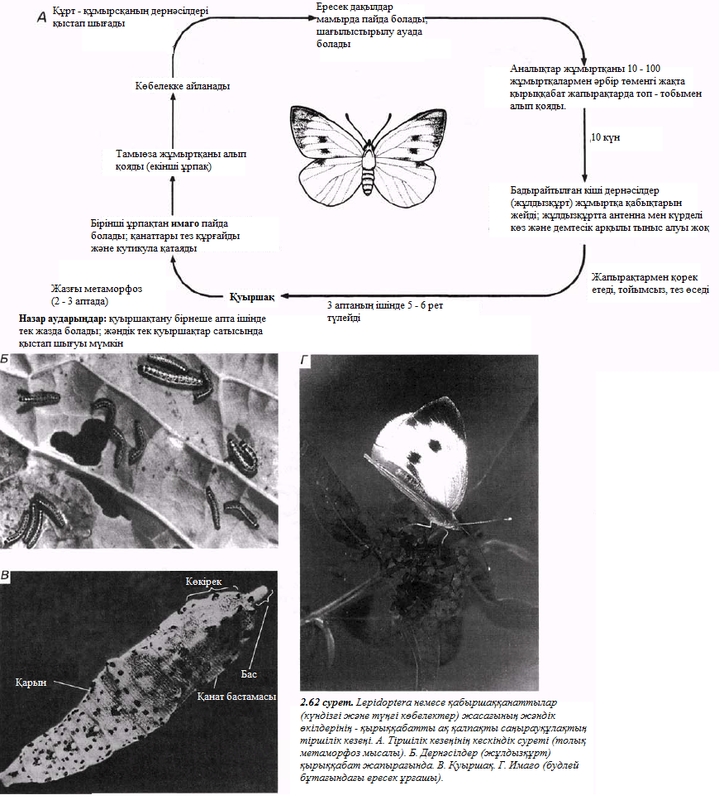
2.8.7. Mollusca типі (моллюсалар)Mollusca типінің систематикасы мн ерекше белгілері 2.18 кестесінде (берілген заттың 6 класынан 3 – уі ғана қаралған) жинақталған. Бұл тип ағзаның әртүрлі топтарын байланыстырады, жай жорғалағыш ұлу мен шырышты ұлу, отырықшы жармаларға біркелкі болып келетін жеуге болатын розиньканың теңіз моллюскілері, венерка және т.б. – рын қосып айтқанда және өте қозғалмалы басаяқтылар (2.63 сурет).
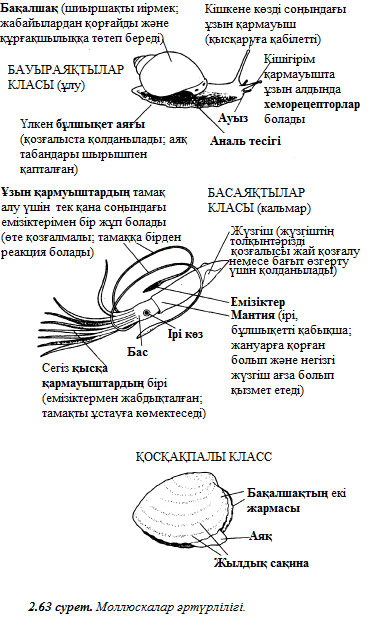
ұл тип қазіргі 80 000 тіршілік етушілерде тұрады және шамамен 35 000 – ы қазынды түрлер; моллюкалар барлық саны бойынша тек қана бауыраятыларға беріле отырып, екінші орынды иемденеді. Моллюсканың бір түрі – гигант кальмары – массасы бірнеше тонға бағаланатын, үзындығы 16 м болатын еі ірі омыртқасыз болып табылады.
Қорғаныш бақалшағыры құрылуы және желбезектердің қолданылуы немесе газалмасу үшін оңайлары моллюскаларға судағылар мен жердегі мекендер сияқты игеруге және ең әртүрлі экологиялық қуыстарға орын ауыстыруға иелік берді. Бірақ бақалшақ қозғалыста бөгет болуы мүмкін және қозғалмалы моллюскалырнда бақалшақ не редукцияланған, не болмаса мүлдем жойылп кеткен. |
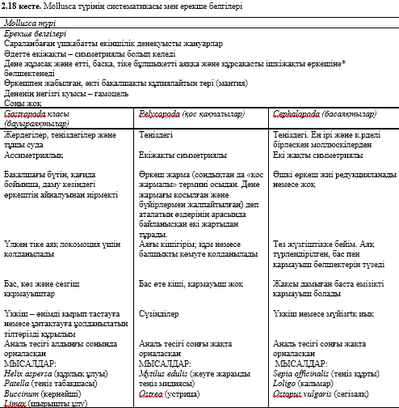

|
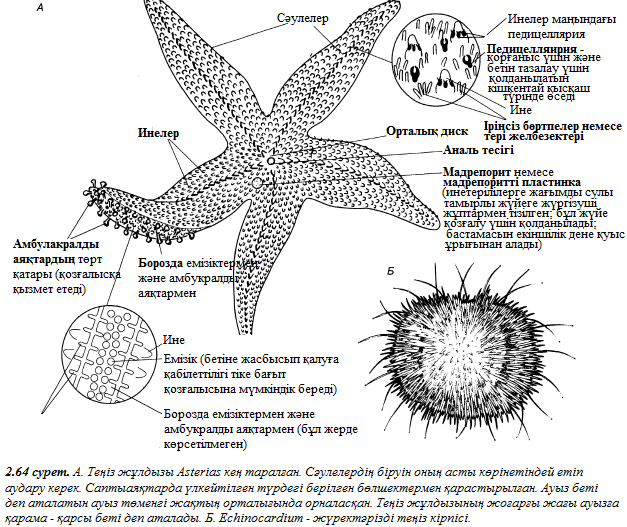
- Chordata типі (хордалылар)
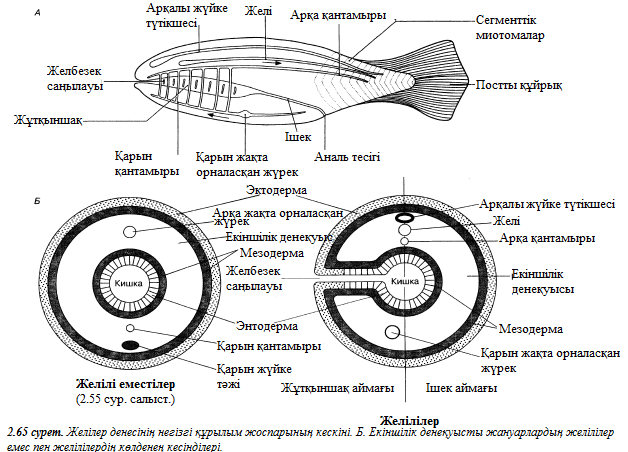
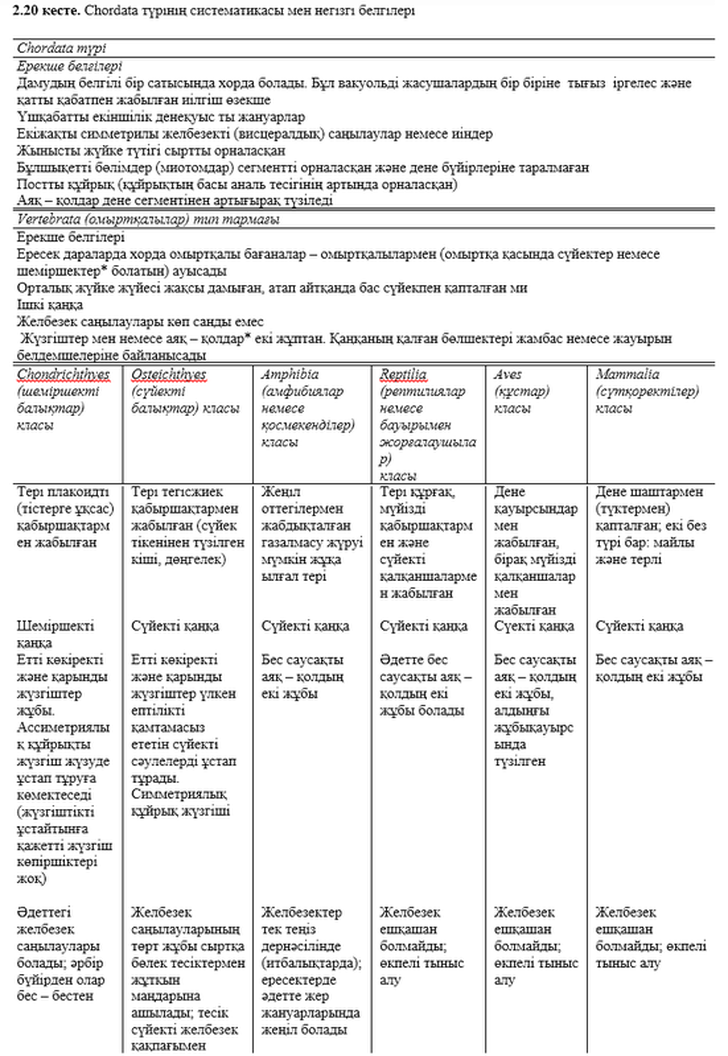
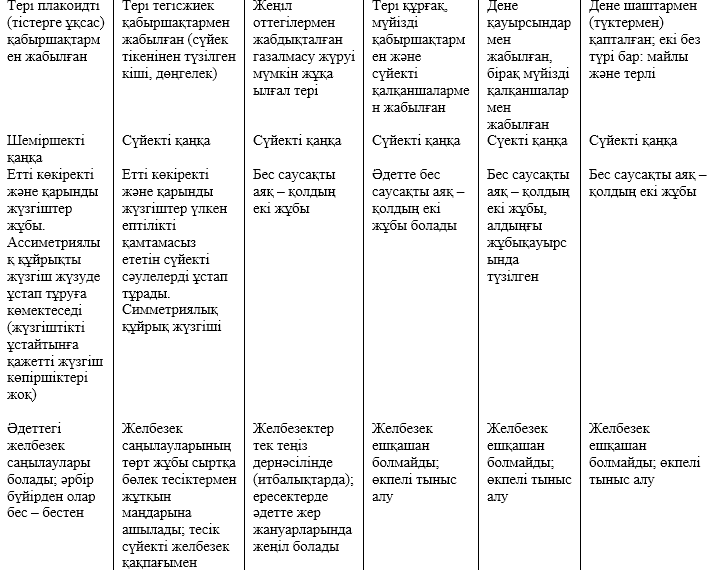
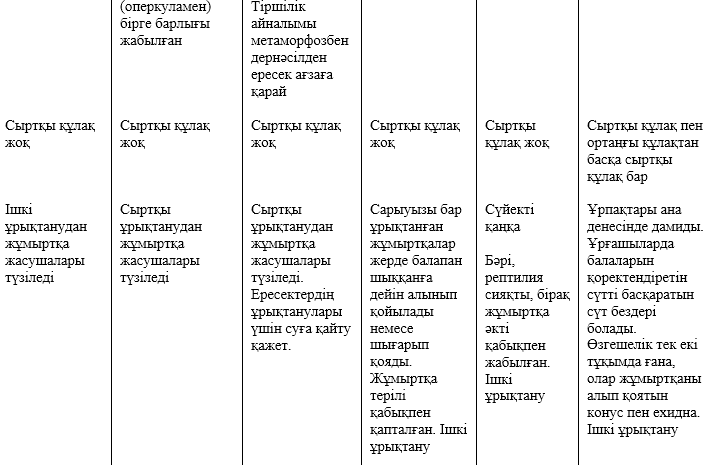
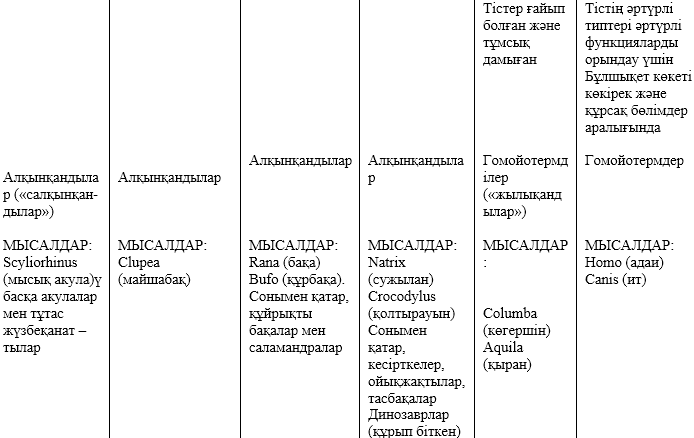

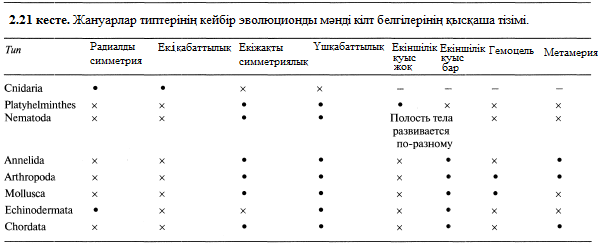
Хордалылардың дене құрылысының негізгі жоспары 2.65, А және Б суреттерінде берілген, ал омыртқалылардың өкілділіктер мысалы – 2.66 суретте.
2.21 кестесінде әртүрлі жануарлардың эволюционды маңызды кілт белгілері берілген.
2.21 кестесінде әртүрлі жануарлардың эволюционды маңызды кілт белгілері берілген.
9 тарау
Энергияның қолданылуы
7 – ші бөлімде біз энергия дегеніміз не және несімен тірі ағзаларға маңызды сұрақтарға келдік. Келтірілген мысалдар бойынша,энергияның түрлі формада кездесетіндігін аңғаруға болады. Олар: химиялық, электрикалық, механикалық, жарық және жанармай түрінде. Оқырман тірі ағзаларда осы түрлі энергия формулаларын қалай қолданылатынын үйнеру керек әрі мысал келтіріп білу керек.(егер ол келтіре алмайтын болса, онда 7.1 бөлімге қайта оралуына тура келеді).
Энергияға байланысты сұрақтар – биологияда маңызды сұрақтардың бірі болып келеді. Жасушадан бастап экожүйеге дейінге барлық тірі жүйелер өз қызметін атқару үшін энергияны қажет етеді. Егер кез – келген жасушаны энергия көздерінен тыс қалдырсақ өлімге ұшырайтының байқаймыз мұндай көріністі аэробты жасушаларға цианидпен әсер еткенде көруге болады, осылай кез – келген экожүйені күн энергиясынан тысқары қалдырсақ өлетінін көреміз.
9. 1. Жануарлардың жарық энергиямен байланысын түсіндіріндер.
Энергияға байланысты сұрақтар – биологияда маңызды сұрақтардың бірі болып келеді. Жасушадан бастап экожүйеге дейінге барлық тірі жүйелер өз қызметін атқару үшін энергияны қажет етеді. Егер кез – келген жасушаны энергия көздерінен тыс қалдырсақ өлімге ұшырайтының байқаймыз мұндай көріністі аэробты жасушаларға цианидпен әсер еткенде көруге болады, осылай кез – келген экожүйені күн энергиясынан тысқары қалдырсақ өлетінін көреміз.
9. 1. Жануарлардың жарық энергиямен байланысын түсіндіріндер.
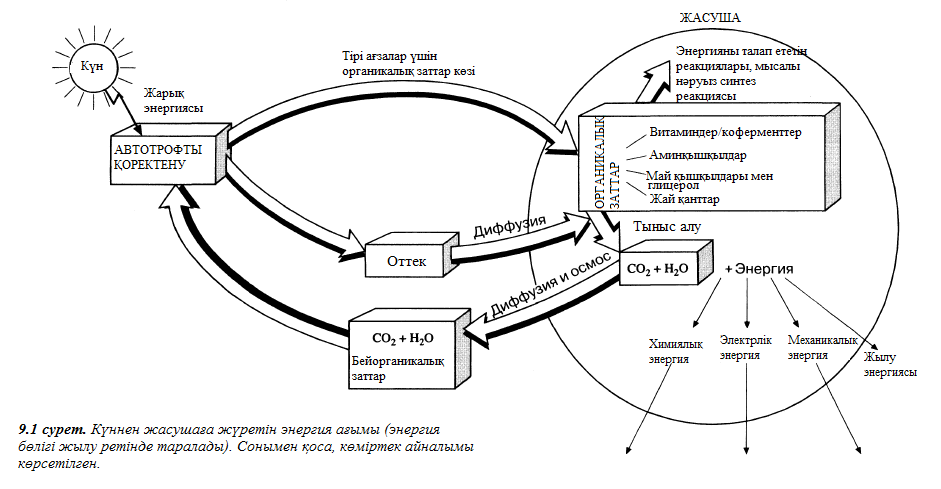
Қоректену – тірі ағзаға қажетті энергиямен қамтамасыз ететін үрдіс. Қоректен ағза жасуша құрылымынаң құрастырылуына және репарацияға керекті материаларды алады. Алайда энергия тағам құрамында жасушаны қолдана алатындай қабілетте, яғни белгілі – бір формада болу керек. Бұл функцияны тыныс алу атқара алады, бұл үрдіске осы бөлімді қарастыруға негізделіп отыр. Автотрофты қоректену, гетеротрофты қоректену және тыныс алумен өзара байланыстарын 7 – ші бөлімде қарастырдық (7.1 – ші сурет). Біршама өзгеше формада 9.1 суретте қоршаған орта мен тірі ағзалардың арасындағы энергия алмасу жайлы көрсетілген.
9.1. Тыныс алу дегеніміз не?
Тыныс алу органикалық заттардың қышқылдану нәтижесінде энергияның бөліну үрдісі. Бұл энергия АТФ формасында тірі жасушалар үшін байланысқа шығады. Осы биохимиялық үрдіс жасушада өткенде, бұл үрдісті жасушалық тыныс алу деп атайды. Егер оған қажет болса, онда тыныс алуды аэробты деп атайды; егер реакция оттектің қажетінсіз өтсе онда мұны анаэробты деп атайды.
Жасушалық тыныс алу үшін сабстрат ретінде көмірсулар (мысалы, глюкоза) немесе майлпр қызмет атқарады. Олар ферментативті рекауциялар қатарында бірте – бірте ыдырайды. Әрбір осындай реакцияда аз мөлшерде энергия бөлінеді және сол энергияның бөлігі заттың молекуласында жинақталады. Бұл зат молекуласын аденозинтрифосфат (АТФ) болып табылады. Қалған энергия жылу ретінде таралады. АТФ жасушаларда энергия тасушы ретінде рөл атқарады. Сол молекуласында сақталған энергия реакцияға жұмсалады.
9.2. АТФ
9.2.1. АТФ құрылымы
9.2 суретінде АТФ құрылысының 2 түрдегі көрінісі бейнеленген. Аденозиндифосфат (АДФ) және аденозинтрифосфат (АТФ) байланыстырушы нуклеотидтер класына жатады. Нуклеотид молекуласы 5 бұрышты қанттан, азотты негізден және фосфорлы қышқылынан тұрады (5.6.1 сурет). АМФ молекуласында қант рибоза түрінде, ал негізі аденин үрінде көрсетілген. АДФ молекуласында 2 фосфат топтары, ал АТФ молекуласында 3 фосфат тобы бар.
9.1. Тыныс алу дегеніміз не?
Тыныс алу органикалық заттардың қышқылдану нәтижесінде энергияның бөліну үрдісі. Бұл энергия АТФ формасында тірі жасушалар үшін байланысқа шығады. Осы биохимиялық үрдіс жасушада өткенде, бұл үрдісті жасушалық тыныс алу деп атайды. Егер оған қажет болса, онда тыныс алуды аэробты деп атайды; егер реакция оттектің қажетінсіз өтсе онда мұны анаэробты деп атайды.
Жасушалық тыныс алу үшін сабстрат ретінде көмірсулар (мысалы, глюкоза) немесе майлпр қызмет атқарады. Олар ферментативті рекауциялар қатарында бірте – бірте ыдырайды. Әрбір осындай реакцияда аз мөлшерде энергия бөлінеді және сол энергияның бөлігі заттың молекуласында жинақталады. Бұл зат молекуласын аденозинтрифосфат (АТФ) болып табылады. Қалған энергия жылу ретінде таралады. АТФ жасушаларда энергия тасушы ретінде рөл атқарады. Сол молекуласында сақталған энергия реакцияға жұмсалады.
9.2. АТФ
9.2.1. АТФ құрылымы
9.2 суретінде АТФ құрылысының 2 түрдегі көрінісі бейнеленген. Аденозиндифосфат (АДФ) және аденозинтрифосфат (АТФ) байланыстырушы нуклеотидтер класына жатады. Нуклеотид молекуласы 5 бұрышты қанттан, азотты негізден және фосфорлы қышқылынан тұрады (5.6.1 сурет). АМФ молекуласында қант рибоза түрінде, ал негізі аденин үрінде көрсетілген. АДФ молекуласында 2 фосфат топтары, ал АТФ молекуласында 3 фосфат тобы бар.
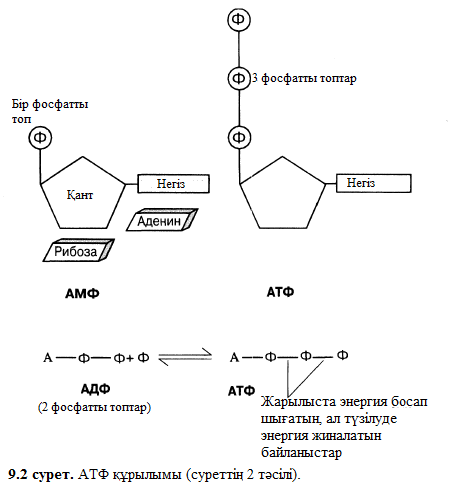
9.2.2. АТФ мәні
АТФ – тың АДФ – ке және бейорганикалық фосфат (Фн) – ды ыдырату кезінде энергия бөлінеді:
АТФ – тың АДФ – ке және бейорганикалық фосфат (Фн) – ды ыдырату кезінде энергия бөлінеді:
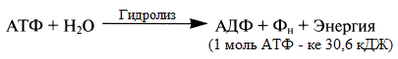
Реакция судың жұтылуымен өтеді, яғни гидролиз түрінде (3 бөлімде бұл биохимиялық реакция түрі жайлы кездеседі). АТФ – тан ұсақтап жарылған 3 –ші фосфатты топ жасушада бейорганикалық фосфат (Фн) түрінде қалып қояды. Бұл рекациядан шығатын еркін энергиясы 1 моль АТФ ке 30,6 кДЖ – лін құрайды.
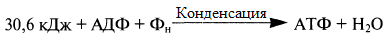
Конденсация реакциясынла су бөлініп шығады. Фосфаттың АДФ – ке қосылысы фосфорилдеу реакциясы деп аталады. жоғары деңгейлі көрсетілген екеуін байланыстыруға болады:
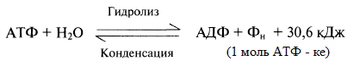
Бұл реакцияны АТФаза деп аталатын фермент катализаторлайды.
Айтып кеткеніміздей, барлық жасушалардың жұмыс жасауы үшін энергия қажет және әрбір ағзаның жасушалары үшін энергия көзі ретінде АТФ қызмет етеді. Сондықтан АТФ – ты жасушаның «әмбебеп энергия тасымалдаушысы» немесе «энергетикалық валютасы» деп атайды. Сәйкес аналогия қызметін электрлі ұуат көзі атқарады. Оларды біз барлық жағдайда да қолданатынымызды еске түсірейік. Біз олардың көмегі арқылы бір жағдайда жарық, ал екінші жағдайда дабыс, кейде механикалық қозғалыс, ал кейде бізге олардан жеке электр энергиясы да қожет болады.
Қуат көзінің пайдасы сол және бір энергия көзі – батареяны – қандай жерге қоюымызға байланысты әртүрлі мақсаттарда қолдана беруге болады. Бұл қызметті жасушадағы АТФ те атқарады. Ол энергияны бұлшық еттің қысқаруын, жүйке импулсьтерін таратуға заттардың белсенді орын ауысуында немесе ақуыздың синтезделуіне және белсенді жасушаның басқа түрлеріне жұмсайды. Бұл үшін ол тек жасушаның сәйкес аппаратына қосылуы керек.
Аналогияны жалғастыруға да болады. Батареяны бірінші дайындау қажет, ал олардың АТФ сынды кейбір түрлерін (аккумуляторлы) қайтадан зарядтап алуға болады. Батареяларды фабрикада дайындаған кещде оларға энергияның белгілі бір мөлшері салынуы керек.
АТФ – тың синтезделуі үшін де энергия қажет; оның қайнар көзі ретінде тыныс алу үрдісіндегі органикалық заттардың қышқылданған түрлері қызмет атқарады. АДФтың фосфорлау кезінде энергия қыщқылдану үрдісінде шығарылатын болған соң, мұндай фосфорлауды қышқылданған деп атайды. Фотосинтез кезінде АТФ жарық энергиясының есебінен түзіледі. Бұл үрдісті фотофосфориленген (7.6.2 бөлім) деп атайды. Жасушада АТФ – тың көп мөлшерін түзетін «фабрикаларда» бар. Бұл митохондриялар; олардың ішінде аэробты тыныс алу үрдісі кезінде АТФ – тың түзілетін химиялы «жинақтауыш сызықтары» болады. Әйтеуір, жасушада қуаты таусылып қалған «аккумулятор» қуаттана бастайды.АТФ өз құрамындағы соңғы энергияны шығарып болып, АДФ және Фн– ге айналған кезде ол органикалық заттардың жаңа бөлігінің қышқылдануынан алатын энергияның арқасында АДФпен Фн– нен қайтадан тез синтезделінеді.
АТФ – тың мөлшері әрбір уақытта көп бола бермейді. Сондықтан АТФ – тен оның депосын емес, энергия тасымалдаушы ретінде қараған жөн. Энергияны көп сақтау қызметін май және гликоген сынды заттар атқарады. Жасушалар АТФ деңгейіне өте сезімтал. Оны қолдану жылдамдығы үлкейген сайын осы деңгейді ұстап отырған тыныс алу үрдісінің де жылдамдығы үлкейе түседі.
Жасушаның тыныс алу мен энергия қабылдаумен бірге жүретін үрдіс арасындағы АТФ – тың рөлі 9.3 суретінде көрсетілген. Оның кескіні қарапайым көрінгенімен, ол өте маңызды заңнаманы бейнелейді.
9.2. 9. 3 суреттегі кескінді толтыр, мұндағы – мақсат - нәтижесінде күн энергиясы глюкоза түрінде жинақталатынын көрсету.
Осылайша тыныс алудың функциясы АТФ – ты шығаруға жұмсалады деген қорытындыға келуге болады.
Айтып кеткеніміздей, барлық жасушалардың жұмыс жасауы үшін энергия қажет және әрбір ағзаның жасушалары үшін энергия көзі ретінде АТФ қызмет етеді. Сондықтан АТФ – ты жасушаның «әмбебеп энергия тасымалдаушысы» немесе «энергетикалық валютасы» деп атайды. Сәйкес аналогия қызметін электрлі ұуат көзі атқарады. Оларды біз барлық жағдайда да қолданатынымызды еске түсірейік. Біз олардың көмегі арқылы бір жағдайда жарық, ал екінші жағдайда дабыс, кейде механикалық қозғалыс, ал кейде бізге олардан жеке электр энергиясы да қожет болады.
Қуат көзінің пайдасы сол және бір энергия көзі – батареяны – қандай жерге қоюымызға байланысты әртүрлі мақсаттарда қолдана беруге болады. Бұл қызметті жасушадағы АТФ те атқарады. Ол энергияны бұлшық еттің қысқаруын, жүйке импулсьтерін таратуға заттардың белсенді орын ауысуында немесе ақуыздың синтезделуіне және белсенді жасушаның басқа түрлеріне жұмсайды. Бұл үшін ол тек жасушаның сәйкес аппаратына қосылуы керек.
Аналогияны жалғастыруға да болады. Батареяны бірінші дайындау қажет, ал олардың АТФ сынды кейбір түрлерін (аккумуляторлы) қайтадан зарядтап алуға болады. Батареяларды фабрикада дайындаған кещде оларға энергияның белгілі бір мөлшері салынуы керек.
АТФ – тың синтезделуі үшін де энергия қажет; оның қайнар көзі ретінде тыныс алу үрдісіндегі органикалық заттардың қышқылданған түрлері қызмет атқарады. АДФтың фосфорлау кезінде энергия қыщқылдану үрдісінде шығарылатын болған соң, мұндай фосфорлауды қышқылданған деп атайды. Фотосинтез кезінде АТФ жарық энергиясының есебінен түзіледі. Бұл үрдісті фотофосфориленген (7.6.2 бөлім) деп атайды. Жасушада АТФ – тың көп мөлшерін түзетін «фабрикаларда» бар. Бұл митохондриялар; олардың ішінде аэробты тыныс алу үрдісі кезінде АТФ – тың түзілетін химиялы «жинақтауыш сызықтары» болады. Әйтеуір, жасушада қуаты таусылып қалған «аккумулятор» қуаттана бастайды.АТФ өз құрамындағы соңғы энергияны шығарып болып, АДФ және Фн– ге айналған кезде ол органикалық заттардың жаңа бөлігінің қышқылдануынан алатын энергияның арқасында АДФпен Фн– нен қайтадан тез синтезделінеді.
АТФ – тың мөлшері әрбір уақытта көп бола бермейді. Сондықтан АТФ – тен оның депосын емес, энергия тасымалдаушы ретінде қараған жөн. Энергияны көп сақтау қызметін май және гликоген сынды заттар атқарады. Жасушалар АТФ деңгейіне өте сезімтал. Оны қолдану жылдамдығы үлкейген сайын осы деңгейді ұстап отырған тыныс алу үрдісінің де жылдамдығы үлкейе түседі.
Жасушаның тыныс алу мен энергия қабылдаумен бірге жүретін үрдіс арасындағы АТФ – тың рөлі 9.3 суретінде көрсетілген. Оның кескіні қарапайым көрінгенімен, ол өте маңызды заңнаманы бейнелейді.
9.2. 9. 3 суреттегі кескінді толтыр, мұндағы – мақсат - нәтижесінде күн энергиясы глюкоза түрінде жинақталатынын көрсету.
Осылайша тыныс алудың функциясы АТФ – ты шығаруға жұмсалады деген қорытындыға келуге болады.
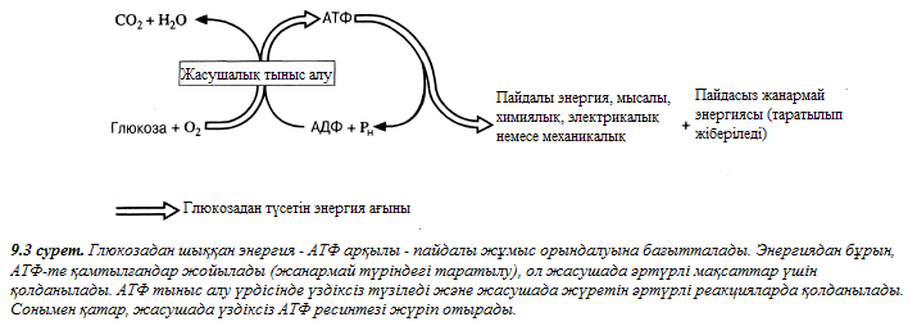
Жоғарыда айтылғандарды қысқаша қорытындылайық:
9.3.1. Тыныс алу субстраттары
Тыныс алу – бұл тыныс алу субстраты болып табылатын органикалық заттардың қышқылдануы. Тыныс алу үшін субстраттар қызметін көміртек, май және нәруыз атқарады.
Көміртектер. Көптеген жасушалар субстрат ретінде осы көміртекте ғана қолданады. Помисахаридтер(өсімдіктерде крахмал және жануарлар мен саңырауқұлақтарда) тыныс алу үрдісінде тек олар моносохоридтерге дейін гидролизденген кезде ғана пайда бола бастайды.
Липидтер (майлар немесе сарымай). Липидтер «негізгі қорды» құрайды және көміртек қорлары біткен кезде негізгімәселеге көшеді. Олар күні бұрын глицерол мен май қышқылдарына дейін гидролизденуі тиіс. Май қышқылдары энегия мен кейбір жасушаларға бай, мысалы бұлшықеттілер, нормада нақты солардан өздеріне қажетті энергия бөлшегін алады.
Нәруыздар. Сондықтан нәруыздар өзге де маңызды функциялар қатарын құрайды, олар энергия өндірісіне, тек қана көміртектің барлық қорлары біткеннен кейін және ұзаққа соылған аштықтағы (8.9.3 бөлімі) майлылықтан кейін қолданылады. Нәруыздар шамалап аминқышқылдарына дейін гидролизденеді, ал аминқышқылдар аминсіздендіреді (өздерінің аминтоптарын жояды). Нәтижесінде, түзілген аминсіздендіретін қышқыл Кребс цикліне қатыстырылады (9.3.5 бөлім) немесе бірінші кейін қышқылдыққа (9.3.5 бөлімі) айналу үшін май қышқылына айналады.
9.3.2. Кейбір кілт реакциялар
Жасуша тыныс алуында реакцияның 2 типі маңызды рөл ойннайды – ұышұылдану және карбоксилсіздендіру.
Қышқылдану
Жасушада қашқылдың реакцияларының 3 типі болады:
- АТФ – тің ФДВ және бейорганикалық фосфоттан синтезделуі үшін 1 моль АТФ – қа 30,6 кДЖ энергия қажет.
- АТФ барлық тірі жасушаларда кездеседі және әмбебеп энергия тасымалдаушы болып табылады. басқа энергия тасымалдайтындар қолданылмайды. Юұл жұмысты жеңілдетеді – маңызды жасушалық аппарат өте қарапайым және жұмысты да жақсы үнемді жасайды.
- АТФ энергияны жасушаның әрбір бөлігіне энергияға қажетсініп тұрған жеріне оңай жеткізеді.
- АТФ энергияны тез сыртқа шығарады. Бұл үшін тек бір ғана реакция – гидролиз қажет.
- АТФ – тың АДФ және органикалық фосфоттан (тыныс алу үрдісі жылдамдығы) түзілуі сәйкес қажеттіліктермен тез байланысып кетеді.
- АТФ тыныс алу кезіндегі глюкоза сияқты органикалық заттардың қышқылдануынан түзілетін химиялық энергияның және фотосинтез кезінде – күн энергиясы есебінен синтезделеді АТФ – тің АДФ пен бейорганикалық фосфаттан түзілуі фосфориленген реакция деп аталады. Егер фосфорилдеуге арналған энергияны қышқылдайтын болса,онда қышқылданған фосфорел (прецесс тыныс алу кезінде шығарылады) туралы айтылады,егер де фосфорилдеуге тарық энергиясы қолданылатын болса,онда процесс фотофосфорилденген (бұл фотосинтезде белгілі бір орынға ие) деп аталады.
9.3.1. Тыныс алу субстраттары
Тыныс алу – бұл тыныс алу субстраты болып табылатын органикалық заттардың қышқылдануы. Тыныс алу үшін субстраттар қызметін көміртек, май және нәруыз атқарады.
Көміртектер. Көптеген жасушалар субстрат ретінде осы көміртекте ғана қолданады. Помисахаридтер(өсімдіктерде крахмал және жануарлар мен саңырауқұлақтарда) тыныс алу үрдісінде тек олар моносохоридтерге дейін гидролизденген кезде ғана пайда бола бастайды.
Липидтер (майлар немесе сарымай). Липидтер «негізгі қорды» құрайды және көміртек қорлары біткен кезде негізгімәселеге көшеді. Олар күні бұрын глицерол мен май қышқылдарына дейін гидролизденуі тиіс. Май қышқылдары энегия мен кейбір жасушаларға бай, мысалы бұлшықеттілер, нормада нақты солардан өздеріне қажетті энергия бөлшегін алады.
Нәруыздар. Сондықтан нәруыздар өзге де маңызды функциялар қатарын құрайды, олар энергия өндірісіне, тек қана көміртектің барлық қорлары біткеннен кейін және ұзаққа соылған аштықтағы (8.9.3 бөлімі) майлылықтан кейін қолданылады. Нәруыздар шамалап аминқышқылдарына дейін гидролизденеді, ал аминқышқылдар аминсіздендіреді (өздерінің аминтоптарын жояды). Нәтижесінде, түзілген аминсіздендіретін қышқыл Кребс цикліне қатыстырылады (9.3.5 бөлім) немесе бірінші кейін қышқылдыққа (9.3.5 бөлімі) айналу үшін май қышқылына айналады.
9.3.2. Кейбір кілт реакциялар
Жасуша тыныс алуында реакцияның 2 типі маңызды рөл ойннайды – ұышұылдану және карбоксилсіздендіру.
Қышқылдану
Жасушада қашқылдың реакцияларының 3 типі болады:
- Молекулярлы оттектің қышқылдануы
- Сутектің сутексіздендіру қышқылдануы. Глюкозалар қышқылдануының аэробты тыныс алуы дәйекті реакция сутексіздендіру жолымен болады. Әрбір ұсақтап жарылған сутексіздендіру де сутегі бұл жағдайда сутегі тасымалдау мысы деп аталатын коферменттің қалпына келтірілуіне қолданылады:

Бұл реакциялардың басым көпшілігі сутек тасымалдаушысы НАД (никотинамидадениндинуклеотид) коферметіне қызмет ететін митохондрияда болады:
НАД + 2Н → НАД ∙ Н2
немесе одан да нақтылау айтса,
НАД+ +2Н →НАД ∙ Н + Н+
НАД ∙ Н (қалыпқа келтірілген НАД) кейін тағы да энергияның босап шығуымен қышқылдануға ұшырайды. Дегидриттерді катализдейтін ферменттер дегидрогеназалар деп аталады. «Дегидриреттеу» реакциясның нәтижесінде глюкозадан бөлініп шығатын сутегі сітегінің жеткізушілеріне беріледі. Бұл сутегі оттегіден суға дейін қышқылданады, кейін босап шығатын энергия АТФ синтезіне қатысады. Қышқылдану кезінде энергияның бөлініп шығатындығын сутегі орналасқан сынауыққа жанып тұрған майшамды қойған кезде ғана көруге болады. Осы кезде жеңіл әрі қысқа шапалақ ретіндегі бір дыбыс естіледі. Тордың ішінде энергия дәл осындай күйде бөлініп шығады,бірақ ол қышқылды – қалыпқа келтірілетін реакция кезінде бөлініп шығады.
3. Электрондардың орын ауыстыруы.Бұндай жағдай, мысалы, темірдің бір ионы пішінінен (Ғе2+) басқасына ауысуыда жүзеге асады.
Ғе2+ →Ғе3+ + е–
Электрондар бір бірінен екіншісіне сутегі секілді ауыса алады. Осындай бірігулер электрон тасымалдаушылар деп аталады. бұл үрдіс митохондрияларда жүзеге асады (9.3.5 бөлім).
Карбоксилсіздендіру
Карбоксилсіздендіру– бұл белгілі бір бірігуден көміртегінің СО2 түзуі арқылы шығуы болып табылады. глюкоза молекулаларының құрамында сітегі мен оттегіден басқа көміртегінің алты атомы бар. Жоғарыда айтылған реакцияларға тек сутегі керек болғандықтан, көміртек карбоксилсіздендіру кезінде жойылып кетеді. Осы кезде пайда болған көміртек диоксиді аэробты тыныс алудың «жағымсыз өнім» рөлін атқарады.
НАД + 2Н → НАД ∙ Н2
немесе одан да нақтылау айтса,
НАД+ +2Н →НАД ∙ Н + Н+
НАД ∙ Н (қалыпқа келтірілген НАД) кейін тағы да энергияның босап шығуымен қышқылдануға ұшырайды. Дегидриттерді катализдейтін ферменттер дегидрогеназалар деп аталады. «Дегидриреттеу» реакциясның нәтижесінде глюкозадан бөлініп шығатын сутегі сітегінің жеткізушілеріне беріледі. Бұл сутегі оттегіден суға дейін қышқылданады, кейін босап шығатын энергия АТФ синтезіне қатысады. Қышқылдану кезінде энергияның бөлініп шығатындығын сутегі орналасқан сынауыққа жанып тұрған майшамды қойған кезде ғана көруге болады. Осы кезде жеңіл әрі қысқа шапалақ ретіндегі бір дыбыс естіледі. Тордың ішінде энергия дәл осындай күйде бөлініп шығады,бірақ ол қышқылды – қалыпқа келтірілетін реакция кезінде бөлініп шығады.
3. Электрондардың орын ауыстыруы.Бұндай жағдай, мысалы, темірдің бір ионы пішінінен (Ғе2+) басқасына ауысуыда жүзеге асады.
Ғе2+ →Ғе3+ + е–
Электрондар бір бірінен екіншісіне сутегі секілді ауыса алады. Осындай бірігулер электрон тасымалдаушылар деп аталады. бұл үрдіс митохондрияларда жүзеге асады (9.3.5 бөлім).
Карбоксилсіздендіру
Карбоксилсіздендіру– бұл белгілі бір бірігуден көміртегінің СО2 түзуі арқылы шығуы болып табылады. глюкоза молекулаларының құрамында сітегі мен оттегіден басқа көміртегінің алты атомы бар. Жоғарыда айтылған реакцияларға тек сутегі керек болғандықтан, көміртек карбоксилсіздендіру кезінде жойылып кетеді. Осы кезде пайда болған көміртек диоксиді аэробты тыныс алудың «жағымсыз өнім» рөлін атқарады.
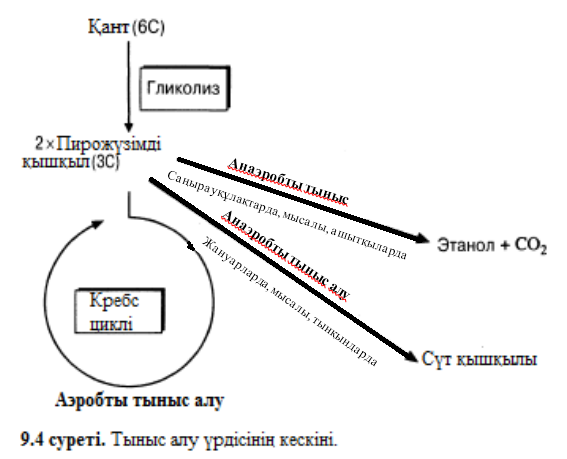
9.3.3. Жасушалы тыныс алудың жалпылай көрінісі
Жасушалы тыныс алуды зерттемес бұрын, онымен толығырақ танысып алған жөн. 9.4 суретте аэробты және анаэробты тыныс алу көрсетілген. Ескерте кететін жағдай аэробты тыныс алудың жолы біреу болса, анаэробтыда – екеу. Екі тыныс алудың алғашқы жолы бірдей және ол – гликолиз.
9.3.4. Гликолиз
Гликолиз дегеніміз – глюкозаның пирожүзім қышқылға дейін қышқылдануы. 9.5 суретте көрсетілгендей, глюкозаның бір молекуласынан пирожүзімді қышқылдың 2 молекуласынан түзеді. Бұл үрдіс митохондрияда емес, жасушашаның цитоплазмсында жүзден асады.
Жасушалы тыныс алуды зерттемес бұрын, онымен толығырақ танысып алған жөн. 9.4 суретте аэробты және анаэробты тыныс алу көрсетілген. Ескерте кететін жағдай аэробты тыныс алудың жолы біреу болса, анаэробтыда – екеу. Екі тыныс алудың алғашқы жолы бірдей және ол – гликолиз.
9.3.4. Гликолиз
Гликолиз дегеніміз – глюкозаның пирожүзім қышқылға дейін қышқылдануы. 9.5 суретте көрсетілгендей, глюкозаның бір молекуласынан пирожүзімді қышқылдың 2 молекуласынан түзеді. Бұл үрдіс митохондрияда емес, жасушашаның цитоплазмсында жүзден асады.
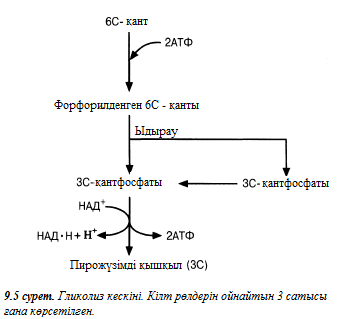
Сондықтан оған ауа керек емес. үрдіс 3 бөліктен тұрады:
- Қанттың фосфорлануы. Бұо реакцияның нәтижесінде қант белсенділеді, себебі оның ореакциялы қасиеті көбейе түседі. Белсендену барысында АТФ кішкентай мөлшері пайдаланылады. Тыныс алудың негізгі ойы АТФ – ты «жоғары көтеру» болғандықтан, оны пайдалану мақсатқа сай емес. бірақ, бұған пайдалы жағынан да қарауға бола, кейін осының бәрі АТФ – ты қайта туғызуға көмектесееді.
- Фосфориенген 6С қанттың екі 3С қантфосфатқа ыдырауы. Осымен «гликолиздің» атауы да пайда болған. Пайда болған екі қантфосфаты – изомерлер. Келесі айналымға өтер алдында екеуінің біреуі біреуіне қосылады да, екі 3С қантфосфат пайда болады.
- Сутектің бөлінуі арқылы қышқылдану. Әрбір 3С – қантфосфат пирожүзімді қышқышға айналады. Осы арқылы қалпына келтірілген НАД – тың бір молекуласы мен АТФ – тың 2 молекуласына түзілуімен сутексіздендіру жүреді. Жалпылама жол (екі молекуладан 3С – қантфосфатына) мыналардан: қалпына келтірілген НАД – тың екі молекуласы және АТФ – тың өрт молекуласынан (9.5 суреті) тұрады.
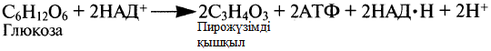
Гликолиз үрдісінде қолданылатын және онда әртүрлі заттардың шығуы 9.1 кестесінде көрсетілген.

Липидтердің тыныс алу үрдісінде қолданылатын глицерол гликолиз жолына түсетін 3С – қантфосфатына оңай айналып кетеді. Бұл жерде АТФ – тың бір молекуласы тарқап кетеді де, 3 молекуласы түзіледі.
Қышқылдың пирожүзімдісінің соңғы тағдыры жасушада оттектің жоқ болуынан тәуелді болады. Егер оттек болса, онды пирожүзімде қышқыл митохондрияға СО2 мен суға (аэробты тыныс алу) дейінгі қышқылдануға ауысады. Ал егер оттек болмаса, онда ол не этанолға, не сүт қышқылына (анаэробты тыныс алу) айналады (9.4 суреті).
9.3.5. Аэробты тыныс алу
Гликолиз үрдісіндегі түзілетін аэробты тыныс алуда пирожүзімді қышқыл соңғы нәтижесінде толығымен оттектен СО2 мен суға дейін қышқылданады. Алғашқы фазада пирожүзімді қышқыл СО2 мен көміртектің түзілуімен жарылады. Бұл үрдіс митохондрия матриксіне өтеді және Кребс циклі деп аталатын реакциялардың жүйелілігін құрайды. Екінші фазада екінші сатыда жарылатын көміртек тыныс алу тізьектері деп аталатын қышқыл – қалпына келтірілген реакциялар қатары арқылы оттектің соңғы молекулярлы есебінде суға дейн қышқылданады. Бұл криста (митохондрияның ішкі мембранасындағы тараққа ұқсас бүкпелерінде болады) деп аталатындарда болады.
Аэробты тыныс алудың бастапқы сатылары 9.6 суреттерінде берілген.
Гликолиз бен Кребс циклі арасындағы өткізгіш саты
Пирожүзімді қышқылдың әрбір молекуласы митохондрия матриксіне енеді және мұнда ацетиь тобы (СН3СОО –) А коферменті (немесе қысқартылғанда КоА) деп аталатын затпен байланысады, нәтижесінде А ацетилкоферменті (ацетил – КоА) түзіледі.Ацетиьді топ көміртектің екі атомына ие (2 Сl, сол үшін, ол пайда болу үшін парожүзімді қышқыл 13С) көміртек атомын жұмсауы қажет.
СО2 күйінде атомның айырылысуы карбоксилсіздендіру деп аталады. бұл – қышқылды карбоксилсіздендіру, себебі дигидритті қышқылдау реакциясымен жүзеге асырылады. Соңында қалпына келтірілген НАД пайда болады.
Кребс циклі
Бұл циклдің аты 1930 жылы оны ашқан зерттеуші – сэр Гомс Кребстің құрметіне қойылған. Сонымен қатар, оны «үшкарбонатты қышқыл циклі» және «лимон қышқыл циклі» деп атайды. Сондықтан, дәл осы қышқылдар қатысады.
Кребс циклі (9.1 сурет) митохонжрия матриксінде өтеді. Ауетильді топтар (2С) циклге топ боып, НС қосылысы – қымыздық сіркесулы қышқылға қосылып, соңында лимон қышқыл (6С) пайда болды. Содан соң, реакция циклі болады, онда циклге келген ацетильді топтар карбоксилсіздендіреді және тасымалдаушыларға қосылған сутектің 4 жұп атомын босатумен дигидритталанады, соңында қалпына келтірілген НАД – ты 3 молекуласы және қалпына келтірілген ФАД – тың 1 молекуласы пайда болады.
Қышқылдың пирожүзімдісінің соңғы тағдыры жасушада оттектің жоқ болуынан тәуелді болады. Егер оттек болса, онды пирожүзімде қышқыл митохондрияға СО2 мен суға (аэробты тыныс алу) дейінгі қышқылдануға ауысады. Ал егер оттек болмаса, онда ол не этанолға, не сүт қышқылына (анаэробты тыныс алу) айналады (9.4 суреті).
9.3.5. Аэробты тыныс алу
Гликолиз үрдісіндегі түзілетін аэробты тыныс алуда пирожүзімді қышқыл соңғы нәтижесінде толығымен оттектен СО2 мен суға дейін қышқылданады. Алғашқы фазада пирожүзімді қышқыл СО2 мен көміртектің түзілуімен жарылады. Бұл үрдіс митохондрия матриксіне өтеді және Кребс циклі деп аталатын реакциялардың жүйелілігін құрайды. Екінші фазада екінші сатыда жарылатын көміртек тыныс алу тізьектері деп аталатын қышқыл – қалпына келтірілген реакциялар қатары арқылы оттектің соңғы молекулярлы есебінде суға дейн қышқылданады. Бұл криста (митохондрияның ішкі мембранасындағы тараққа ұқсас бүкпелерінде болады) деп аталатындарда болады.
Аэробты тыныс алудың бастапқы сатылары 9.6 суреттерінде берілген.
Гликолиз бен Кребс циклі арасындағы өткізгіш саты
Пирожүзімді қышқылдың әрбір молекуласы митохондрия матриксіне енеді және мұнда ацетиь тобы (СН3СОО –) А коферменті (немесе қысқартылғанда КоА) деп аталатын затпен байланысады, нәтижесінде А ацетилкоферменті (ацетил – КоА) түзіледі.Ацетиьді топ көміртектің екі атомына ие (2 Сl, сол үшін, ол пайда болу үшін парожүзімді қышқыл 13С) көміртек атомын жұмсауы қажет.
СО2 күйінде атомның айырылысуы карбоксилсіздендіру деп аталады. бұл – қышқылды карбоксилсіздендіру, себебі дигидритті қышқылдау реакциясымен жүзеге асырылады. Соңында қалпына келтірілген НАД пайда болады.
Кребс циклі
Бұл циклдің аты 1930 жылы оны ашқан зерттеуші – сэр Гомс Кребстің құрметіне қойылған. Сонымен қатар, оны «үшкарбонатты қышқыл циклі» және «лимон қышқыл циклі» деп атайды. Сондықтан, дәл осы қышқылдар қатысады.
Кребс циклі (9.1 сурет) митохонжрия матриксінде өтеді. Ауетильді топтар (2С) циклге топ боып, НС қосылысы – қымыздық сіркесулы қышқылға қосылып, соңында лимон қышқыл (6С) пайда болды. Содан соң, реакция циклі болады, онда циклге келген ацетильді топтар карбоксилсіздендіреді және тасымалдаушыларға қосылған сутектің 4 жұп атомын босатумен дигидритталанады, соңында қалпына келтірілген НАД – ты 3 молекуласы және қалпына келтірілген ФАД – тың 1 молекуласы пайда болады.
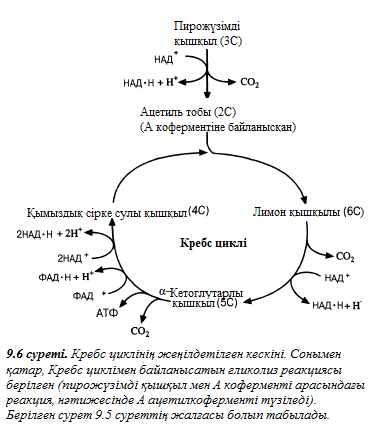
Сонымен қатар, әрбір циклдің айналымы АТФ – ң 1 молекуласын береді (2 глюкозаның 1 молекуласымен 2 ауетельді топ пайда болатынын еске түсірейік, сонда глюкозаның әрбір молекуласы қышқылдану қышқылдану үшін, циклдің екі аайналымы қажет). Циклдің соңында қымыздық сіркесудықышқыл регенерацияланады және енді өзіне жаңа ацетильді топты ұоса алады.
Аэробты тыныс алудың осы сатыдағы жалпы теңгерімі 9.2 кестесінде келтірілген.
Аэробты тыныс алудың осы сатыдағы жалпы теңгерімі 9.2 кестесінде келтірілген.

Жинақталған теңдеу мынадай қылып жазыла алады:

Глюкозаның молекуласындағы барлық сутек соңында тасымалдаушыларда (НАД және ФАД) пайда болады. Барлық көміртек СО2 күйінде жоғалады. (Бұл теңдеуде судың 6 молекуласының болуы таңғаларлық болуы мүмкін. Су оттектің қайнар көзі ретінде карбоксилсіздендіру реакциясында керек. дәл осы жаратылыс СО2 – де оттектің бір бөлігіне ие. Бұл, негізі санауға қажет емес бөлік болып табылады).
Тынысалу тізбегі және қышқылды форфолирлеу
2 тасумалдаушылардағы сутек (қалпына келтірілген НАД – тың 10 молекуласы және қалпына келтірілген ФАД – тың 2 молекуласы) енді митохонжрияның ішкі мембранасына бағыттайды.
Тынысалу тізбегі және қышқылды форфолирлеу
2 тасумалдаушылардағы сутек (қалпына келтірілген НАД – тың 10 молекуласы және қалпына келтірілген ФАД – тың 2 молекуласы) енді митохонжрияның ішкі мембранасына бағыттайды.
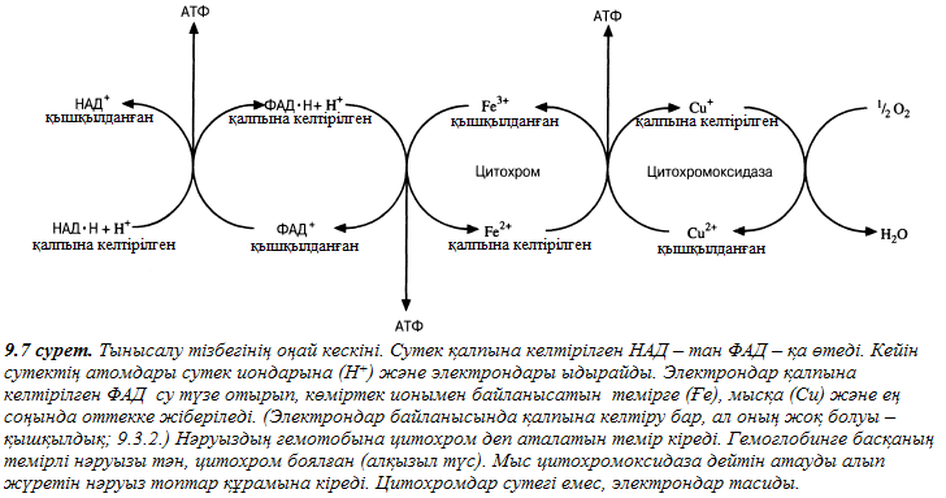
Бұл мембрана кристалар деп аталатын өзінің көлемін үлкейтетін қатпарлар түзеді (9. 12 сурет). Сутек бұл – жанармай. Молекулалы оттекпен оның қышқылдануынан су және энергия түзілетінін белгілеген болатынбыз.
2Н2 + О2 2Н2О+энергия
Энергияның бір бөлігі АДФ – дағы АТФ – тың синтезіне және қышқылды фосфорилену кезіндегі бейорганикалық фосфотты қолданылады (9.22 сурет). Энергия тек бір реакцияның ішінде пайда болмайды. Үрдісті бірнеше топтарға бөлеті, оның ішінде АТФ синтезіне жеткілікті энергия бөлінетіні де болады. Берілген реакцияның реті тынысалу тізбегі ретінде белгілі. Тынысалу тізбегіндегі сутектің тысамалдаушылары мен электрондары қатысады.
Бұл тізбек оттекпен бітеді. Сутек немесе электрондар бір тасылмалдаушыдан екіншісіне энергетикалық мағынада «астына» қозғалады. Бұл соңында молекулярлы оттекті суға дейін қалпына келгенше жаслғасады. Әрбір сатыда белгілі бір мөлшерде энергия бөлінеді: және әрбір пунктте бұл АТФ синтезімен жүргізілген (бұл пункттер 9.7 суретінде сызықтармен белгіленген). 9.7 суретіндегі жазуда тынысалу тізбегінде одан да анығырақ айтылған. Соңында мысы бар тасымалдаушы цитохромонсидаза әсер етеді. Цианид сынаппен байланысады, содан кейін оттек олмен енді байланыса алмайды.
9.7 суретінде тыныс алу тізбегіне түсетін әрбір қалпына келтірілген НАД – тың молекуласында сутегі мен электрондардың оттекке өтуінде АТФ – тың 3 молекуласы түзіледі. Бірақ қайта келтірілген ФАД – тың әрбір молекуласы АТФ – тың бар жоғы 2 молекуласы түзіледі, себебі қайта келтірілген ФАД өте төмен энергетикалық деңгейде тыныс алу тізбегіне түседі.
2Н2 + О2 2Н2О+энергия
Энергияның бір бөлігі АДФ – дағы АТФ – тың синтезіне және қышқылды фосфорилену кезіндегі бейорганикалық фосфотты қолданылады (9.22 сурет). Энергия тек бір реакцияның ішінде пайда болмайды. Үрдісті бірнеше топтарға бөлеті, оның ішінде АТФ синтезіне жеткілікті энергия бөлінетіні де болады. Берілген реакцияның реті тынысалу тізбегі ретінде белгілі. Тынысалу тізбегіндегі сутектің тысамалдаушылары мен электрондары қатысады.
Бұл тізбек оттекпен бітеді. Сутек немесе электрондар бір тасылмалдаушыдан екіншісіне энергетикалық мағынада «астына» қозғалады. Бұл соңында молекулярлы оттекті суға дейін қалпына келгенше жаслғасады. Әрбір сатыда белгілі бір мөлшерде энергия бөлінеді: және әрбір пунктте бұл АТФ синтезімен жүргізілген (бұл пункттер 9.7 суретінде сызықтармен белгіленген). 9.7 суретіндегі жазуда тынысалу тізбегінде одан да анығырақ айтылған. Соңында мысы бар тасымалдаушы цитохромонсидаза әсер етеді. Цианид сынаппен байланысады, содан кейін оттек олмен енді байланыса алмайды.
9.7 суретінде тыныс алу тізбегіне түсетін әрбір қалпына келтірілген НАД – тың молекуласында сутегі мен электрондардың оттекке өтуінде АТФ – тың 3 молекуласы түзіледі. Бірақ қайта келтірілген ФАД – тың әрбір молекуласы АТФ – тың бар жоғы 2 молекуласы түзіледі, себебі қайта келтірілген ФАД өте төмен энергетикалық деңгейде тыныс алу тізбегіне түседі.

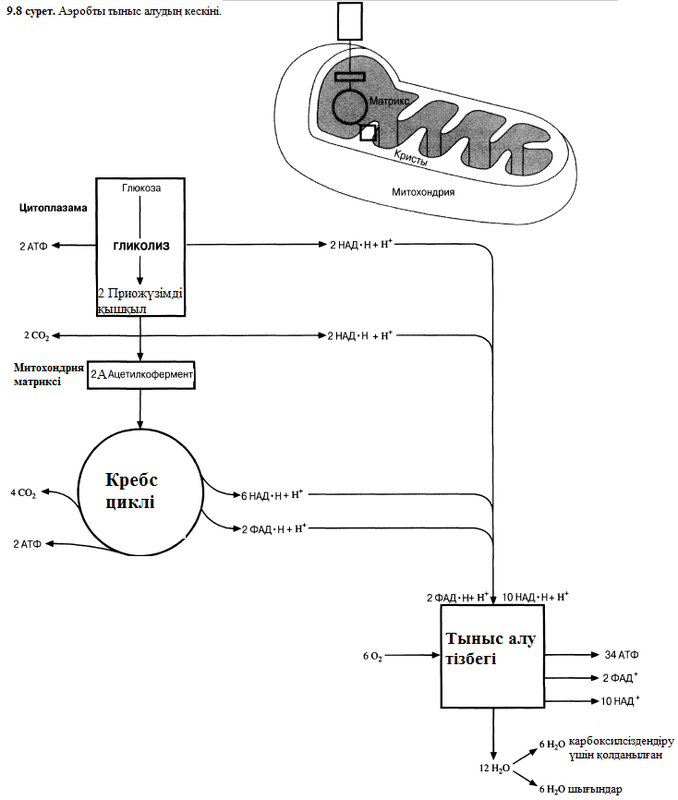
9.3 кестесінде тыныс алу тізбегінің жалпы теңгерімі берілген.
Тыныс алу тізбегінің жинақталған деңгейі мынадай түрі болады:
12Н2 + 6О2 12Н2О+34АТФ
Екі берілген деңгейлерді төменде біріктіреміз, 1 және 2:
Тыныс алу тізбегінің жинақталған деңгейі мынадай түрі болады:
12Н2 + 6О2 12Н2О+34АТФ
Екі берілген деңгейлерді төменде біріктіреміз, 1 және 2:
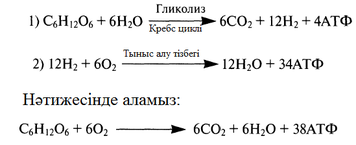
Сонымен, аэробты тыныс алу процесінде қышқылдандырылған глюкозаның әр молекуласына АТФ – тың 38 молекуласы түзіледі.
Аэробты тыныс алу процесінің ортақ сызбасы 9.8 суретінде көрсетілген.
9.3. Тыныс алу үрдісіндегі оттектің қандай рөл атқарады?
Майлы қышқылдардың қышқылдануы
Тыныс алу субстраты ретінде липидтер қолданылған кезде олар алдымен глицеролға және майлы қышқылдарға гидролизденеді, содан соң майлы қышқыл молекуласынан ретті түрде екі көміртекті бөлшектер айырылады, сондықтан да әр кезеңде бұл ұзын молекула екі көміртек атомын қысқарады. Екі көміртекті ацетильді топ А коферментімен бірігеді және түзілген ацетил – КоА, әдеттегіше Кребч цикліне түседі. Майлы қышқылдың әр молекуласынан энергияның көп мөлшері бөлінеді: стеаринді қышқыл қышқылданғанда, мысалы, АТФ – тің шығуы 147 молекуласы құрайды. Сондықтан да, майлы ышқылдар – энергияның басты көзі екендігі таңқаларлық емес. жүрек бұлшықеттерінің (тыныштықта), бүйректердің немесе бауырдың қарапайым энергиялық шығындарының, шамамен, жартысы, майлы қышқылдардың қышқылдануы есебінен жабылады.
9.3.6. Анаэробты тыныс алу
Анаэробты тыныс алуды жиі ашу деп атайды. Көптеген микроағзалар өздерінің АТФ – інің көп бөлігін анаэробты тыныс алу есебінен алады. Кейбір бактериялар үшін оттегі, қалыпты атмосферадағы мөлшерде болғанның өзінде, мүлдем жойғыш, сондықтан да олар оттегі жоқ жерде өмір сүруге мәжбүр. Мұндай ағзаларды облигатты анаэробтар деп атайды (мысал – Ceostridium tetani, сіреспенің қоздырғышы).
Оттексіз де, оттек болғанда да өмір сүре алатын басқа да ағзалар белгілі, мысалы, ашытқылар және ішек жолдарының паразиттер (ленталық құрттар және т.б.). оларды факультативті анаэробтар деп аталады: қажет болғанда олар анаэробты тыныс алуға көшеді, алайда оттек бар кезде аэробты жолды қолданады.
Уақытша оттек жетіспеушілігін сезінетін кейбір жасушалар (жекеше алғанда, бұлшықет жасушалары) да анаэробты тыныс алу қабілетіне ие.
Анаэробты тыныс алудың да бірінші фазасы гликолиз болып табылады. Ол нәтижесінде глюкозаның әр молекуласына пирожүзімді қышқылдың екі молекуласын, АТФ – тің екі молекуласын және қалпына келтірілген НАД – тың екі молекуласын береді (9.1 кестесіне қараңыздар). Аэробты тыныс алуда НАД – қа қосылған сутек, энергияны бөлумен жүретін реакциялар қатарынан соң, ең соңында оттекте тасымалданады және суға дейін қышқылданады. Анаэробты тыныс алуда бұл мүмкін емес болады, қанша дегенмен оттек жоқ. Мұның орнына сутек қайтадан приожүзімді қышқылға қосылады, сондықтан глюкоза молекуласында қамалған энергияның бір бөлігі өндірілмейді (ашудың соңғы өнімінде қалады). Төменде біз бұның саңырауқұлақтрда және жануар жасушасында қалай жүретіндігін нақтырақ қарастырамыз.
Саңырауқұлақтардағы анаэробты тыныс алу, мысалы, ашытқылардағы
Аэробты тыныс алу процесінің ортақ сызбасы 9.8 суретінде көрсетілген.
9.3. Тыныс алу үрдісіндегі оттектің қандай рөл атқарады?
Майлы қышқылдардың қышқылдануы
Тыныс алу субстраты ретінде липидтер қолданылған кезде олар алдымен глицеролға және майлы қышқылдарға гидролизденеді, содан соң майлы қышқыл молекуласынан ретті түрде екі көміртекті бөлшектер айырылады, сондықтан да әр кезеңде бұл ұзын молекула екі көміртек атомын қысқарады. Екі көміртекті ацетильді топ А коферментімен бірігеді және түзілген ацетил – КоА, әдеттегіше Кребч цикліне түседі. Майлы қышқылдың әр молекуласынан энергияның көп мөлшері бөлінеді: стеаринді қышқыл қышқылданғанда, мысалы, АТФ – тің шығуы 147 молекуласы құрайды. Сондықтан да, майлы ышқылдар – энергияның басты көзі екендігі таңқаларлық емес. жүрек бұлшықеттерінің (тыныштықта), бүйректердің немесе бауырдың қарапайым энергиялық шығындарының, шамамен, жартысы, майлы қышқылдардың қышқылдануы есебінен жабылады.
9.3.6. Анаэробты тыныс алу
Анаэробты тыныс алуды жиі ашу деп атайды. Көптеген микроағзалар өздерінің АТФ – інің көп бөлігін анаэробты тыныс алу есебінен алады. Кейбір бактериялар үшін оттегі, қалыпты атмосферадағы мөлшерде болғанның өзінде, мүлдем жойғыш, сондықтан да олар оттегі жоқ жерде өмір сүруге мәжбүр. Мұндай ағзаларды облигатты анаэробтар деп атайды (мысал – Ceostridium tetani, сіреспенің қоздырғышы).
Оттексіз де, оттек болғанда да өмір сүре алатын басқа да ағзалар белгілі, мысалы, ашытқылар және ішек жолдарының паразиттер (ленталық құрттар және т.б.). оларды факультативті анаэробтар деп аталады: қажет болғанда олар анаэробты тыныс алуға көшеді, алайда оттек бар кезде аэробты жолды қолданады.
Уақытша оттек жетіспеушілігін сезінетін кейбір жасушалар (жекеше алғанда, бұлшықет жасушалары) да анаэробты тыныс алу қабілетіне ие.
Анаэробты тыныс алудың да бірінші фазасы гликолиз болып табылады. Ол нәтижесінде глюкозаның әр молекуласына пирожүзімді қышқылдың екі молекуласын, АТФ – тің екі молекуласын және қалпына келтірілген НАД – тың екі молекуласын береді (9.1 кестесіне қараңыздар). Аэробты тыныс алуда НАД – қа қосылған сутек, энергияны бөлумен жүретін реакциялар қатарынан соң, ең соңында оттекте тасымалданады және суға дейін қышқылданады. Анаэробты тыныс алуда бұл мүмкін емес болады, қанша дегенмен оттек жоқ. Мұның орнына сутек қайтадан приожүзімді қышқылға қосылады, сондықтан глюкоза молекуласында қамалған энергияның бір бөлігі өндірілмейді (ашудың соңғы өнімінде қалады). Төменде біз бұның саңырауқұлақтрда және жануар жасушасында қалай жүретіндігін нақтырақ қарастырамыз.
Саңырауқұлақтардағы анаэробты тыныс алу, мысалы, ашытқылардағы
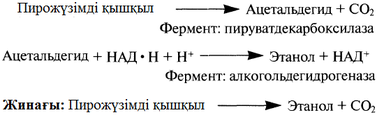
Мұнда спирттік ашу атын алын жүретін процесінің соңғы кезеңдері көрсетілген. Спирттік ашуда АТФ тек оның ерте кезеңдерінде түзіледі – глбкозаның пирожүзімді қышқылға дейін ыдырауы кезінде. Спирттік ашу сыра, шарап және басқа спирттік шырындарды өндіруде қолданылады. Нан өнімдерін өндіруде ашытқымен спирттік ашу процесінде бөлінетін – СО2 – ні қолданады – бұл газдың көпіршіктері қамырды көтерілуге мәжбүрлейді. Спирттік ашудың соңғы өнімі – этанол – құрамында жеткілікті түрде көп энергия бар (Бразилияда, мысалы, автокөліктер жүретін газохол содан жасалады). Алайда, оттегі жоқ болса,этанолдан энергияны бөлуге болмайды.
Спирттік ашудағы АТФ – тің жалпы шығуы, бір глюкоза молекуласына екі АТФ молекуласын құрайды.
Жануар жасушаларындағы анаэробты тыныс алу, мысалы, бұлшықет ұлпаларындағы
Спирттік ашудағы АТФ – тің жалпы шығуы, бір глюкоза молекуласына екі АТФ молекуласын құрайды.
Жануар жасушаларындағы анаэробты тыныс алу, мысалы, бұлшықет ұлпаларындағы
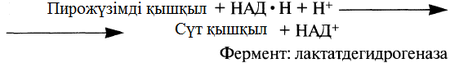
Спирттік ашумен салыстырғанда, сүттіқышқылды ашуда не СО2, не этанол түзілмейді. Бұл жағдайдағы соңғы өнім бұлшықеттегі жиналуы шашршау сезімін, ал кейде құрысуды тудыратын сүт қышқылы болып табылады.
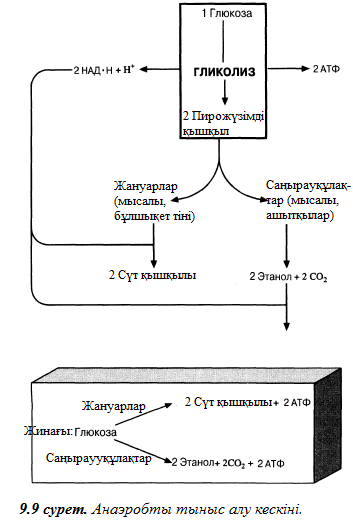
Күшейтілген бұлшықет жұмысында туатын қышқылдық қарыздар туралы, біз 9.3.8. бөлімінде айтатын боламыз.
Сүтті қышқылды ашу кезінде, спирттік ашудағы сияқты, бір глюкоза молекуласына екі АТФ молекуласы түзіледі. Оның соңғы өнымы- сүт қышқылында – тағы көп энергия сақталады.
Анаэробты тыныс алудың ортақ нүктесі 9.9 суретінде көрсетілген.
9.3.7. Аэробты және анаэробты тыныс алу кезіндегі энергияның айналуының әсерлілігі
Аэробты тыныс алу
Аэробты тыныс алу кезінде әр қышқылданған глюкоза молекуласына 38АТФ молекуласы түзіледі.
Сүтті қышқылды ашу кезінде, спирттік ашудағы сияқты, бір глюкоза молекуласына екі АТФ молекуласы түзіледі. Оның соңғы өнымы- сүт қышқылында – тағы көп энергия сақталады.
Анаэробты тыныс алудың ортақ нүктесі 9.9 суретінде көрсетілген.
9.3.7. Аэробты және анаэробты тыныс алу кезіндегі энергияның айналуының әсерлілігі
Аэробты тыныс алу
Аэробты тыныс алу кезінде әр қышқылданған глюкоза молекуласына 38АТФ молекуласы түзіледі.

Глюкозаның толық қышқылдануы кезінде босатылатын энергияның жалпы мөлшері 7 мольге 2880кДж құрайды.
АТФ – тің 1 молінде 30,6 кДЖ құлыпталған
АТФ – тың 30, 6 молінде 30,6 × 38 =1162,8 кДж құлыпталған
Осылайша, аэробты тыныс алудағы энергияның айналуы 1162,8 / 2280 = 40,4 % құрайды.
Анаэробты тыныс алу
Глюкозадан ол этанолға айналғана кезде бөлінетін энергияның жалпы мөлшері 1 мольге 210 кДж – ді құрайды.
АТФ – тың 2 молінде 2 × 30,6 = 61,2 кДж құлыпталған
Сондықтан да, спирттік ашудағы энергияның айналуының әсерлілігі 61,2 / 21, = 29, 1% - ды құрайды.
Глюкозаның ол сүт қышқылына айналған кезде бөлінетін энергияның жалпы мөлшері 1 мольге 150 кДж – ді құрайды.
Осылайша, қышқылды сүт ферменттеу кезіндегі энергияның айналу эффекттілігі 61,2/150 = 40,8%.
Көрсетілген цифрлар әр системадағы энергияның айналу эффекттілігінің бензинді және булы системаларарға қарағанда, тиімдірек екенін көрсетеді. Аэробты тыныс алу арқылы АТФ түрінде сақталған энергия мөлшері (бірінші жағдайда глюкоза молекуласы бір 38 молекула АТФ және екі молекула АТФ) анаэробтыға қарағанда, 19 есе артық болып табылады. Осы тұрғыдан бастап, анаэробтыдан аэробты тыныс әлдеқайда тиімді болып табылады. Бұл энергия кезінде анаэробты тыныс айтарлықтай бөлігі этанол немесе сүт қышқылы «қамалып» екендігіне байланысты. Тиімсіз процесі - энергетика, демек энергетикалық мағынасында алкоголь ашытуды этанол қамтылған және ашытқы үшін тұрақты қол жетімді болып қалады. Сүт қышқылы қалпында болмауы да мүмкін, себебі оттегі көрсетілгенде, кейінірек энергиясы өте көп болуы мүмкін. Оттегі бар сүт қышқылы, бауырда пировиноградной қышқылымен айналады. Соңғысы содан Кребс циклы беріледі және толық қосымша ATФ молекулалардың үлкен мөлшерін тудырады, c02 және H2 O дейін тотығады. Тағы бір жолы бар - пировиноградной қышқылынан АТФ энергиясын қайтадан гликолиза топтастыру болып табылатын глюкоза процесінде құрылуы мүмкін.
9.3.8. Оттегілі қарыздар және бұлшық жүктемені дереу әсері
АТФ жасушаларында қор өте төмен болып табылады. Әдетте ол органға жұмсалады, шамада осы акциялар бойынша өтейді, бірақ қалған өсті бұлшық жұмысқа өткір көшуге, ол бұл жаңа мемлекет реттеу үшін біраз уақыт кетеді. Әлі жеткілікті аэробты тыныс алу қарқындылығы кемуге дейін бұлшық қажетті электр энергиясымен қамтамасыз ете тетіктері бар. Осындай бір тетігі - анаэробты тыныс. Ол анаэробты тыныс орнына оның балама қарағанда, аэробтық қосымша екенін түсіну маңызды.
АТФ – тің 1 молінде 30,6 кДЖ құлыпталған
АТФ – тың 30, 6 молінде 30,6 × 38 =1162,8 кДж құлыпталған
Осылайша, аэробты тыныс алудағы энергияның айналуы 1162,8 / 2280 = 40,4 % құрайды.
Анаэробты тыныс алу
- Ашытқылар (спирттік ашу). Спирттік ашуда глюкозаның әр молекуласына АТФ – тың екі молекуласы түзіледі.
Глюкозадан ол этанолға айналғана кезде бөлінетін энергияның жалпы мөлшері 1 мольге 210 кДж – ді құрайды.
АТФ – тың 2 молінде 2 × 30,6 = 61,2 кДж құлыпталған
Сондықтан да, спирттік ашудағы энергияның айналуының әсерлілігі 61,2 / 21, = 29, 1% - ды құрайды.
- Бұлшықеттер (сүтті қышқылды ашу). Сүтті қышқылды ашу кезінде глюкозаның әр молекуласына АТФ – тың екі молекуласы түзіледі.
Глюкозаның ол сүт қышқылына айналған кезде бөлінетін энергияның жалпы мөлшері 1 мольге 150 кДж – ді құрайды.
Осылайша, қышқылды сүт ферменттеу кезіндегі энергияның айналу эффекттілігі 61,2/150 = 40,8%.
Көрсетілген цифрлар әр системадағы энергияның айналу эффекттілігінің бензинді және булы системаларарға қарағанда, тиімдірек екенін көрсетеді. Аэробты тыныс алу арқылы АТФ түрінде сақталған энергия мөлшері (бірінші жағдайда глюкоза молекуласы бір 38 молекула АТФ және екі молекула АТФ) анаэробтыға қарағанда, 19 есе артық болып табылады. Осы тұрғыдан бастап, анаэробтыдан аэробты тыныс әлдеқайда тиімді болып табылады. Бұл энергия кезінде анаэробты тыныс айтарлықтай бөлігі этанол немесе сүт қышқылы «қамалып» екендігіне байланысты. Тиімсіз процесі - энергетика, демек энергетикалық мағынасында алкоголь ашытуды этанол қамтылған және ашытқы үшін тұрақты қол жетімді болып қалады. Сүт қышқылы қалпында болмауы да мүмкін, себебі оттегі көрсетілгенде, кейінірек энергиясы өте көп болуы мүмкін. Оттегі бар сүт қышқылы, бауырда пировиноградной қышқылымен айналады. Соңғысы содан Кребс циклы беріледі және толық қосымша ATФ молекулалардың үлкен мөлшерін тудырады, c02 және H2 O дейін тотығады. Тағы бір жолы бар - пировиноградной қышқылынан АТФ энергиясын қайтадан гликолиза топтастыру болып табылатын глюкоза процесінде құрылуы мүмкін.
9.3.8. Оттегілі қарыздар және бұлшық жүктемені дереу әсері
АТФ жасушаларында қор өте төмен болып табылады. Әдетте ол органға жұмсалады, шамада осы акциялар бойынша өтейді, бірақ қалған өсті бұлшық жұмысқа өткір көшуге, ол бұл жаңа мемлекет реттеу үшін біраз уақыт кетеді. Әлі жеткілікті аэробты тыныс алу қарқындылығы кемуге дейін бұлшық қажетті электр энергиясымен қамтамасыз ете тетіктері бар. Осындай бір тетігі - анаэробты тыныс. Ол анаэробты тыныс орнына оның балама қарағанда, аэробтық қосымша екенін түсіну маңызды.
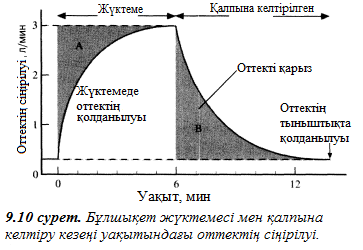
9.10. графигінде көрсетілген бұлшықетті қысымнан және сол мезетте қоюынан оттегінің қалай жұтылатынын байқауға болады. Аэробты тыныс алу арқылы энергия үшін сұранысты қанағаттандыру мақсатында минутына оттегі денесі 3 литр тұтынуы керек. Бұл мысалда бұл бұлшық жұмыстың басынан бастап тек алты минут қол жеткізуге болады (см қараңыз). Оттегі борыш Граф саласындағы ұсынылған (талап етіледі оттегі мөлшері, бірақ тыныс үшін сырттан орган келiп түспесе) алғашқы алты минут деп төменде сипатталған түрлі тетіктері бар энергия қажетті мөлшерін қамтамасыз ете алады.
9.4. Бірден бұлшық жұмыс басталғаннан кейін оттегі сіңіру деңгейі артады. Қалай ұяшықтарға ортадан оттегін жеткізу, бұлшық жүктемесі артпай?
Оттегі қоры
Денеге оттегін аударғым кейбір резервтері бар. Әдеттегіден көп, ол жеңіл және гемоглобин дене сұйықтары оттегін алу үшін болады. Оттегі сақтауға арналған бұлшық белок миоглобине ретінде қызмет етеді. Ол гемоглобин (сур. 3.36) үшін табиғатта өте ұқсас және гемоглобин, сол сияқты, қайтымды оттегін байланыстыруға қабілетті. Алайда, бұл оттегі ең гемоглобин береді кейін оттегі деңгейі, яғни, өте төмен болған кезде ғана оттегі шығарады. Сондықтан, миоглобине сақталған оттегі, тек қажет болған жағдайда ең шеткі пайдаланылатын.
Фосфокреатин жүйесі
Бұлшық қамтылған ATФ барынша бұлшық жүктеме кезінде шамамен үш секунд артық емес жеткілікті болады. Фосфокреатин - энергия фосфат тобының бөлшектенуінің бойынша босатылатын, оның бұл басқа мәселе. Бұл АДФ және Пи АТФ синтезі үшін жеткілікті энергия. Бұлшық phosphocreatine қор ATФ қорын қарағанда 2-4 есе жоғары болып табылады, және қажет болған жағдайда, фосфокреатин энергетикалық тез АТФ берілуі мүмкін. АТФ және 8-10 секунд ішінде ең көп бұлшық жүктемені теңселмей phosphocreatine энергияның Бірыңғай қор.
Анаэробты тыныс алу. Сүтті-гликогенді қышқыл жүйесі
Глюкоза молекуласы бір анаэробты тыныс тек екі молекула АТФ және аэробты шығарылады қарамастан - 38 молекулаларды, бірақ бұрынғы жағдайда, АТФ синтезі 2,5 есе жылдам аэробты оның барысында (анаэробты тыныс уақыт сол мерзiмге АТФ бес молекула береді Ол екі) береді. Анаэробты тыныс, сондықтан тез энергияны болады. Глюкоза көзі осылайша бұлшық сақталады гликоген табылады. оған алынған Энергия90 секундқа кезде максималды бұлшық қызметін жеткілікті.
Регулярлы қысымда бұның бәрі жақсы жұмыс атұарады.
Сондықтан біз бірақ тек қысқа уақытқа, жүйелік phosphocreatine және анаэробты тыныс тез энергиясын жеткізуге жатқанын көріп отырмыз. Аэробты жүйесі кезде жеткілікті респираторлық субстрат белгісіз мерзімге энергия көзі ретінде қызмет көрсете алады. Осындай қысқа қашықтықты іске қосылған немесе кір тасын көтеру сияқты бұлшық қызметінің қысқа мерзімді және күрт арттыру үшін әзірленген осы спорт, жылы, энергетикалық жеткізілімдер негізінен фосфокреатин жүйесі. Әйелдер 200 метрге анаэробты тыныс энергия қосымша көзі ретінде қызмет ете алады. Уақыт осындай теннис, сквош немесе футбол, барлық дерлік энергия ретінде ойындар жүйесі арқылы кернеу берілетін шектеу, ал 400 метр, ол қазірдің өзінде, энергияны барынша жеткізеді. Спорт түрлерін бастысы - төзімділік, мұндай марафон, жүгіру немесе кросс шаңғы тебу жарысынан ретінде аэробты тыныс толықтай дерлік тәуелді.
Бұлшық жұмыс соңында (сур. 9.10) кезде, оттегі тұтыну дереу тыныштық деңгейі тән (0,25 л / мин) оралу жоқ. Адам қатты дем үшін біраз уақыт жалғастыруда қалпына келтіру кезеңде. Бұл оттегі пайдаланылады:
Осы реакцияда 1 моль АТФ – ке 30,6 кДж көлемінде бос энергия жұмсалады.
АДФпен фосфоттан қайтадан АТФ синтезделуі мүмкін. Бірақ, қайтадан АТФ – ты түзу үшін 1 моль – ге 30,6 кДж көлемінде энергия қажет болады. Бұл реакцияда су бөлінеді. Мұндай реакцияны – конденсация реакциясы дейді. АДФ – ке фосфаттың қосылуын фосфорлану реакциясы деп атайды. Жоғарыда келтірілген ескі теңдеуді біріктіруге болады:
9.3.9. Өндіріс мақсаты үшін ашыту үрдісінің қолданылуы
Әртүрлі ашыту өндірістірінде ашыту үрдісі микроағзалардың көмегімен жүзеге асады. Бұл үрдіс заманауи шаруашылықта үлкен рөл атқарады. Бұл микроағзаларды арнайы контейнерлерде өсіреді. Мұндай контейнерлерде ферменттер немесе биореакторлар дейді. Бұл үрдіс жайында 12 бөлімде айтатын боламыз.
Тәжірибе 9.1. Кребс циклінің аралық өнімдерінің бірін қышқылдануын зерттеу
Субстраттан энергияны алудың және оны келесіге қолдану үшін қорғап сақтаудың айрықша тиімді әдісті сол үрдісті катализдеуші ферменттерді қатысуымен бір қатар қайтымды реакцияларға болып табылады. осындай аралық реакцияларының бірі – сутекті ыдырату арқылы кәріптас қышқылын фумарлығы тотықтыру.
Қазірге мәлім біршама заттар бар. Олар сутектің ыдыраған атомдары бойланыстыра алады әрі өз түсін өзгертеді. Мұндай затарға 2,6 – дихлорфеномендофенол (ДХФИФ) болып табылады. оның тотыққан пішіні қоб көк түске боялады, ал қалпына келтірілгені түссіз болады.
Егер ДХФИФ – тың тотыққан пішінін сіріндісімен араластырғанда түссізденеді. Бұған себеп ретінде сутек атомдарының кәріптас қышқылынан алынып оғанқосылуы болып табылады. қоспаға кәріптас қышқылын қосамыз. Егер ендігіде түссізденудің жылдамдығы тез артса, онда бұл біздің болжамымызда ДХФИФ сутек атомдарының акцепторы ретінде екенін толықтырады. Ол кәріптас қышқылынан бөлініп шығады.
Тірі ағзалардағы реакциялар ферменттердің катализімен өтетіні анық. Өйткені, тотығу үрдісі ортада тиісті фермент болса, сонда ғана жүреді. Кәріптас қышқылының тотығу реакциясын сунцинатдегидрогеназа ферментінің катализі әсер етеді. Әрі бұл ферменттің кәріптас қышқылының құрамында бар екен тәжірибелер арқылы көреміз анықталған. Біздің тәжірибеде фермент көзі болып маш тұқымының өсіп келе жатқан митохондрияның суспензиясы болады.
Митохондрияларды барынша тез шығарып тұру қажет, өйткені жасушалар жоюлуынан кейін метаболизм ұзаққа созылмайды.
Тәжірибе 2 кезеңге бөлінеді: 1) қажетті ферментті шығару; 2) кәріптас қышқылының тотығуы үішн қышқыл қолдану. ДХФИФ индикатор ретінде реакция жүретінін әлде жүрмейтінің анықтайды. 1 – ші тәжірибе кезеңін қажетті (яғни, фермент сіріндісін дайындау үшін) маш тұқымын 1 сағат шамасында жұмысты бастамас бүрын тоңазтқышқы қою.
Құрал – жабдықтар
4 центрифужды түтіктер (15 мл – ден)
2 шыны таяқшалар
2 дәйектелген тамшуыр 10 мл – ден
Сыйымдылығы 1 л, болатын 2 зертханалық стақан (жарлы этилен болған қолайлы)
Мұз
Тұз
Маш тұқымы
Штатив түтіктер
1 дәйектелген тамшуыр 1 мл – ден
Таймер
Ерітінділер (тәжірибе соңында ерітіндіні дайындауға қажетті мәлімет берілген)
Буферлі сахароза ерітіндісі
Буферлі сахароза ерітіндісі + кәріптас қышқылының ерітіндісі
0,1 % ДХФИФ ерітіндісі (буферлі сахароза ерітіндісінде дайындалады)
Дистелденген су
Жұмыс барысы:
Буферлі сахароза ерітіндісі (100 мл)
Натрий гидрофосфаты (Na2HPO4)
Калий дигидрофосфаты (КH2PO4)
Сахароза
Магний сульфаты
Буферлі сахароза ерітіндісі + кәріптас қышқылы (100 мл)
Буферлі сахароза ерітіндісі. Жоғарыда дайындау жайында айтылған
Кәріптас қышқыл
Натрий гидрокарбонаты
Тәжірибенің 2 кезеңі үшін буферлі сахароза ерітіндісін жеткілікті етіп дайындап қойған ыңғайлы (ерітінді дистилденген суда дайынжалады). Содан кейін ерітіндіні екіге тең бөліп, біреуіне кәріптас қышқылы мен натрий гидрокарбонатын қосу керек (тиісті концтрациясында).
Буферлі сахароза ерітіндісіне кәріптас қышқылы мен натрий гидрокарбонатын қосқанда, сұйықтық қайнайды.көмірқышқыл газын шығрау үшін ерітіндіні жақсылап шайқау қажет. Өйткені, оның құрамда болуы экспериментке әсер ету мүмкін.
ДХФИФ ерітіндісі. 10 мл мөлшеріндегі буферленген сахароза ерітіндісіне (эксперименттің 2 бөлігі үшін кәріптас қышқылынсыз) 0,1 г мөлшердегі дихлорфонолиндофенолды ерітіңіз. Бұл заттың еріткіштігі өте төмен, сондықтан суспензиянымұқият араластырғаннан кейін оны сүзу қажет.
9.3.10. Митохондриялар
Митохондриялар эукариот жасушалардың бәрінде бар. Бұл органелалалар – жасушаның аэробты тыныс белсенілігінің басты жері. 1850 жылы алғашында митохондриялар түйіршік түрінде табылған.
9.4. Бірден бұлшық жұмыс басталғаннан кейін оттегі сіңіру деңгейі артады. Қалай ұяшықтарға ортадан оттегін жеткізу, бұлшық жүктемесі артпай?
Оттегі қоры
Денеге оттегін аударғым кейбір резервтері бар. Әдеттегіден көп, ол жеңіл және гемоглобин дене сұйықтары оттегін алу үшін болады. Оттегі сақтауға арналған бұлшық белок миоглобине ретінде қызмет етеді. Ол гемоглобин (сур. 3.36) үшін табиғатта өте ұқсас және гемоглобин, сол сияқты, қайтымды оттегін байланыстыруға қабілетті. Алайда, бұл оттегі ең гемоглобин береді кейін оттегі деңгейі, яғни, өте төмен болған кезде ғана оттегі шығарады. Сондықтан, миоглобине сақталған оттегі, тек қажет болған жағдайда ең шеткі пайдаланылатын.
Фосфокреатин жүйесі
Бұлшық қамтылған ATФ барынша бұлшық жүктеме кезінде шамамен үш секунд артық емес жеткілікті болады. Фосфокреатин - энергия фосфат тобының бөлшектенуінің бойынша босатылатын, оның бұл басқа мәселе. Бұл АДФ және Пи АТФ синтезі үшін жеткілікті энергия. Бұлшық phosphocreatine қор ATФ қорын қарағанда 2-4 есе жоғары болып табылады, және қажет болған жағдайда, фосфокреатин энергетикалық тез АТФ берілуі мүмкін. АТФ және 8-10 секунд ішінде ең көп бұлшық жүктемені теңселмей phosphocreatine энергияның Бірыңғай қор.
Анаэробты тыныс алу. Сүтті-гликогенді қышқыл жүйесі
Глюкоза молекуласы бір анаэробты тыныс тек екі молекула АТФ және аэробты шығарылады қарамастан - 38 молекулаларды, бірақ бұрынғы жағдайда, АТФ синтезі 2,5 есе жылдам аэробты оның барысында (анаэробты тыныс уақыт сол мерзiмге АТФ бес молекула береді Ол екі) береді. Анаэробты тыныс, сондықтан тез энергияны болады. Глюкоза көзі осылайша бұлшық сақталады гликоген табылады. оған алынған Энергия90 секундқа кезде максималды бұлшық қызметін жеткілікті.
Регулярлы қысымда бұның бәрі жақсы жұмыс атұарады.
Сондықтан біз бірақ тек қысқа уақытқа, жүйелік phosphocreatine және анаэробты тыныс тез энергиясын жеткізуге жатқанын көріп отырмыз. Аэробты жүйесі кезде жеткілікті респираторлық субстрат белгісіз мерзімге энергия көзі ретінде қызмет көрсете алады. Осындай қысқа қашықтықты іске қосылған немесе кір тасын көтеру сияқты бұлшық қызметінің қысқа мерзімді және күрт арттыру үшін әзірленген осы спорт, жылы, энергетикалық жеткізілімдер негізінен фосфокреатин жүйесі. Әйелдер 200 метрге анаэробты тыныс энергия қосымша көзі ретінде қызмет ете алады. Уақыт осындай теннис, сквош немесе футбол, барлық дерлік энергия ретінде ойындар жүйесі арқылы кернеу берілетін шектеу, ал 400 метр, ол қазірдің өзінде, энергияны барынша жеткізеді. Спорт түрлерін бастысы - төзімділік, мұндай марафон, жүгіру немесе кросс шаңғы тебу жарысынан ретінде аэробты тыныс толықтай дерлік тәуелді.
Бұлшық жұмыс соңында (сур. 9.10) кезде, оттегі тұтыну дереу тыныштық деңгейі тән (0,25 л / мин) оралу жоқ. Адам қатты дем үшін біраз уақыт жалғастыруда қалпына келтіру кезеңде. Бұл оттегі пайдаланылады:
- Ағзадағы оттегі жабдықтау толықтыру үшін, бұл тіндердің сұйықтар, миоглобине және гемоглобин жарық оның қалыпты деңгейін қалпына келтіру үшін, болып табылады.
- Фосфокреатин регенерациясы үшiн - бұлшық креатин жұмыстың соңында қайтадан фосфат мән; Осы аэробты тыныс үшін энергияны қамтамасыз етеді.
- Орган және phosphocreatine қалпына келтіру оттегі жабдықтау толықтыру тез жүреді; (Сур. 9.10) жаңарту бірінші минутынан сәйкес қисық тік бөлігін төмен бара дәлел. Баяу қалпына келтіру (қисық бөлігін көлбеу) - анаэробты тыныс алу бойынша жинақталған бұлшық жылғы сүт қышқылының жою, бар кезде осы кезең. Сүт қышқылы қанға шығарды және ол қышқылы пировиноградной дейін тотығады НАД азаяды бауыр, бұлшық үшін түскен тасымалданады. Ылу алмасу шарап қышқылы бөлігі Кребс циклінің арқылы аэробты қалыпты жолмен барады және АТФ қалыптастыру нәтижесінде тотығу өтеді. Бұл ATP содан гликолиз қарап тұрғанын процесс арқылы кері глюкоза ішіне пировиноградной қышқылы (шамамен 75%), қалған түрлендіру үшін пайдаланылады. Жүрек бұлшықетінің ауыр жүктеме астында сүт қышқылы, сондай-ақ пировиноградной қышқылы айналдыруға болады НАД тотығады, және бұл процесс, мұнда энергияның қосымша көзі болып табылады.
Осы реакцияда 1 моль АТФ – ке 30,6 кДж көлемінде бос энергия жұмсалады.
АДФпен фосфоттан қайтадан АТФ синтезделуі мүмкін. Бірақ, қайтадан АТФ – ты түзу үшін 1 моль – ге 30,6 кДж көлемінде энергия қажет болады. Бұл реакцияда су бөлінеді. Мұндай реакцияны – конденсация реакциясы дейді. АДФ – ке фосфаттың қосылуын фосфорлану реакциясы деп атайды. Жоғарыда келтірілген ескі теңдеуді біріктіруге болады:
9.3.9. Өндіріс мақсаты үшін ашыту үрдісінің қолданылуы
Әртүрлі ашыту өндірістірінде ашыту үрдісі микроағзалардың көмегімен жүзеге асады. Бұл үрдіс заманауи шаруашылықта үлкен рөл атқарады. Бұл микроағзаларды арнайы контейнерлерде өсіреді. Мұндай контейнерлерде ферменттер немесе биореакторлар дейді. Бұл үрдіс жайында 12 бөлімде айтатын боламыз.
Тәжірибе 9.1. Кребс циклінің аралық өнімдерінің бірін қышқылдануын зерттеу
Субстраттан энергияны алудың және оны келесіге қолдану үшін қорғап сақтаудың айрықша тиімді әдісті сол үрдісті катализдеуші ферменттерді қатысуымен бір қатар қайтымды реакцияларға болып табылады. осындай аралық реакцияларының бірі – сутекті ыдырату арқылы кәріптас қышқылын фумарлығы тотықтыру.
Қазірге мәлім біршама заттар бар. Олар сутектің ыдыраған атомдары бойланыстыра алады әрі өз түсін өзгертеді. Мұндай затарға 2,6 – дихлорфеномендофенол (ДХФИФ) болып табылады. оның тотыққан пішіні қоб көк түске боялады, ал қалпына келтірілгені түссіз болады.
Егер ДХФИФ – тың тотыққан пішінін сіріндісімен араластырғанда түссізденеді. Бұған себеп ретінде сутек атомдарының кәріптас қышқылынан алынып оғанқосылуы болып табылады. қоспаға кәріптас қышқылын қосамыз. Егер ендігіде түссізденудің жылдамдығы тез артса, онда бұл біздің болжамымызда ДХФИФ сутек атомдарының акцепторы ретінде екенін толықтырады. Ол кәріптас қышқылынан бөлініп шығады.
Тірі ағзалардағы реакциялар ферменттердің катализімен өтетіні анық. Өйткені, тотығу үрдісі ортада тиісті фермент болса, сонда ғана жүреді. Кәріптас қышқылының тотығу реакциясын сунцинатдегидрогеназа ферментінің катализі әсер етеді. Әрі бұл ферменттің кәріптас қышқылының құрамында бар екен тәжірибелер арқылы көреміз анықталған. Біздің тәжірибеде фермент көзі болып маш тұқымының өсіп келе жатқан митохондрияның суспензиясы болады.
Митохондрияларды барынша тез шығарып тұру қажет, өйткені жасушалар жоюлуынан кейін метаболизм ұзаққа созылмайды.
Тәжірибе 2 кезеңге бөлінеді: 1) қажетті ферментті шығару; 2) кәріптас қышқылының тотығуы үішн қышқыл қолдану. ДХФИФ индикатор ретінде реакция жүретінін әлде жүрмейтінің анықтайды. 1 – ші тәжірибе кезеңін қажетті (яғни, фермент сіріндісін дайындау үшін) маш тұқымын 1 сағат шамасында жұмысты бастамас бүрын тоңазтқышқы қою.
Құрал – жабдықтар
4 центрифужды түтіктер (15 мл – ден)
2 шыны таяқшалар
2 дәйектелген тамшуыр 10 мл – ден
Сыйымдылығы 1 л, болатын 2 зертханалық стақан (жарлы этилен болған қолайлы)
Мұз
Тұз
Маш тұқымы
Штатив түтіктер
1 дәйектелген тамшуыр 1 мл – ден
Таймер
Ерітінділер (тәжірибе соңында ерітіндіні дайындауға қажетті мәлімет берілген)
Буферлі сахароза ерітіндісі
Буферлі сахароза ерітіндісі + кәріптас қышқылының ерітіндісі
0,1 % ДХФИФ ерітіндісі (буферлі сахароза ерітіндісінде дайындалады)
Дистелденген су
Жұмыс барысы:
- Маш тұқымдарын (24 дана) ылғал мақтаға орналастырып, 3 – 4 күнге қарңғы жерге қойыңыз.
- Мұзды монша жасаңыз: температураны төмендету үшін сыйымдылығы 1 л жартыэтилен стақанына мұз, тұз салыңыз.
- Мұзды моншаға шыны сауытты орналастырып, оған буферлі сахароза ерітіндісі мен 2 центрафужды түтіктерді салыңыз.
- 12 өскен тұқымдардан түп қабықтарын алыңыз.
- Центрафужды түтіктердің әрқайсысына 6 тұқымнан салыңыз
- Кәріптас қышқылы қоалмаған буферлі сахароза ерітіндісін 1 мл мөлшерінде әрбір түтікке салыңыз.
- 7. Салқындатылған шыны тояқшаларының көмегімен тұқымдарды әбден ұсақтаңыз. Жұмыс жасау барысынд түтіктер үнемі мұз салынған стақанда тұру қажет.
- Әрбір центрафужы түтіктерге 10 мл мөлшерінде буферлі сахароза ерітіндісін салыңыз.
- 9. Центрафужды түтіктерді центрифугаға (қарама – қарсы етіп) орналастырыңыз да 3 млн барысында максималды жылдамдықта айналдырыңыз.
- Центрафужы түтіктерді мұз салынған стақанға қайта қойыңыз.
- 15 мл мөлшерде тамшуырмен кез келген бір түтікке дистилденген су тамызыңыз да тегіршік орнала суын белгілеңіз.
- Дестилденген суды құйып бастаңыз және түтіктің белгісіне дейін бетінде қалғитын сұйықтықты центрифужды түтіктен алып құйыңыз.
- Бұл кезеңді асы тез жылдамдықта жасау керек: сол зертханалық түтікке 0,5 мл мөлшерде ДХФИФ ерітіндісін құйыңыз. Қоспаны араластырып, бас бармақпен түтіктің ауызын жауып тұрып, оны төңкеріңіз.
- Ерітіндіні араластыру барысында санап тұрыңыз.
- Ерітінді түсінін 20 млн кейін белгілеңіз.
- Осы экспериментті қайта қайталаңыз. Бұл кезекте кәріптас қышқылы бар буферлі сахароза ерітіндісін қолданыңыз.
- Қызыл фильтрді қойып, 5 минутқа колориметрді қосып, жылытуға қойыңыз.
- Жоғарыда айтылғандай 15 мин мөлшерден бетінде қалқып жүрген сұйықтыққа 0,5 мл мөлшерде ДХФИФ ерітіндісін қосыңыз.
- Ерітіндіні араластырып, уақытты бақылаңыз.
- Түтікті колорометрге орналастырып, құрылғаны 0% өткізгіштікке қойыңыз.
- Колометрдегі көрсеткіштерді 1, 2, 5, 10 және 20 минуттан кейін белшілеп отырыңыз.
- Сол тәжірибені кәріптас қышқылы бар буферлі сахароза ерітіндісімен қайта орындаңыз.
- Өткізу көрсеткішінің (ордината осінде) уақытқа тәуелді график тұрғызыңыз.
- Тәжірибе нәтижелеріне сүйіне отырып қорытынды жасаңыз.
Буферлі сахароза ерітіндісі (100 мл)
Натрий гидрофосфаты (Na2HPO4)
Калий дигидрофосфаты (КH2PO4)
Сахароза
Магний сульфаты
Буферлі сахароза ерітіндісі + кәріптас қышқылы (100 мл)
Буферлі сахароза ерітіндісі. Жоғарыда дайындау жайында айтылған
Кәріптас қышқыл
Натрий гидрокарбонаты
Тәжірибенің 2 кезеңі үшін буферлі сахароза ерітіндісін жеткілікті етіп дайындап қойған ыңғайлы (ерітінді дистилденген суда дайынжалады). Содан кейін ерітіндіні екіге тең бөліп, біреуіне кәріптас қышқылы мен натрий гидрокарбонатын қосу керек (тиісті концтрациясында).
Буферлі сахароза ерітіндісіне кәріптас қышқылы мен натрий гидрокарбонатын қосқанда, сұйықтық қайнайды.көмірқышқыл газын шығрау үшін ерітіндіні жақсылап шайқау қажет. Өйткені, оның құрамда болуы экспериментке әсер ету мүмкін.
ДХФИФ ерітіндісі. 10 мл мөлшеріндегі буферленген сахароза ерітіндісіне (эксперименттің 2 бөлігі үшін кәріптас қышқылынсыз) 0,1 г мөлшердегі дихлорфонолиндофенолды ерітіңіз. Бұл заттың еріткіштігі өте төмен, сондықтан суспензиянымұқият араластырғаннан кейін оны сүзу қажет.
9.3.10. Митохондриялар
Митохондриялар эукариот жасушалардың бәрінде бар. Бұл органелалалар – жасушаның аэробты тыныс белсенілігінің басты жері. 1850 жылы алғашында митохондриялар түйіршік түрінде табылған.
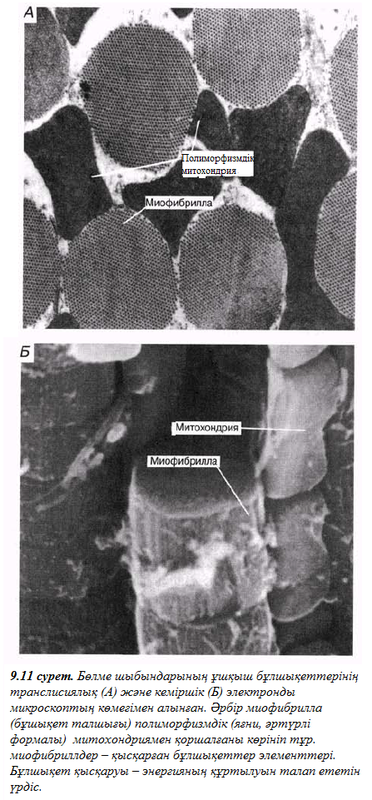
Жасушадағы митохондрия саны өте тұрақсыз; ол ағза түрінен және жасуша табиғатына тәуелді болады. Энергияны өте көп қажет ететін жасушаларда көп митохондрия болады (мысалы, бір бауыр жасушасында олардың шамамен 1000 – ы болады). Белсенділігі төмен жасушаларда , митохондриялар саны да сонша аз болып келеді. Сонымен қатар, митохондрияның өлшемдері мен пішінлері шектен тыс түрлерін өзгертіп отырады. Митохондриялар шиыршықты, дөңгелек , созылған, тостағаншатәрізді және тіпті бұтақтанған да болып келеді, кішкене белсенді жасушаларда олар ірілеу болып келеді.
9.6. Неге соншама үлкен әртүрлі ұзындықтағы митохондриялардың диаметрі салыстырғаны тұрақты?
Митохондриялар өздерінің пішінін өзгертуі қабілетті, ал кейбіреулері ерекше белсенді жасушақатысушыларына ауысуы мүмкін. Мұндай ауысым жасушаға АТФ қажет етілетін жерлерге митохондрияның көп санын жинауға мүмкіндік тудырады. Басқа жағдайларда митохондрия ахуалы тұрақтырақ (мысалы, жәндіктердің ұшқыш бұлшықеттерінде, 9.11 сурет).
Митохондрия құрылымы
9.7. Дем алу барысында цитоплазма мен митохондрия қандай химиялық заттар мен әрекеттеседі? Митохондрияға кіретін 4 затты табуға керек әне митохондриядан цитоплазмаға айналатын затты табу керек.
5 – бөлімде жазылғандай, митохондрия жасушадан таза түріндегі гаомогеннің көмегімен фракция бөліп шығарады. Бұдан соң оларды электорндық имкроскопта зерттеуге болады, ол үшін бірнеше әдістер арқылы кесу жыне болымсыз кереғарлығын салыстыру керек. Әрбір митохондрия қабықшамен қапталған 2 мембранадан мембрана ішкі мембранадан азғантай ғана аралық бөліп тұрады. Мұны ішкі мембрандық кеңістік дейміз. Ішкі мембрана көптеген саңыруқұлақты қалташалардан қтұрады, оларды кристалар деп аталады. Кристалар ішкі мембрананы әлдеқайда жоғары көтеріп тұрады. Соның арқасында белгілі компоненттерге дем алуға орын табылады. Ішкі митохондриялық мембрана арқылы белсенді АДФ пен АТФ құралады. Болымсыз кереғарлық әдісі бойынша құрылымдары боялмайды, ал айналасы боялады, ерекше жағдайларда кез келген бөлімшені ішкі жағынан байқауға болады. Әрбір мұндай бөлімшелер басынан аяғынан және тұтас бөлімінен құралады. Микрофотографиялар элементар бөлшектер митохондриядан матрикске енетініне куәлік етіп отыр, бірақ дұл жасанды деп саналады, құрылғылар сол үрдіске арнайы жабдықталған болатын болса, онда шынымен – ақ олар толығымен мембранада қоршалған болареді.Бөлік бастары АТФ синтезі бөлінуіне жауапты, олард АТФазаферменті болады. Олар АДФ – ты фосфорилдеу реакциясын құрайды. Бөліктердің негізі мембрананың бүкіл денесін құрайды. Негізгі компоненттері дем алу құрамында болады. Митохондриялық матриксте көптеген ферменттер болады. Олар Кребс цикліне және май қышқылдардың жұмысына қатысады. Бұл жерде ДНҚ, РНҚ және 70S – рибосомаларда қатысады.
9.6. Неге соншама үлкен әртүрлі ұзындықтағы митохондриялардың диаметрі салыстырғаны тұрақты?
Митохондриялар өздерінің пішінін өзгертуі қабілетті, ал кейбіреулері ерекше белсенді жасушақатысушыларына ауысуы мүмкін. Мұндай ауысым жасушаға АТФ қажет етілетін жерлерге митохондрияның көп санын жинауға мүмкіндік тудырады. Басқа жағдайларда митохондрия ахуалы тұрақтырақ (мысалы, жәндіктердің ұшқыш бұлшықеттерінде, 9.11 сурет).
Митохондрия құрылымы
9.7. Дем алу барысында цитоплазма мен митохондрия қандай химиялық заттар мен әрекеттеседі? Митохондрияға кіретін 4 затты табуға керек әне митохондриядан цитоплазмаға айналатын затты табу керек.
5 – бөлімде жазылғандай, митохондрия жасушадан таза түріндегі гаомогеннің көмегімен фракция бөліп шығарады. Бұдан соң оларды электорндық имкроскопта зерттеуге болады, ол үшін бірнеше әдістер арқылы кесу жыне болымсыз кереғарлығын салыстыру керек. Әрбір митохондрия қабықшамен қапталған 2 мембранадан мембрана ішкі мембранадан азғантай ғана аралық бөліп тұрады. Мұны ішкі мембрандық кеңістік дейміз. Ішкі мембрана көптеген саңыруқұлақты қалташалардан қтұрады, оларды кристалар деп аталады. Кристалар ішкі мембрананы әлдеқайда жоғары көтеріп тұрады. Соның арқасында белгілі компоненттерге дем алуға орын табылады. Ішкі митохондриялық мембрана арқылы белсенді АДФ пен АТФ құралады. Болымсыз кереғарлық әдісі бойынша құрылымдары боялмайды, ал айналасы боялады, ерекше жағдайларда кез келген бөлімшені ішкі жағынан байқауға болады. Әрбір мұндай бөлімшелер басынан аяғынан және тұтас бөлімінен құралады. Микрофотографиялар элементар бөлшектер митохондриядан матрикске енетініне куәлік етіп отыр, бірақ дұл жасанды деп саналады, құрылғылар сол үрдіске арнайы жабдықталған болатын болса, онда шынымен – ақ олар толығымен мембранада қоршалған болареді.Бөлік бастары АТФ синтезі бөлінуіне жауапты, олард АТФазаферменті болады. Олар АДФ – ты фосфорилдеу реакциясын құрайды. Бөліктердің негізі мембрананың бүкіл денесін құрайды. Негізгі компоненттері дем алу құрамында болады. Митохондриялық матриксте көптеген ферменттер болады. Олар Кребс цикліне және май қышқылдардың жұмысына қатысады. Бұл жерде ДНҚ, РНҚ және 70S – рибосомаларда қатысады.
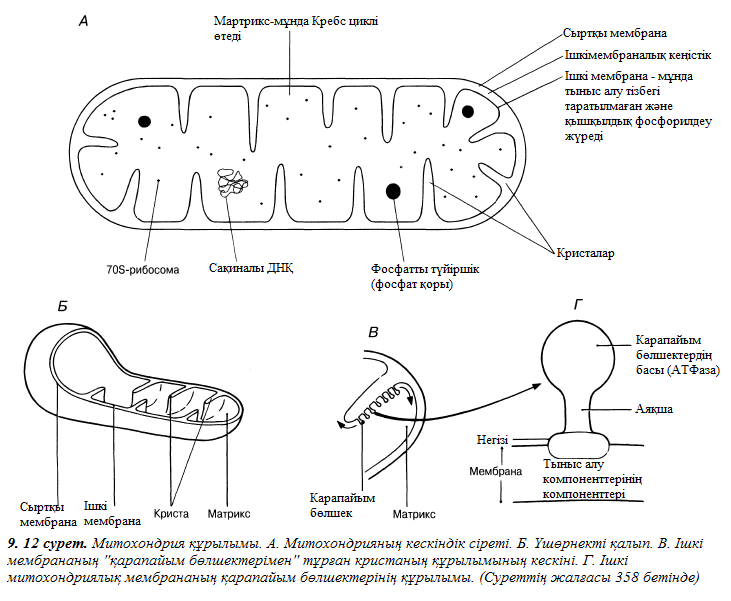
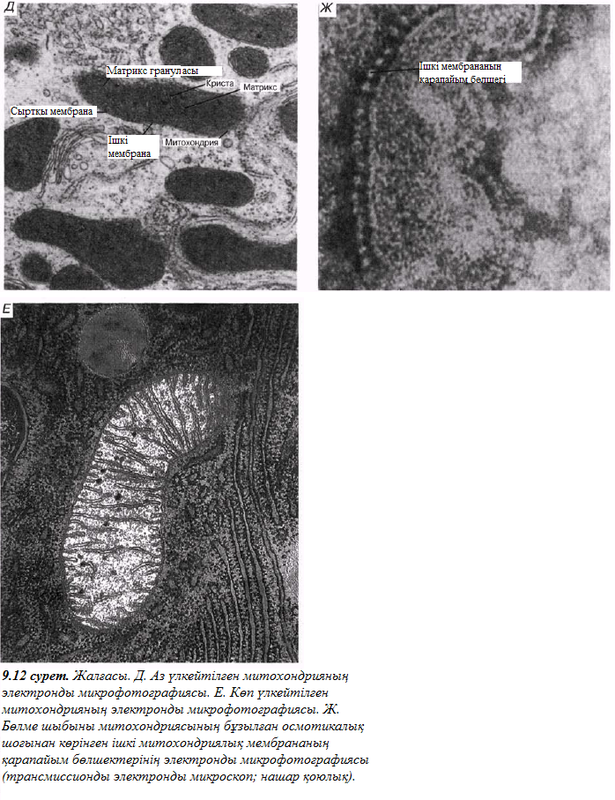
Митохондрия эволюциясы: эндосимбиотикалық болжам
Митохондрия бактерия сияқты ДНҚ сиқинасын құрайды және 70S – рибосоманы құрайды. Бұл жағдай басқа дәлелдермен салыстырғанда, митохондрияның хлоропласттар тәрізді бір кездері еркін бактерия болғаны жөнінде айтады. Кенеттен, бір жасушаа кіріп кеткенде ыңғайлы симбиоз құрған. Бұл жағдайды 2.6.1. бөлімінде көрсетілген.
9.4. Газалмасу
Газалмасу деп демалу газдарымен ағзаның және қоршаған ортаның араласуын айтады. Аэробтарға дем алу үшін сырттан кірген оттегінің маңызы зор. Аэробтар ортасы көміртек қышқылын бөледі және соңғы өнімі дем шығарады. Бұл айналыстың негізін дем алу өнімі деп атайды. Демлу беті аталады. Физикалық жолмен өткен газалмасу диффузия деп аталады. Диффузия тиімді әсер ету үшін дем алу беті бірнеше жағдайлармен қанағаттандырылады.
Митохондрия бактерия сияқты ДНҚ сиқинасын құрайды және 70S – рибосоманы құрайды. Бұл жағдай басқа дәлелдермен салыстырғанда, митохондрияның хлоропласттар тәрізді бір кездері еркін бактерия болғаны жөнінде айтады. Кенеттен, бір жасушаа кіріп кеткенде ыңғайлы симбиоз құрған. Бұл жағдайды 2.6.1. бөлімінде көрсетілген.
9.4. Газалмасу
Газалмасу деп демалу газдарымен ағзаның және қоршаған ортаның араласуын айтады. Аэробтарға дем алу үшін сырттан кірген оттегінің маңызы зор. Аэробтар ортасы көміртек қышқылын бөледі және соңғы өнімі дем шығарады. Бұл айналыстың негізін дем алу өнімі деп атайды. Демлу беті аталады. Физикалық жолмен өткен газалмасу диффузия деп аталады. Диффузия тиімді әсер ету үшін дем алу беті бірнеше жағдайлармен қанағаттандырылады.
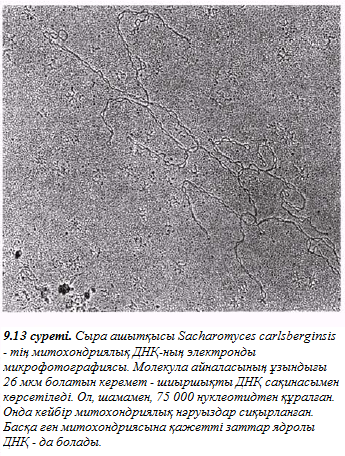
1) оны газ өткізу үшін ыңғайлы болу керек.
2) оны құрайтын қабат жіңішке болуы тиіс, себебі диффузия 1 см-ден аспайтын қашықтықта ғана әсерлі болады;
3) ағзаның қажеттіліктеріне сәйкес, тыныс алу беті арқылы газдардың жеткілікті мөлшері алмасып тұруы үшін, тыныс алу бетінің ауданы үлкен болу керек;
4) қан газдарды тасымалдау ортасы болып қызмет ететін ағзаларда, ол қанмен молырақ қамтамасыз етілуі тиіс (ал кейде бір вентиляцилық меxанизм керек). Бұл оның екі жағының арасында күшті диффузиялық градиентті, яғни концентрацияның үлкен айырмашылығын сақтауға мүмкіндік береді.
Диффузияның тыныс алу беті арқылы максимальді жылдамдыққа қалай жете алатынын түсіну үшін, Фик заңына жүгіну керек. Бұл заңға сәйкес, тыныс алу беті арқылы газдардың диффузиясының жылдамдығы келесі өрнекке пропорционал.(5,9,8 бөлімдерін қарау керек, ол жерде мембрана арқылы диффузия туралы айтылады);
2) оны құрайтын қабат жіңішке болуы тиіс, себебі диффузия 1 см-ден аспайтын қашықтықта ғана әсерлі болады;
3) ағзаның қажеттіліктеріне сәйкес, тыныс алу беті арқылы газдардың жеткілікті мөлшері алмасып тұруы үшін, тыныс алу бетінің ауданы үлкен болу керек;
4) қан газдарды тасымалдау ортасы болып қызмет ететін ағзаларда, ол қанмен молырақ қамтамасыз етілуі тиіс (ал кейде бір вентиляцилық меxанизм керек). Бұл оның екі жағының арасында күшті диффузиялық градиентті, яғни концентрацияның үлкен айырмашылығын сақтауға мүмкіндік береді.
Диффузияның тыныс алу беті арқылы максимальді жылдамдыққа қалай жете алатынын түсіну үшін, Фик заңына жүгіну керек. Бұл заңға сәйкес, тыныс алу беті арқылы газдардың диффузиясының жылдамдығы келесі өрнекке пропорционал.(5,9,8 бөлімдерін қарау керек, ол жерде мембрана арқылы диффузия туралы айтылады);
Ағза өзіне қажетті оттекті атмсферадан немесе өзі ерітілген судан алынады. Судағы оттектің құрылымы ауадағыдан әлдеқайда өзгеше. Ауаның көлем бірлігінде оттек біршама артық (21 ), сондай көлемдегі суға қарағанда (0, 8 ). Осыған сәйкес су ағзалары өзіннің метоболистикалық қажеттіліктері үшін дем алу бетінен су көлемін өткізу қажет, мысалы, балықтар, жердегі оыртқалыларға айтарлықтай көп көлемде ауа қажет. Бұл су жануарларының басқа вентеляция механизмінің бар болуын қарастырады. Одан басқа, суға қарағанда тығыздығы 700 есе артықтау, ал қаттылығы 100 есе көп. Ал ол дегеніміз демалу бетінен су өткізу үшін көп энергия қажет етеді. Соңында, оттек ауаға қарағанда, су арқылы 1000 есе жай диффуздайды және ол дегеніміз су ішінде демалу бетінің екі жағының арасындағы күшті концентруияланған градиентті қолдау қиынырақ. Сондықтан өкпесімен демалатын жануарларға қарағанда балықтардың метоблисттік белсенділігі әлдеқайда кем болуы таңқаларлықтай емес.
9.4.1. Біржасушалы ағзалар, амеба
Амеба – қарапайымдылардың патшалығына жататын біржасушалы ағза (Protozoa). Оның денесінің пішіні тұрақты емес, қозғалған сайын өзгеріп тұрады. Әдетте, оны диаметрі 1 мм – ден кем, ол біржасушалы ағзларға тән,сондықтан дене бетінің көлеммен қарым - қатынасы өте үлкен. Амебаның дем алу бетінің орнына сыртқы жасушының мембранасы қызмет етеді. Бұл жануардың барлық метаболисттік қажеттіліктерін қамтамасыз ету үшін,газ диффузиясы бүкіл денесінің сыртынан өте тез жылдамдықпен жүзеге асады. Оттек жасушаға енеді, ал СО2 өзінің диффузиондық градиентімен жасушадан шығады (мембрана арқылы жүретін диффузияны суреттейтін 5.9.8 бөліміне қараңыздар).
9.4.2. Мамандандырылған дем алу құрылымы мен пигменттерінің қажет етілуі
Дене көлемінің өсуіне байланысты дене беті мен көлемнің қатынасы кемиді. Енді диффузия оттегіні жасушаларға жеткілікті өткізе алмайды. Себебі олар сыртқы ортамен тікелей қатынаста емес.
9.4.1. Біржасушалы ағзалар, амеба
Амеба – қарапайымдылардың патшалығына жататын біржасушалы ағза (Protozoa). Оның денесінің пішіні тұрақты емес, қозғалған сайын өзгеріп тұрады. Әдетте, оны диаметрі 1 мм – ден кем, ол біржасушалы ағзларға тән,сондықтан дене бетінің көлеммен қарым - қатынасы өте үлкен. Амебаның дем алу бетінің орнына сыртқы жасушының мембранасы қызмет етеді. Бұл жануардың барлық метаболисттік қажеттіліктерін қамтамасыз ету үшін,газ диффузиясы бүкіл денесінің сыртынан өте тез жылдамдықпен жүзеге асады. Оттек жасушаға енеді, ал СО2 өзінің диффузиондық градиентімен жасушадан шығады (мембрана арқылы жүретін диффузияны суреттейтін 5.9.8 бөліміне қараңыздар).
9.4.2. Мамандандырылған дем алу құрылымы мен пигменттерінің қажет етілуі
Дене көлемінің өсуіне байланысты дене беті мен көлемнің қатынасы кемиді. Енді диффузия оттегіні жасушаларға жеткілікті өткізе алмайды. Себебі олар сыртқы ортамен тікелей қатынаста емес.
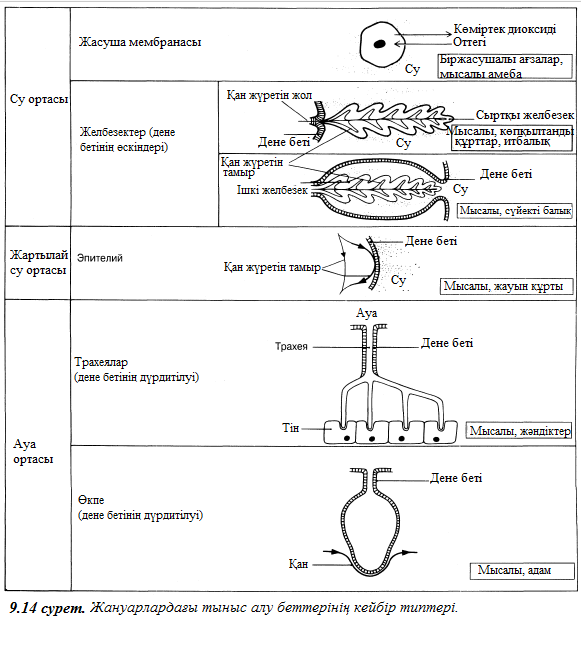
Сонымен қатар, көбіне жоғары метаболистикалық денелер отттегіні үлкен көлемде қажет етеді. Оттегіні көп қажет етудің әсерінен, кейбір дене мүшелері эволюцияға ұшырап дем алу бетіне жақын орналасқан. Әр түрлі жануарларда демалу беті әр-түрлі орналасқан. Әрбір жағдайда белгілі бір табиғатына қарай қалыптасады. 9.14 суретте көрсетілгендей әртүрлі классқа бөлуге болады. Негізінде демалу беті көлемі қандайда бір транспорттық қан жүретін жүйесі бойынша өте кеңейтілген. Транспорттық жүйе дененің барлық өзге тіндері мен дем алу бетінің байланыстарын қамтамасыз етеді және тыным алу бетімен жасушалар арасындағыоттек пен көмірқышқыл үздік алмасуын мүмкін қылып береді. Қанда демалу пегментінің болуы қанның оттегімен қамтамасыз етіп тұруына себебі болады. Одан басқа үлкен желдеткіш қозғалыстарағзадағы газалмасу қызметін жылдамдатып диффузиялық градиенттерді құрайды.
Демалу пигменттері
Демалу пигменттері дегеніміз – бұл түске боялған заттар, бұлар оттегімен тығыз араласып оны жеткізеді. Барлық белгілі демалу пигменттері боялған ақуыз емес бөліктен тұрады. Мысалы, гемоглабинде бұл гем – ақуыз молекуласымен қосындысы. Гемоглабин – қызыл пигмент жоғары кереғарлықтағы оттегіде пигмент жеңіл қосылады, ал төменде - тез береді. Қандай да бір демалу пигменті бар қан жақсы өткізгіш болады. Ал демалу пигменті жоқ қан олай өткізе алмайды. Пигмент үлкен көлемдегі оттегіні өткізеді. Сүтқоректілерде және омыртқалыларда эротроциттің құрамындағы гемоглабин демалу рөлін ойнайды. 14 бөлімінде оттегінің гемоглабинмен өткізгіштігін қарауға болады.
9.4.3. Буынқұрттар, мысалы жауын құрты
Жауын құрты. Бунақденелілер тобына жатады. Оның газөткізгішке арналған арнайы мүшелері жоқ , сондықтан газ алмасу диффузия арқылы дененің беткі қабатымен жүреді. Оларға арнайы өткізгіш мүшелері қажет емес, себебі цилиндрлік формадағы денесі арқылы төменгі белсенділікте қозғалатындықтан, оларға көп оттегі қажет емес. Бірақ буынды құрттарда қан айналу жүйесі бар, ал қан құрамында демалу пигменті -гемоглабин бар. Ірі қан тамырлары газбен бірге бүкіл денесіне жүгіреді. Сол арқылы диффузиялық градиенттер тұрақты қалыптасады. Жауын құртының жұқа терісі эпителийден бөлінетін сілекеймен ылғалданып тұрады. Эпителийлерда кутиул астында капилярлар орналасқан. Дене мен қан тамырлары арасындағы қашықтық үлкен емес, соның арқасында қанда оттегінің диффузиялануы тез жүреді. Жауын құрттардың кеуіп қалудан қорғанысы жоқ, сондықтан ылғи ылғал жерде болады.
9.4.4. Жәндіктер, мысалы шегіртке
Жәндіктерде газ айналу жұмысы трахейдегі түтікшелер жүйесі арқылы жүреді. Мұндай жүйе оттегіні тіке ауадан жасушаларға жеткізу болып саналады, сондықтан ауаны қанмен тасымалдау қажет етілмейді, бұл өте тез тәсіл және жоғарғы метобализм белсенділігін қажет етеді.
Жәндік денесіндегі түтікшелер бірінші және екінші тұстарына , асқазанға және ауа бөлігіне әкеледі. Бұл бөліктерден түтікшелер бөлініп шығады, бұларды трахей дейміз. Әрбір трахей жұқа хитин материалынан құралған. Әдетте бұл қатты қабық шиыршықты сақиналармен қалыңдатылған, соның арқасында ауа өткізгіш жолдары ашық болады. Денедегі әрбір трахей көптеген уақ түтікшелерге бөлінген, оларды трахеола дейді, трахеолаларда бөлінеді. Трахеолалардың бөліну дәрежесі метаболикалық дене қажетіне байланысты.
Демалу пигменттері
Демалу пигменттері дегеніміз – бұл түске боялған заттар, бұлар оттегімен тығыз араласып оны жеткізеді. Барлық белгілі демалу пигменттері боялған ақуыз емес бөліктен тұрады. Мысалы, гемоглабинде бұл гем – ақуыз молекуласымен қосындысы. Гемоглабин – қызыл пигмент жоғары кереғарлықтағы оттегіде пигмент жеңіл қосылады, ал төменде - тез береді. Қандай да бір демалу пигменті бар қан жақсы өткізгіш болады. Ал демалу пигменті жоқ қан олай өткізе алмайды. Пигмент үлкен көлемдегі оттегіні өткізеді. Сүтқоректілерде және омыртқалыларда эротроциттің құрамындағы гемоглабин демалу рөлін ойнайды. 14 бөлімінде оттегінің гемоглабинмен өткізгіштігін қарауға болады.
9.4.3. Буынқұрттар, мысалы жауын құрты
Жауын құрты. Бунақденелілер тобына жатады. Оның газөткізгішке арналған арнайы мүшелері жоқ , сондықтан газ алмасу диффузия арқылы дененің беткі қабатымен жүреді. Оларға арнайы өткізгіш мүшелері қажет емес, себебі цилиндрлік формадағы денесі арқылы төменгі белсенділікте қозғалатындықтан, оларға көп оттегі қажет емес. Бірақ буынды құрттарда қан айналу жүйесі бар, ал қан құрамында демалу пигменті -гемоглабин бар. Ірі қан тамырлары газбен бірге бүкіл денесіне жүгіреді. Сол арқылы диффузиялық градиенттер тұрақты қалыптасады. Жауын құртының жұқа терісі эпителийден бөлінетін сілекеймен ылғалданып тұрады. Эпителийлерда кутиул астында капилярлар орналасқан. Дене мен қан тамырлары арасындағы қашықтық үлкен емес, соның арқасында қанда оттегінің диффузиялануы тез жүреді. Жауын құрттардың кеуіп қалудан қорғанысы жоқ, сондықтан ылғи ылғал жерде болады.
9.4.4. Жәндіктер, мысалы шегіртке
Жәндіктерде газ айналу жұмысы трахейдегі түтікшелер жүйесі арқылы жүреді. Мұндай жүйе оттегіні тіке ауадан жасушаларға жеткізу болып саналады, сондықтан ауаны қанмен тасымалдау қажет етілмейді, бұл өте тез тәсіл және жоғарғы метобализм белсенділігін қажет етеді.
Жәндік денесіндегі түтікшелер бірінші және екінші тұстарына , асқазанға және ауа бөлігіне әкеледі. Бұл бөліктерден түтікшелер бөлініп шығады, бұларды трахей дейміз. Әрбір трахей жұқа хитин материалынан құралған. Әдетте бұл қатты қабық шиыршықты сақиналармен қалыңдатылған, соның арқасында ауа өткізгіш жолдары ашық болады. Денедегі әрбір трахей көптеген уақ түтікшелерге бөлінген, оларды трахеола дейді, трахеолаларда бөлінеді. Трахеолалардың бөліну дәрежесі метаболикалық дене қажетіне байланысты.
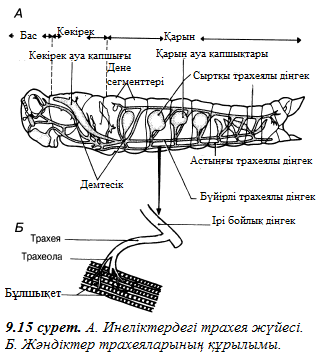
|
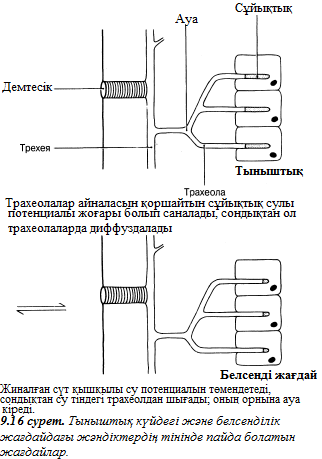
|
Трахеолаларда хитин материалдары жоқ. Қозғалыстық жағдайда олар суға толтырылып тұрады. Бұл кезде оттегі жасушаларға жәндіктің қауіпсіз жағдайын толықтырып тұрады. Белсенді кезде бұлшықтардың жылдам қозғалысы метаболиттрдің құралуына әкеледі, көбіне сүт қышқылына, ал жасушаларда осмотикалық қысым өседі. Мұндай жағдайда трахеолдан осматикалық күш арқылы сұйықтық жасушаларға сіңеді және трахеолдарға ауа көптеп келеді. Сол себепті оттегі тіке жасушаларға керек кезінде жетеді.
Жалпы ауа көлемі жәндік денесіне демалыс түтікшелері арқылы кіріп реттеліп тұрады. Әрбір түтікше басы кішкентай бұлшықтардан құралған қалпақшаболады. Әрбір қалпақшаның шетінде түтікшелері бар: олар түтікшелерге ауадан басқа заттардың кіруіне кедергі болады.
Жәндіктің жылдам демалуы СО2– нің де жылдам қозғалуына әкеледі. Хеморецепторлар бұны бақылап түтікшелер ашылады. Осы қимылдар вентеляциондық қызметінде атқарады, ірі жәндіктерде болады,әсіресе шегірткеде. Дорсовенталдық бұлшықтар тартылып, жәндік денесін ықшам қылады.
Сол себепті трахей жүйе көлемі кішіриеді, ал ауа сыртқа шығады. Ауаны сору жәй жүреді, бұл кезде дене сигменттері өзінің серпімділігіне қалыпты пішінге түседі. Кеуде түтікшелері кезекпен ашылып –жабылып, соның арқасында денеге вентеляционды ауа түтікше арқылы кіріп кеуде арқылы шығады. Т
рахейлық жүйе газ алмасу жағдайында тиімді, сондықтан көптеген жәндіктерде газ алмасу тек диффузияның дене арқылы дем алуында болады,бундай кішкентай арақашықтықта диффузияға тиімді 2 см дан аспайды, сондықтан ұзындығы 30 см жәндіктер кездессе де олардың дене қалыңдығы 2см дан аспауы керек.
9.4.5. Сүйекті балықтар, мысалы майшабақ
Сүйекті балықтарда жұтқыншақ қабырғасында Желбезек шель болады, солар арқылы балықтар сыртқы ортамен араласады, мысалы сумен. Желбезек шелдері арасында сүйектерден орналасқан жабыр айналымдары бар. Сүйекті балықтарда төрт жабыр жұбы болады, және бөлініп тұрған бес жабыр жұбы болады. Әрбір жабыр екі қатар желбезек жапырақтардан тұрады. Олар V әрпі түрінде орналасқан. Жапырақтары жаелбезек пластинкаларды құрайды. Олар көптеген капиллярдан тұрады. Желбезек пластинкалар ауа өткізу ауқымдылығын күшейтеді. Қанды судан айырып тұрған барьер қалыңдығы бірнеше клеткадан аспайды. Сондықтан бұл барьер арқылы газдың диффузиялануы өте жылдам жүреді. Қозғалыстағы желбезек қақпақша жабырларды жауып және қорғап тұрады. Желбезектерді ішкі қабаттан айырып тұратын жерін желбезек қуыс дейді. Желбезек қақпақша вентеляциондық қызмет атқарады. Ол ашылып және мықты жабылып ауа
Жалпы ауа көлемі жәндік денесіне демалыс түтікшелері арқылы кіріп реттеліп тұрады. Әрбір түтікше басы кішкентай бұлшықтардан құралған қалпақшаболады. Әрбір қалпақшаның шетінде түтікшелері бар: олар түтікшелерге ауадан басқа заттардың кіруіне кедергі болады.
Жәндіктің жылдам демалуы СО2– нің де жылдам қозғалуына әкеледі. Хеморецепторлар бұны бақылап түтікшелер ашылады. Осы қимылдар вентеляциондық қызметінде атқарады, ірі жәндіктерде болады,әсіресе шегірткеде. Дорсовенталдық бұлшықтар тартылып, жәндік денесін ықшам қылады.
Сол себепті трахей жүйе көлемі кішіриеді, ал ауа сыртқа шығады. Ауаны сору жәй жүреді, бұл кезде дене сигменттері өзінің серпімділігіне қалыпты пішінге түседі. Кеуде түтікшелері кезекпен ашылып –жабылып, соның арқасында денеге вентеляционды ауа түтікше арқылы кіріп кеуде арқылы шығады. Т
рахейлық жүйе газ алмасу жағдайында тиімді, сондықтан көптеген жәндіктерде газ алмасу тек диффузияның дене арқылы дем алуында болады,бундай кішкентай арақашықтықта диффузияға тиімді 2 см дан аспайды, сондықтан ұзындығы 30 см жәндіктер кездессе де олардың дене қалыңдығы 2см дан аспауы керек.
9.4.5. Сүйекті балықтар, мысалы майшабақ
Сүйекті балықтарда жұтқыншақ қабырғасында Желбезек шель болады, солар арқылы балықтар сыртқы ортамен араласады, мысалы сумен. Желбезек шелдері арасында сүйектерден орналасқан жабыр айналымдары бар. Сүйекті балықтарда төрт жабыр жұбы болады, және бөлініп тұрған бес жабыр жұбы болады. Әрбір жабыр екі қатар желбезек жапырақтардан тұрады. Олар V әрпі түрінде орналасқан. Жапырақтары жаелбезек пластинкаларды құрайды. Олар көптеген капиллярдан тұрады. Желбезек пластинкалар ауа өткізу ауқымдылығын күшейтеді. Қанды судан айырып тұрған барьер қалыңдығы бірнеше клеткадан аспайды. Сондықтан бұл барьер арқылы газдың диффузиялануы өте жылдам жүреді. Қозғалыстағы желбезек қақпақша жабырларды жауып және қорғап тұрады. Желбезектерді ішкі қабаттан айырып тұратын жерін желбезек қуыс дейді. Желбезек қақпақша вентеляциондық қызмет атқарады. Ол ашылып және мықты жабылып ауа
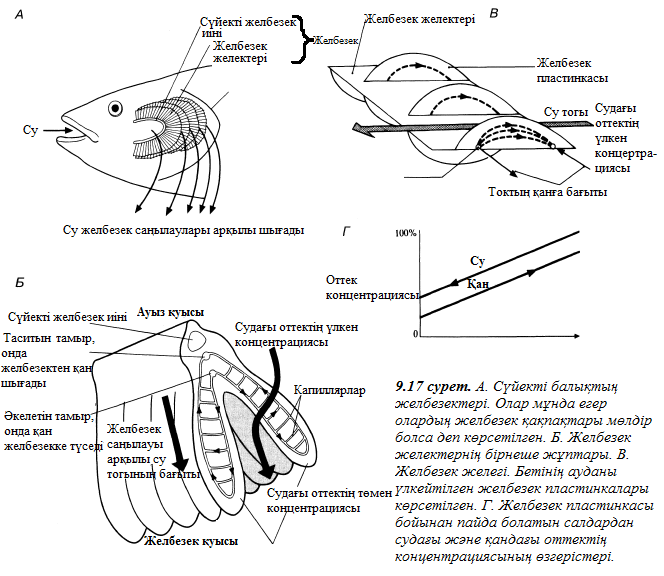
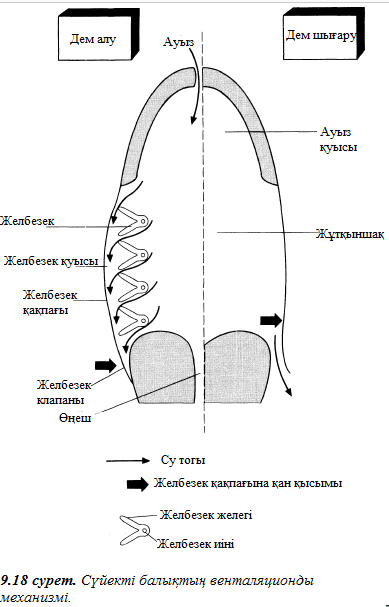
|
тоғының ішке кіріп шығуын қадағалап тұрады. Ау жұтқан кезде ауыз қуысы үлкиеді, ішке кірген су жабыр қақпақшалары арқылы жабылып қалады. Жабыр қуысындағы қысым түсіп ауыз қуысы арқылы жабыр қуысынан газ алмасу қызметі жүреді. Ауа шығарған кезінде ауыз қуысы мен асқазан қуысы жабылады, АУЫЗ ТҮБІ ҚУЫСЫ КӨТЕРІЛЕДІ, сол себепті су жабыр қуыстарына қарай итеріледі . сөйтіп, соңғы жабыр қақпақшасы арқылы сыртқа шығады. Жабыр пластинкаларындағы қан суға қарама қарсы ағады. Бұндай қарама қарсы жүйе өте тиімді. Қарама қарсы жүйеде жүретін қан өз жолында үнемі сумен кездесіп отырады, ал онаң құрамында ерітілген оттегінің қатынасы өте жоғары.
|
9.5. Сүтқоректілердің газ алмасуы
9.5.1. Дем алу жүйесінің құрылымы
Сүтқоректілердің демалу бетін альвеола деп аталатын ауамен толтырылған коптеген көпіршіктер құрайды. Альвеолалар атмосфералық ауамен байланысты бир қатар пневматикалық түтіктермен және журектің қасындағы кеуде қуысында орналасқан (9.19 сурет) Осы түтіктермен ауа өкпеге өтеді. 12 жұб сүйек қабырғалары жүрек пен өкпені қоршап, қорғап тұрады. Қабырғаларға қабырға бұлшығы тіркелген , ал кеуде қуысын іш қуысынан кең диафрагма айырады. Бұл бұлшық және диафрагма біз 9.5.4. бөлімінде қарастыратын вентиляционный миханизм жумысында да қатысады.
Ауа ағзаға бөтен нәрселерден сақтайтын шаш талшықтары бар екі мұрын танауы арқылы жетеді. Мұрын жолымен өткен ауа жылынады және ылғалданады. Біруақытта жануарлар оларға келген иісті сезеді. Әрі қарай ауа көмей арқылы трахеяға түсіп кету керек. трахея өңеш алдында тіке жатады және көкірек қуысында бітеді. Трахея қабырғалары С – тәрізді шеміршектермен бекінген; ол осы шеміршектердің арқасында ашық болып қалады. Өзінің жабылмаған жағында С – тәрізді шеміршектер өңешке байланысқан (9.19 сурет); олар трахеяларды тыныс алуда құлап қалуына жол бермейді. Мұндай шеміршекті трахея арқылы көлденең кесіндіде көруге болады (9.20, А сурет).
Трахея төменгі соғында 2 бронхаға бөлінеді. Өкпеде әрбір бронх бронхиолалар деп аталатын көпеселі кішілеу түтікшелерге бөлінеді. Бұл соңғысы өз кезегінде өте кішілеу түтікшелерді түзеді, ол альвеолды қаптарды алып жүретін альвеолды жүрістен деп аталады. Әрбір осындай қапта оған альвеол ашып беретін топ болады. 9.21 суретінде бұл тыныс алу жолының және оның бөлек бөлімшелерінің гистологиялық сипаттамаларының кескіні берілген. Ауалы жолдың басым көпшілігінің қабырғалары жыбырлағыш эпителийлермен тқселген. Эпителий қалығдығы шырын құпиясын бөліп шығаратын құтыша тәрізді жасушаларындікіндей (9.20 және 6.16 сурет). Шырыш танау айналасындағы кез келген бөлшектерді түк жиегі арқылы біледі, мысалы тозаң бөлшектері мен бактериялар. Кірпік соққылары бұл сезілген шырыш бөлшектерін ауыз қуысының артқы қабырғасына бағыттайды және бұл жерде олар жұтылып қойылады (сондықтан да олар ауалы жолға тіптен түсе алмайды). Шет бөлшектер кірпікшелермен сезілмейтінін белгілейміз, олар шырышпен сезіледі, онда танау айналасындағы талшық шеті сияқты, оларды тек қана талшықтар ұстап тұрады. Шырыш басқа да функцияны орындайды: ол дем алған ауаны ылғалдатады.
9.5.1. Дем алу жүйесінің құрылымы
Сүтқоректілердің демалу бетін альвеола деп аталатын ауамен толтырылған коптеген көпіршіктер құрайды. Альвеолалар атмосфералық ауамен байланысты бир қатар пневматикалық түтіктермен және журектің қасындағы кеуде қуысында орналасқан (9.19 сурет) Осы түтіктермен ауа өкпеге өтеді. 12 жұб сүйек қабырғалары жүрек пен өкпені қоршап, қорғап тұрады. Қабырғаларға қабырға бұлшығы тіркелген , ал кеуде қуысын іш қуысынан кең диафрагма айырады. Бұл бұлшық және диафрагма біз 9.5.4. бөлімінде қарастыратын вентиляционный миханизм жумысында да қатысады.
Ауа ағзаға бөтен нәрселерден сақтайтын шаш талшықтары бар екі мұрын танауы арқылы жетеді. Мұрын жолымен өткен ауа жылынады және ылғалданады. Біруақытта жануарлар оларға келген иісті сезеді. Әрі қарай ауа көмей арқылы трахеяға түсіп кету керек. трахея өңеш алдында тіке жатады және көкірек қуысында бітеді. Трахея қабырғалары С – тәрізді шеміршектермен бекінген; ол осы шеміршектердің арқасында ашық болып қалады. Өзінің жабылмаған жағында С – тәрізді шеміршектер өңешке байланысқан (9.19 сурет); олар трахеяларды тыныс алуда құлап қалуына жол бермейді. Мұндай шеміршекті трахея арқылы көлденең кесіндіде көруге болады (9.20, А сурет).
Трахея төменгі соғында 2 бронхаға бөлінеді. Өкпеде әрбір бронх бронхиолалар деп аталатын көпеселі кішілеу түтікшелерге бөлінеді. Бұл соңғысы өз кезегінде өте кішілеу түтікшелерді түзеді, ол альвеолды қаптарды алып жүретін альвеолды жүрістен деп аталады. Әрбір осындай қапта оған альвеол ашып беретін топ болады. 9.21 суретінде бұл тыныс алу жолының және оның бөлек бөлімшелерінің гистологиялық сипаттамаларының кескіні берілген. Ауалы жолдың басым көпшілігінің қабырғалары жыбырлағыш эпителийлермен тқселген. Эпителий қалығдығы шырын құпиясын бөліп шығаратын құтыша тәрізді жасушаларындікіндей (9.20 және 6.16 сурет). Шырыш танау айналасындағы кез келген бөлшектерді түк жиегі арқылы біледі, мысалы тозаң бөлшектері мен бактериялар. Кірпік соққылары бұл сезілген шырыш бөлшектерін ауыз қуысының артқы қабырғасына бағыттайды және бұл жерде олар жұтылып қойылады (сондықтан да олар ауалы жолға тіптен түсе алмайды). Шет бөлшектер кірпікшелермен сезілмейтінін белгілейміз, олар шырышпен сезіледі, онда танау айналасындағы талшық шеті сияқты, оларды тек қана талшықтар ұстап тұрады. Шырыш басқа да функцияны орындайды: ол дем алған ауаны ылғалдатады.
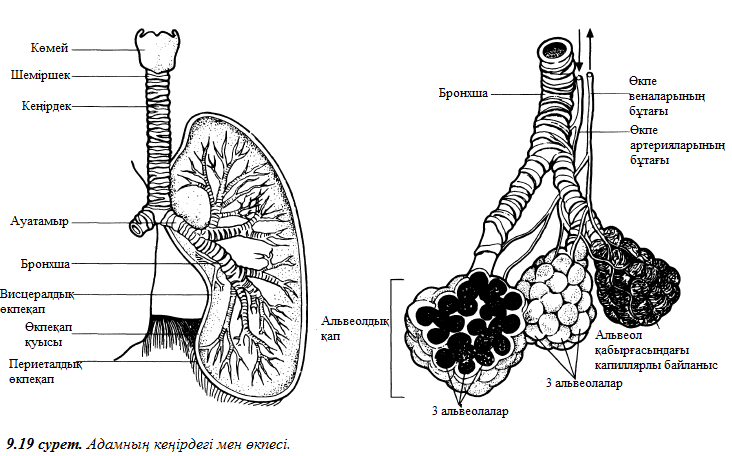
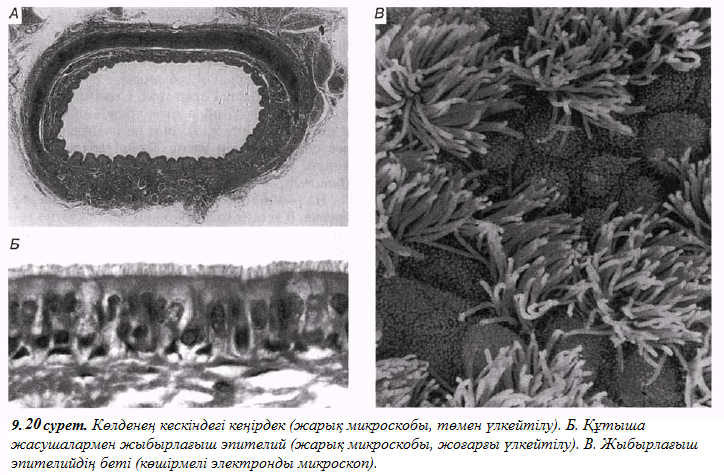
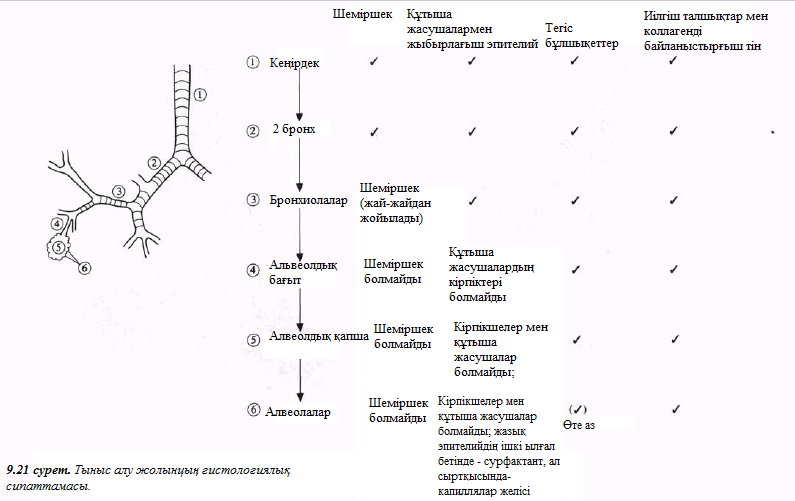
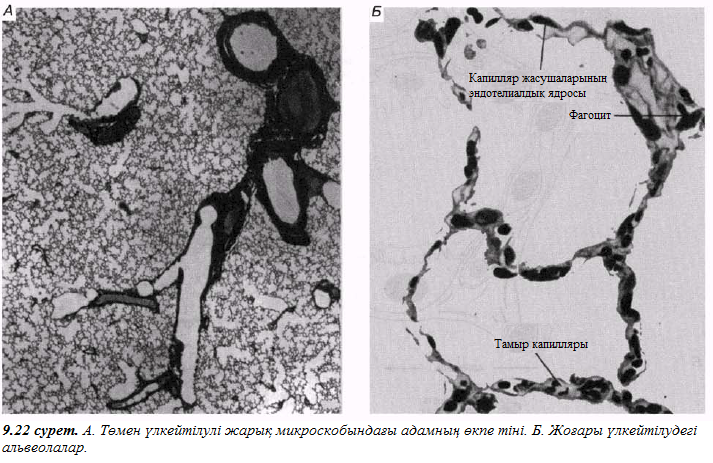
Альвеол құрылымы
Альвеол қабырғасы – газалмасу жүретін бет (9.22 сурет). Адам өкпесінде жалпылама аудан бетінде, яғни 70 – 90 кв. м – де 700 млн – ге дейін альвеол болады. Альвеол қабырғасының қалыңдығы бар болғаны 0,0001 мм – ді (0,1 мкм) құрайды. Альвеол қабырғасының мыртқы жағы қан жүретін тығыз желімен жабылған; оның барлығы бастауды өкпе артериясынан алады және ең соңында өкпе венасын түсе отырып байланысады (9.23 және 9.24 суреттер). Әрбір алвеола ылғал тегіс эпителиймен төселген. Оның жасушалары газ диффуздайтын кішілеу тосқауыл жасайтын болып тегістелген (9.24 сурет). Альвеолды қабырғаларда коллаген мен оған иілгіштік пен альвеолдарына өздерінің көлемін тыныс алу мен тыныс шығаруда өзгертуге мүмкіндік беретін иілгіш талшық болмайды.
Альвеол қабырғасы – газалмасу жүретін бет (9.22 сурет). Адам өкпесінде жалпылама аудан бетінде, яғни 70 – 90 кв. м – де 700 млн – ге дейін альвеол болады. Альвеол қабырғасының қалыңдығы бар болғаны 0,0001 мм – ді (0,1 мкм) құрайды. Альвеол қабырғасының мыртқы жағы қан жүретін тығыз желімен жабылған; оның барлығы бастауды өкпе артериясынан алады және ең соңында өкпе венасын түсе отырып байланысады (9.23 және 9.24 суреттер). Әрбір алвеола ылғал тегіс эпителиймен төселген. Оның жасушалары газ диффуздайтын кішілеу тосқауыл жасайтын болып тегістелген (9.24 сурет). Альвеолды қабырғаларда коллаген мен оған иілгіштік пен альвеолдарына өздерінің көлемін тыныс алу мен тыныс шығаруда өзгертуге мүмкіндік беретін иілгіш талшық болмайды.
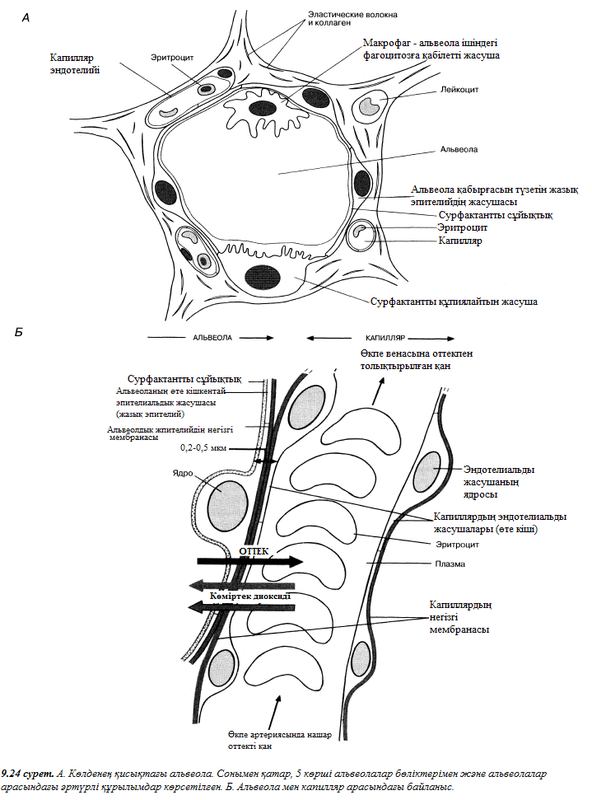
Альвеол қабырғасының ерекше жасушалары оның ішкі бетіне жуғыш қасиеттері бар зат сурфактнатты бөліп шығарады. Бұл зат эпителий альвеоларына төселген дымқыл қабаттың беткі керілуін төмендетеді, тыныс алуда өкпенің кеңеюінің арқасында күш аз кетеді. Сурфактант, сонымен қатар, бұл дымқыл қабат арқылы өтетін оттек пен СО2 – лердің тасылымын тездетеді. Одан басқа, альвеоларда пайда болған бактерияларды да жоя алады. Сау өкпелерде сурфактант үздіксіз құпияланады және қайта сіңіріледі. Адам ұрығында, шамамен, алғаш рет 23 – ші аптада пайда болады. Бұл маңызды себептердің бірі болып табылады. егер ұрық 24 аптаға дейін баратын болса, онда ол өздігінен өмір сұруге қабілетсіз. Бұған да нақты мерзім белгіленеді, күні бұрынды ынталандыру берілетін тұқымға Ұлыбританияда тыйым салған екен. Күніне жетпей туылған бөбектерде сурфактант болмайды деп ұйғарылған. Бұның артынан шала туылған бөбектердің өлімінің ең маңызды себептерінің бірі болып саналатын – тыныс алу бұзылуының синдромы пайда болады. Сурфактантсыз альвеоланың керілу сұйық бетінде норманы 10 есе арттырады және альвеолалар әрбір дем шығарудан кейін құлайды. Ал олардың дем алуда қайтып кеңеюлері үшін айтарлықтай көп күш талап етіледі.
9.5.2. Альвеолалардағы газалмасу
Оттегі альвеолара альвеолды қабырғасының эпителийі мен капиллярлар эндотелиясынан тұратын кіші тосқауыл арқылы диффуздалады (9.24, Б сурет). Ол, алдымен, қан плазмасына түседі және нәтижесінде оксигемоглобинге айналатын эритроцит гемоглобинімен байланысады. Көмірқышқыл газы (көміртек диоксиді) – кері бағытта диффуздайды, яғни, қаннан альвеол қуысына.
Тиімді диффузиялар қабілетті:
9.5.2. Альвеолалардағы газалмасу
Оттегі альвеолара альвеолды қабырғасының эпителийі мен капиллярлар эндотелиясынан тұратын кіші тосқауыл арқылы диффуздалады (9.24, Б сурет). Ол, алдымен, қан плазмасына түседі және нәтижесінде оксигемоглобинге айналатын эритроцит гемоглобинімен байланысады. Көмірқышқыл газы (көміртек диоксиді) – кері бағытта диффуздайды, яғни, қаннан альвеол қуысына.
Тиімді диффузиялар қабілетті:
- Альвел беті ауданының үлкендігі;
- Диффуздалған газды бөліп алуға талап етілетін қысқа арақашықтық;
- Қанның тұрақты түсімі және оттегіні тасымалдайтын – гемоглобині болатын вентеляциямен жабдықталған мықты диффузионды градиенті бар;
- Сурфактанттың болуы.
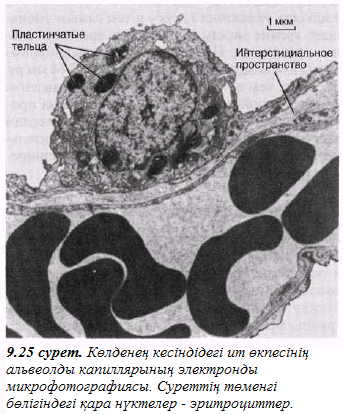
Капилляр альвеоларының диаметрі эритроциттің диаметриінен кішілеу және эритроциттер олар арқылы қан қысымын қысып тастайды. Мұндайда олар формасын өзгертеді (9.25 сурет) және олардың бетінің көп үлесі альвеол бетімен байланысуға әкеледі, осының арқасында олар оттекті көптеп сіңіре алады. Одан басқа, эритроциттер каппилярмен жай қозғалады, сондықтан алмасу ұзағырақ болуы мүмкін. Қан альвеолаларды тастаған кезде, оттек пен СО2 – ның парциалдық қысымы оның ішінде де альвеолдық ауада болады.
9.5.3. Өкпеқап қуысы
Көкірек жасушасында болатын өкпе олардың қабырғаларынан иілгіш әйнектей қабатпен (өкпеқаппен) жабылған тесік тәрізді көлік – өкпеқап қуысымен бөлінген. Ол өкпені қорғайды, ауаны олардан көкірек қуысына өтіп кетпеуіне жол бермейді және өкпе мен көкрек жасушаларының қабырғаларының арасындағы үйкелісті азайтады. Ішкі, ішкіағзалық өкпеқап жапырағы өкпені жабады, ал сыртқы қабырғалығы көкірек жасушалары мен көкетті төсейді. Өкпеқап қуысында өкпеқаптан бөлініп шығатын сұйықтық болады. Өкпеқап қуысы ауа үшін өткізбейтін болып табылады және өкпеге қарағанда онда қысым 3 – 4 мм рт. ст төмендеу. Өкпеқап қуысында теріс қысым барлық дем алу бойында ұсталады, альвеолаларға тегістеліне алады және көкірек жасушаларының үлкеюінде пайда болатын кез келген қосымша кеңістікті толтырады.
9.5.4. Вентеляция механизмі (дем алу)
Ауа өкпе кіреді және қабырғааралық бұлшықеттер мен диафрагма қызметтерінің арқасында ол жақтан шығады; нәтижесінде, олардың үзіліс сайынғы қысқартылуы және көкірек жасушаларының көлемінің әлсіздігі өзгереді. Әрбір қабырға жұптарының арасында қабырғааралық бұлшықеттердің 2 тобы орналасқа, олар бір – біріне бұрыштап бағытталғын, яғни, сыртқылары – төменде және алда, ал ішкілері – төменде және артта (9.26 сурет). Көкет айналасында орталықты сіңірлі колагеннен тұратын бөлім орналасқан сақиналы және радиалды бұлшықет талшықтарынан тұрады.
9.5.3. Өкпеқап қуысы
Көкірек жасушасында болатын өкпе олардың қабырғаларынан иілгіш әйнектей қабатпен (өкпеқаппен) жабылған тесік тәрізді көлік – өкпеқап қуысымен бөлінген. Ол өкпені қорғайды, ауаны олардан көкірек қуысына өтіп кетпеуіне жол бермейді және өкпе мен көкрек жасушаларының қабырғаларының арасындағы үйкелісті азайтады. Ішкі, ішкіағзалық өкпеқап жапырағы өкпені жабады, ал сыртқы қабырғалығы көкірек жасушалары мен көкетті төсейді. Өкпеқап қуысында өкпеқаптан бөлініп шығатын сұйықтық болады. Өкпеқап қуысы ауа үшін өткізбейтін болып табылады және өкпеге қарағанда онда қысым 3 – 4 мм рт. ст төмендеу. Өкпеқап қуысында теріс қысым барлық дем алу бойында ұсталады, альвеолаларға тегістеліне алады және көкірек жасушаларының үлкеюінде пайда болатын кез келген қосымша кеңістікті толтырады.
9.5.4. Вентеляция механизмі (дем алу)
Ауа өкпе кіреді және қабырғааралық бұлшықеттер мен диафрагма қызметтерінің арқасында ол жақтан шығады; нәтижесінде, олардың үзіліс сайынғы қысқартылуы және көкірек жасушаларының көлемінің әлсіздігі өзгереді. Әрбір қабырға жұптарының арасында қабырғааралық бұлшықеттердің 2 тобы орналасқа, олар бір – біріне бұрыштап бағытталғын, яғни, сыртқылары – төменде және алда, ал ішкілері – төменде және артта (9.26 сурет). Көкет айналасында орталықты сіңірлі колагеннен тұратын бөлім орналасқан сақиналы және радиалды бұлшықет талшықтарынан тұрады.
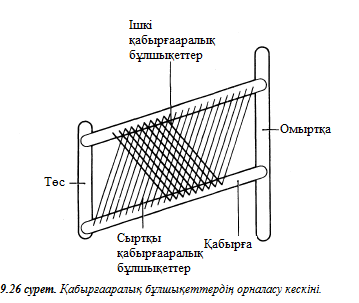
Дем алу
Дем алу – бұл белсенді үрдіс. Ол келесі кейіппен жүреді.
Дем алу – бұл белсенді үрдіс. Ол келесі кейіппен жүреді.
- Сыртқы қабырғааралық бұлшықеттер қысқарады, ал ішкілері әлсірейді.
- Қабырға осының салдарынан омыртқадан жойылып, алға бұрылып кетеді. (Бұны сезу оңай, қолды дем алужа көкірекке қойып қарау керек).
- Бір уақытта, көкет бұлшықеттері қысқарады.
- Диафрагма тегістеу болады.
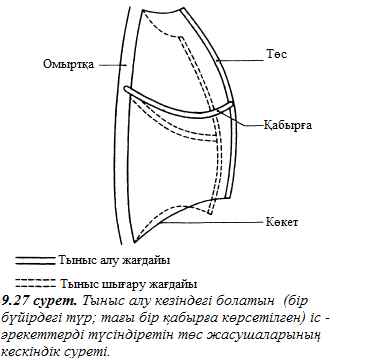
5. Көкірек жасушалар қысымының нәтижесінде және демек, өкпеде атомосфералықтан төмен болып қалады.
6. Ауа ішке кіреді және альвеолаларды өкпедегі қысым атмосфераныкіндей болмайынша, толтыра береді.Дем шығару
Дем шығару – созылыңқы өкпе тіндерінің иілгіштік қысқаруы, дем алу бұлшықеттері бөлшектерінің әрсіретілуі мен көкірек жасушаларының ауырлық күшінің әсерінен түсіп қалуы нәтижесінде болатын әдеттегі жағдайларда негізінде енжар жағдайында болатын үрдіс.
9.5.5. Дем алу реттелулері
Әдетте, адам оның қалай тыныс алып жатқанын байқамайды, себебі бұл оның еркіне тәуелсіз реттеледі. Бірақ, белгілі бір дәрежеде, тыныс алу саналы түрде реттелуі мүмкін, біз мұны төменде қарастырамыз.
Тыныс алудың еріксіз реттелуі сопақша мидағы (артқы мидың бір бөлімі) (9.28 сурет) тыныс алу орталығында жүзеге асады. Тыныс алу орталығының құрсақтұс (төменгі) жағы тыныс алудың ынталандырылуына жауапты; оны тыныс алу орталығы (ентігу орталығы) деп атайды. Бұл орталықтың ынталандырылуы жиілік пен дем алу тереңдігін арттырады. Сыртқы (жоғарғы) және қос бүйір бөлімдері тыныс алуды тежейді; олар тыныс алу орталығы (қиындау орталығы) деп аталатын жинақтама атауды алып жүреді. Тыныс алу орталығы қабырғааралық бұлшықеттерің қабырғааралық жүйкелерімен байланысты, ал көкетпен – көкеттік байланысты. Бронхиалды ағаш (бронха мен бронхиолалардың жинағы) жылжымалы жүйкемен жүйкеленеді (9.28 сурет). Көкет пен қабырғааралық бұлшықеттерге бағытталатын бір қалыпта қайталана беретін жүйке импулсьтері вентеляциондық қозғалыстардың жүзеге асуын қамтамасыз етеді.
Дем шығаруда өкпенің кеңейтілуі бронхиалды ағашта болатын созылу рецепторларын ынталандырады және олар жылжымалы жүйке арқылы көп – көптен испульстерді қиындау орталығына жіберіп отырады. Бұл уақытында ернтігк орталығы мен дем алуды басып тастайды. Сыртқы қабырғааралық бұлшықеттер енді әлсіретіледі, созылған өкпе тіні иілгішті қысқарады – дем шығару болады. днм шығарудан кейінгі созылу рецепторлары бронхиалды ағашта енді ынталандырылуда кездеспейді. Сондықтан қиындау центрі сөнеді және тыныс алу қайтадан басталуы мүмкін. Бұл барлық айналым үздіксіз және бір қалыпта ағзаның барлық тіршілік бойына қайталана береді. Жылдамдатылған тыныс алу ішкі қабырғааралыө бұлшықеттердің қатысуымен жүзеге асады.
Тыныс алудың негізгі ырғағы сопақша мидың тыныс алу орталығымен қуатталады, тіпті егер барлық оған кіретін кесілген жүйкелер де. Бірақ әдеттегі жағдайларда бұл негізгі ырғақта әртүрлі әсерлер таңылады. Тыныс алу жиілігі реттелуінің негізгі факторы қандағы оттек концентрациясы емес, СО2 концентрациясы болып табылады. СО2 жүктемесі артқан кезде (мысалы, физикалық жүктелуде) қан жүретін тамырда болатын күретамыр хеморецепторлары мен қолқа түйіршігін (9.28 сурет) жүйке импульстері ентігу орталығына жіберіледі. Нағыз сопақша миында хеморецепторлар да болады. ентігу орталығынан көкет пен қабырғааралық жүйкелер арқылы импульстер көкет пен сыртқы қабырғааралық бұлшықеттерге түседі, оларды жиі қысқаруға апарады, Ағзада жиналған СО2 ағзаға үлкен қауіп төндіруі мүмкін. СО2 – нің сумен байланысында фермент және өзге де нәруыздардың денатурациясын (табиғатын жоғалтқан) шақыртуға бейім қышқылды түзеді. Сондықтан ағзалар эволюциясы кезеңінде СО2 – нің кез келген концентрациясының көтерілуіне өте тез реакция жаттықтырылды. Егер СО2 концентрациясы ауада 0,25 %, онда өкпе вентеляциясы екі есе арттырылады. Осындай нәтижені шақырту үшін, оттек концентрациясы ауа 20 %– дан 5% – ға дейін түсуі керек. оттек концентрациясы тыныс алуға жа әсер береді, бірақ оттектің әдеттегі жағдайларында әрқашан жеткілікті болады, сондықтан оған тиер әсер үлкен емес. оттек концентрациясына әсерін тигізетін хеморецепторлар слпақша мида, куретамырда және қолқа көпіршіктерінде, сонымен қатар СО2 рецепторларында орналасады.
6. Ауа ішке кіреді және альвеолаларды өкпедегі қысым атмосфераныкіндей болмайынша, толтыра береді.Дем шығару
Дем шығару – созылыңқы өкпе тіндерінің иілгіштік қысқаруы, дем алу бұлшықеттері бөлшектерінің әрсіретілуі мен көкірек жасушаларының ауырлық күшінің әсерінен түсіп қалуы нәтижесінде болатын әдеттегі жағдайларда негізінде енжар жағдайында болатын үрдіс.
- Сыртқы қабырғааралық бұлшықеттер әлсірейді, ал ішкілері – қысқарады. Көкірек жасушасы, ең алдымен, өзіндік ауырлық әсерінен түсіп қалады.
- Біруақытта көкет әлсіретіледі. Түсп қалған көкірек жасушасы оның негізгі күмбез тәрізді пішінін қабылдауға мәжбүр болады.
- Бұның салдарынан, көкірек жасушасының көлемі кішінрейеді және ондағы қысым атомосфералықтан жоғарылау болады.
- Нәтижесінде, ауа өкпеден шығарылады.
9.5.5. Дем алу реттелулері
Әдетте, адам оның қалай тыныс алып жатқанын байқамайды, себебі бұл оның еркіне тәуелсіз реттеледі. Бірақ, белгілі бір дәрежеде, тыныс алу саналы түрде реттелуі мүмкін, біз мұны төменде қарастырамыз.
Тыныс алудың еріксіз реттелуі сопақша мидағы (артқы мидың бір бөлімі) (9.28 сурет) тыныс алу орталығында жүзеге асады. Тыныс алу орталығының құрсақтұс (төменгі) жағы тыныс алудың ынталандырылуына жауапты; оны тыныс алу орталығы (ентігу орталығы) деп атайды. Бұл орталықтың ынталандырылуы жиілік пен дем алу тереңдігін арттырады. Сыртқы (жоғарғы) және қос бүйір бөлімдері тыныс алуды тежейді; олар тыныс алу орталығы (қиындау орталығы) деп аталатын жинақтама атауды алып жүреді. Тыныс алу орталығы қабырғааралық бұлшықеттерің қабырғааралық жүйкелерімен байланысты, ал көкетпен – көкеттік байланысты. Бронхиалды ағаш (бронха мен бронхиолалардың жинағы) жылжымалы жүйкемен жүйкеленеді (9.28 сурет). Көкет пен қабырғааралық бұлшықеттерге бағытталатын бір қалыпта қайталана беретін жүйке импулсьтері вентеляциондық қозғалыстардың жүзеге асуын қамтамасыз етеді.
Дем шығаруда өкпенің кеңейтілуі бронхиалды ағашта болатын созылу рецепторларын ынталандырады және олар жылжымалы жүйке арқылы көп – көптен испульстерді қиындау орталығына жіберіп отырады. Бұл уақытында ернтігк орталығы мен дем алуды басып тастайды. Сыртқы қабырғааралық бұлшықеттер енді әлсіретіледі, созылған өкпе тіні иілгішті қысқарады – дем шығару болады. днм шығарудан кейінгі созылу рецепторлары бронхиалды ағашта енді ынталандырылуда кездеспейді. Сондықтан қиындау центрі сөнеді және тыныс алу қайтадан басталуы мүмкін. Бұл барлық айналым үздіксіз және бір қалыпта ағзаның барлық тіршілік бойына қайталана береді. Жылдамдатылған тыныс алу ішкі қабырғааралыө бұлшықеттердің қатысуымен жүзеге асады.
Тыныс алудың негізгі ырғағы сопақша мидың тыныс алу орталығымен қуатталады, тіпті егер барлық оған кіретін кесілген жүйкелер де. Бірақ әдеттегі жағдайларда бұл негізгі ырғақта әртүрлі әсерлер таңылады. Тыныс алу жиілігі реттелуінің негізгі факторы қандағы оттек концентрациясы емес, СО2 концентрациясы болып табылады. СО2 жүктемесі артқан кезде (мысалы, физикалық жүктелуде) қан жүретін тамырда болатын күретамыр хеморецепторлары мен қолқа түйіршігін (9.28 сурет) жүйке импульстері ентігу орталығына жіберіледі. Нағыз сопақша миында хеморецепторлар да болады. ентігу орталығынан көкет пен қабырғааралық жүйкелер арқылы импульстер көкет пен сыртқы қабырғааралық бұлшықеттерге түседі, оларды жиі қысқаруға апарады, Ағзада жиналған СО2 ағзаға үлкен қауіп төндіруі мүмкін. СО2 – нің сумен байланысында фермент және өзге де нәруыздардың денатурациясын (табиғатын жоғалтқан) шақыртуға бейім қышқылды түзеді. Сондықтан ағзалар эволюциясы кезеңінде СО2 – нің кез келген концентрациясының көтерілуіне өте тез реакция жаттықтырылды. Егер СО2 концентрациясы ауада 0,25 %, онда өкпе вентеляциясы екі есе арттырылады. Осындай нәтижені шақырту үшін, оттек концентрациясы ауа 20 %– дан 5% – ға дейін түсуі керек. оттек концентрациясы тыныс алуға жа әсер береді, бірақ оттектің әдеттегі жағдайларында әрқашан жеткілікті болады, сондықтан оған тиер әсер үлкен емес. оттек концентрациясына әсерін тигізетін хеморецепторлар слпақша мида, куретамырда және қолқа көпіршіктерінде, сонымен қатар СО2 рецепторларында орналасады.
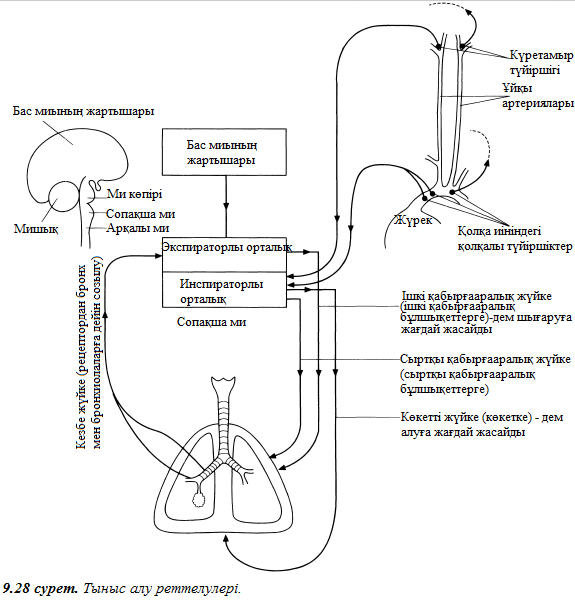
Жиіліктің белгілі шектерінде және тыныс алу тереңдігі еркімен реттей алады, мысалға, куә ретінде, біздің қабілеттілігіміз «тыныс алуды жасырады». Тыныс алудың ерікті реттелуіне біз жылдамдатылған тыныс алуларды, сөйлесіде, әр айтқанда, түскүргенде және жөтелгенде қолданамыз. Бұл жағдайда, бас миының жартышарында туындайтын импулсьтер сәйкес әрекеттерді орындайтын тыныс алу орталығына жіберіледі.
Тыныс алу реттелуі созылу реуепторлары мен хеморецепторлар көмегімен болым кері байланысты мысал ретінде көрсете алады.бас миының жартышарының ерікті белсенділігі бұл механизм әрекетін иеленуге қабілетті.
9.5.3. Өкпе ауасының көлемі мен өкпе сыйымдылығы
Жас ересек әйелдердің өкпелері шамамен орташада 40 л ауаны, ал жас ересек ер адамдарда шамамен, 50 л ауаны араластырады. 9.29 суретінде жас ересек ер адамдардың өкпе жұмыстарының кейбір көрсеткіштері көрсетлген. Әйелдер үшін ол, шамамен, 20 %жалпылама дене массасы айырмашылықтарын қамтып көрсетеді.
Тыныс алу реттелуі созылу реуепторлары мен хеморецепторлар көмегімен болым кері байланысты мысал ретінде көрсете алады.бас миының жартышарының ерікті белсенділігі бұл механизм әрекетін иеленуге қабілетті.
9.5.3. Өкпе ауасының көлемі мен өкпе сыйымдылығы
Жас ересек әйелдердің өкпелері шамамен орташада 40 л ауаны, ал жас ересек ер адамдарда шамамен, 50 л ауаны араластырады. 9.29 суретінде жас ересек ер адамдардың өкпе жұмыстарының кейбір көрсеткіштері көрсетлген. Әйелдер үшін ол, шамамен, 20 %жалпылама дене массасы айырмашылықтарын қамтып көрсетеді.
- Дем алу көлемі – бір дем алу мен дем шығаруды алмастыратын ауа көлемі. Тыныстық күйде адам 450 мл ауаны жұтады және шығарады. Ал максималд физикалық жүктемеде тыныс алу көлемі, шамамен 3 л – ге жетеді.
- 450 мл үстінен адам тағы 1500 мл – ді жұта аладыү бұл қосымша ауа деп аталады.
- Тыныш дем шығарудан кейін ол 1500 мл – дә қосымша шығаруы мүмкін. Бұл – резервті ауа.
- Егер максимал тыныс алуды істеп, сосын максимал тыныс шығаруды істесек, онда дем шығарған ауаның жалпы саны өмірлік өңеш сыйымдылығының шамасын береді.
- Тіпті максимал терең дем шығарудан кейін өкпеде 1500 мл ауа қалады. Оған дем шығару мүмкіндігі болмайды; ол қалдық ауа деп аталады.
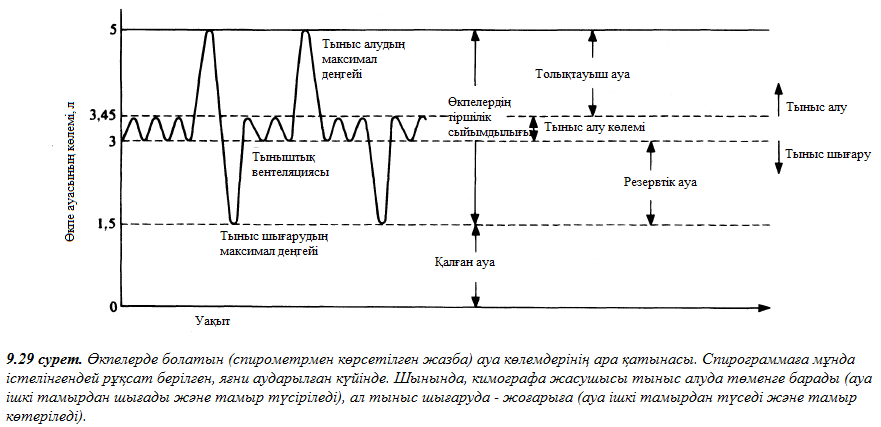
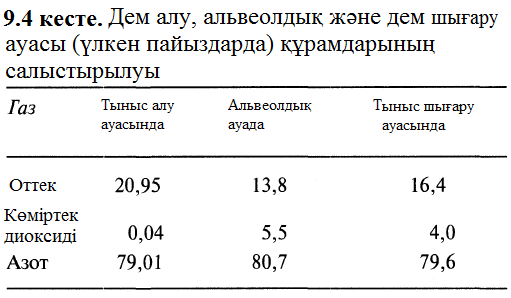
Альвеол ауасымен дем алу және дем шығару ауаларының құрылымын салустыру қызықты (9.4 кесте). Кестеде түскен оттектің 1/5 бөлігін ағза өз қажеттілігі үшін ұстайды, онда СО2 дем шығару саны сияқты тыныс алудан түсетін ауадан 100 есе көбірек санды болады. Қанмен тығыз байланыста альвеол ауасы түседі. Дем алу ауасымен салытырып қарағанда ол оттекті аз үстайды және СО2 – ні көп ұстайды.
9.5.7. Спирометрдің көмегімен тыныс алуды өлшеу
Мектептерде, лабороторияларда және ауруханаларда әдетте, өкпеге түскен ауа көлемін және одан шыққан ауа ауа көлемін спирометрмен өлшейді. Ол сулы тамырдан және оның ауасында болатын сыйымдылығы 6 л – ден кем емес үстіңгі тереңдігіне ауысқан басқа тамырдан тұрады. Бұл екінші тамырдан екі түтікше шығады. Олар өлшейтін кезде ауызға алып сыналатын мүншікпен байланысады. Мүншік клапанмен жабдықталған, сондықтан сынаушы дем алған кезде ауа оған бір түтікшеден кіреді және өзгесінен шығады. Дем шығарудағы бөлінген СО2 – ні сіңіру үшін құрал – жабдыққа натронды әкті орналастырады. (спирометрдегі СО2 концентрадциясының жәй жәйдан өсуі қауыпты болар еді.) Спирометрдің толуы үшін қышқылдық қоспаны («медициналық оттек») қолдануғат болады. Сынаушы тыныс алған кезде өкпесіндегі және спирометрдегі ауабір жабық жүйені құрайды.
Ішкі тамыр теңестірілген және ауа ол жерден шыққан кезде (егер сынаушы дем шығарса) ол сәйкес көтеріледі және түседі. Кимография жазушысы бұл барлық қозғалыстарды айналып жатқан дабылға жазып отырады.
Спирометрге бұл жұмыстың барлық, түп – түгел құрылғылары тіркеледі және бұл жерде біз оны талқыламаймыз. Бірақ, маңызды зат бар, құрылғы анализденрін саралап білу керек және олардан қандай ақпарат алуға болаты туралы өзіндік сұралым алып отыру керек.
Спирометр метоболизм қарқындылығын, дем алу коэфицентін,дем алу көлемін, дем алу жиілігін және оттек қолданысын анықтауға мүмкіндік береді.
Жиілікте дем алу минутына дем алу қозғалыстарының айналымының санын түсінеді. Өкпе вентеляциясы (ӨВ) бұл санды дем алу көлеміне көбейте отырып біледі.
ӨВ = Дем алу жиілігі × Дем алу көлемі
Егер, мысалы, дем алу жиілігі 15/мин – ды құраса, ал дем алу көлемі 400 мл – ді құрайды, сонда ӨВ = 15×400 мл=4000мл/мин (яғни, 1 мин – да сынаушы ағзасы мен орта арасындағы 6 л ауа ауысады).
9.10 неге альвеол вентеляциясы өкпеден аз?
Астында кебір шамалар берілген:
Тыныштықтағы тыныс алу жиілігі – минутына 15 айналым
Максимал жүктемеден кейінгі тыныс алу жиілігі – минутына 40 – 50 айналымдары
Тыныштықтағы ӨВ – минутына 6,75 л
Максимал жүктемеден кейінгі ӨВ (тыныс алу көлем минутына 3 л × тыныс алу жиілігі минутына 45 айналым) – минутына 135 л
Тыныштықта оттектің қолданылуы: минутына 0,25 л – 0,4 л
Максимал жүктемеден кейінгі оттектің қолданылуы минутына – минутына 3,6 л (марафондық дистанциясының айналымында – минутына 5,1 л – ге дейін)
Метоболизм қарқындылығының өлшенуі
9.11. 9.30 суретпен танысыңыздар. Жануар аз болған сайын, метоболизм қарқындылығы сонша жоғары екенін байқау оңай. Бұның себебі неде?
9.12. жануарлардағы метоболизм қарқындылығын өлшемдері арқылы қалай салыстыруға болады?
Сондықтан ағзаның кезкелген метоболистикалық белсенділігінің түрі тыныс алумен тікелей байланысты, онда, тыныс алуды өлшей отырып, біз метоболизм қарқындылығын жеткілікті түрде басқаруға мүмкінік аламыз. Оны оттек қолданысын анықтай отырып, есептеуге болады. Спирометрде ішкі тамыр оттектің сіңіруіне қарай сынаушы ағзасына жай жайдан түседі (СО2 танторды әкте сезіледі, яғни байланыстан шығатынын еске саламыз). Бұл түсірілім берілген кезең уақытына оттек сіңірілуінің өлшемі болып қызмет етеді (9.16 сұраққа қараңыздар, 9.5.9. бөлім).
9.5.7. Спирометрдің көмегімен тыныс алуды өлшеу
Мектептерде, лабороторияларда және ауруханаларда әдетте, өкпеге түскен ауа көлемін және одан шыққан ауа ауа көлемін спирометрмен өлшейді. Ол сулы тамырдан және оның ауасында болатын сыйымдылығы 6 л – ден кем емес үстіңгі тереңдігіне ауысқан басқа тамырдан тұрады. Бұл екінші тамырдан екі түтікше шығады. Олар өлшейтін кезде ауызға алып сыналатын мүншікпен байланысады. Мүншік клапанмен жабдықталған, сондықтан сынаушы дем алған кезде ауа оған бір түтікшеден кіреді және өзгесінен шығады. Дем шығарудағы бөлінген СО2 – ні сіңіру үшін құрал – жабдыққа натронды әкті орналастырады. (спирометрдегі СО2 концентрадциясының жәй жәйдан өсуі қауыпты болар еді.) Спирометрдің толуы үшін қышқылдық қоспаны («медициналық оттек») қолдануғат болады. Сынаушы тыныс алған кезде өкпесіндегі және спирометрдегі ауабір жабық жүйені құрайды.
Ішкі тамыр теңестірілген және ауа ол жерден шыққан кезде (егер сынаушы дем шығарса) ол сәйкес көтеріледі және түседі. Кимография жазушысы бұл барлық қозғалыстарды айналып жатқан дабылға жазып отырады.
Спирометрге бұл жұмыстың барлық, түп – түгел құрылғылары тіркеледі және бұл жерде біз оны талқыламаймыз. Бірақ, маңызды зат бар, құрылғы анализденрін саралап білу керек және олардан қандай ақпарат алуға болаты туралы өзіндік сұралым алып отыру керек.
Спирометр метоболизм қарқындылығын, дем алу коэфицентін,дем алу көлемін, дем алу жиілігін және оттек қолданысын анықтауға мүмкіндік береді.
Жиілікте дем алу минутына дем алу қозғалыстарының айналымының санын түсінеді. Өкпе вентеляциясы (ӨВ) бұл санды дем алу көлеміне көбейте отырып біледі.
ӨВ = Дем алу жиілігі × Дем алу көлемі
Егер, мысалы, дем алу жиілігі 15/мин – ды құраса, ал дем алу көлемі 400 мл – ді құрайды, сонда ӨВ = 15×400 мл=4000мл/мин (яғни, 1 мин – да сынаушы ағзасы мен орта арасындағы 6 л ауа ауысады).
9.10 неге альвеол вентеляциясы өкпеден аз?
Астында кебір шамалар берілген:
Тыныштықтағы тыныс алу жиілігі – минутына 15 айналым
Максимал жүктемеден кейінгі тыныс алу жиілігі – минутына 40 – 50 айналымдары
Тыныштықтағы ӨВ – минутына 6,75 л
Максимал жүктемеден кейінгі ӨВ (тыныс алу көлем минутына 3 л × тыныс алу жиілігі минутына 45 айналым) – минутына 135 л
Тыныштықта оттектің қолданылуы: минутына 0,25 л – 0,4 л
Максимал жүктемеден кейінгі оттектің қолданылуы минутына – минутына 3,6 л (марафондық дистанциясының айналымында – минутына 5,1 л – ге дейін)
Метоболизм қарқындылығының өлшенуі
9.11. 9.30 суретпен танысыңыздар. Жануар аз болған сайын, метоболизм қарқындылығы сонша жоғары екенін байқау оңай. Бұның себебі неде?
9.12. жануарлардағы метоболизм қарқындылығын өлшемдері арқылы қалай салыстыруға болады?
Сондықтан ағзаның кезкелген метоболистикалық белсенділігінің түрі тыныс алумен тікелей байланысты, онда, тыныс алуды өлшей отырып, біз метоболизм қарқындылығын жеткілікті түрде басқаруға мүмкінік аламыз. Оны оттек қолданысын анықтай отырып, есептеуге болады. Спирометрде ішкі тамыр оттектің сіңіруіне қарай сынаушы ағзасына жай жайдан түседі (СО2 танторды әкте сезіледі, яғни байланыстан шығатынын еске саламыз). Бұл түсірілім берілген кезең уақытына оттек сіңірілуінің өлшемі болып қызмет етеді (9.16 сұраққа қараңыздар, 9.5.9. бөлім).
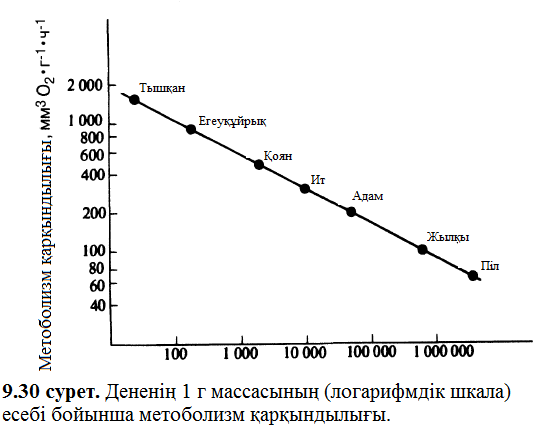
9.5.8. Негізгі алмасу
Негізгі алмасу (негізгі метоболизм) деп заттардың және толық тыныштық немесе ұйқы кезіндегі тек қана тіршілік ұстанымы үшін қажетті энергетикалық шығындардың минималды дәрежедегі алмасуы. Негізгі ауысымды анықтау адам баласы 12-14 сағат ішінде физикалық немесе психикалық тыныштық жағдайында болуы үшін жəне ешқандай өнім қабылдамауын қарауы үшін керек. Бұл сынаушының ішек – қарын жолы өлшемі кезеңінде бос тұруына кепілдік береді. Негізгі ауысым шамасы жасқа, жынысқа, дене массасына жəне өзіндік денсаулық жағдайына тəуелді, сонымен қатар, оның өлшеміне дене бетінің байланысымен арақатынас құрайды.
9.5.9. Дем алу коэффиценті
Келесі теңдеуді қарастырамыз:
Негізгі алмасу (негізгі метоболизм) деп заттардың және толық тыныштық немесе ұйқы кезіндегі тек қана тіршілік ұстанымы үшін қажетті энергетикалық шығындардың минималды дәрежедегі алмасуы. Негізгі ауысымды анықтау адам баласы 12-14 сағат ішінде физикалық немесе психикалық тыныштық жағдайында болуы үшін жəне ешқандай өнім қабылдамауын қарауы үшін керек. Бұл сынаушының ішек – қарын жолы өлшемі кезеңінде бос тұруына кепілдік береді. Негізгі ауысым шамасы жасқа, жынысқа, дене массасына жəне өзіндік денсаулық жағдайына тəуелді, сонымен қатар, оның өлшеміне дене бетінің байланысымен арақатынас құрайды.
9.5.9. Дем алу коэффиценті
Келесі теңдеуді қарастырамыз:

Бұл теңдеуден, тыныс алу кезінде берілген уақытта көміртек қышқылдаунынан тұзілетін көміртек диоксидінің көлемі сол уақытқа сəйкес қажет етілетін оттек көлеміне тең екені көрініп тұр. (Берілген температура мен қысым жағдайларындағы кез келген газ молі сол көлемнің орын алатынын еске саламыз). СО2:О2 байланысы дем алу коэффиценті деп аталады. Көміртектер үшін дем алу коэффиценті 1 тең, яғни.

Осылайша, жоғарыда келтірген теңдеуден
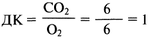
9.13. Трипальмитин майының қышқылдануы тыныс алу үрдісінде келесі теңдеумен жасылады:
2С51Н98О6 + 14О2 → 102СО2 + 98 Н2О
9.14. Трипальмитин үшін ДК не затқа тең?
Этанол мен СО2 - ге дейінгі анаэробты жағдайлардағы глюкозаның қышқылдануындағы ДК не затқа тең?
2С51Н98О6 + 14О2 → 102СО2 + 98 Н2О
9.14. Трипальмитин үшін ДК не затқа тең?
Этанол мен СО2 - ге дейінгі анаэробты жағдайлардағы глюкозаның қышқылдануындағы ДК не затқа тең?
Тыныс алу коэффиценттерін үйрену тыныс алу субстраттары мен метоболизм түрлерінің табиғаты туралы бағалы жинақтар береді (9.5 кестесі).

9.15. Неге адамда әдетте ДК 0,7 ден 1 –ге дейін айналып тұрады?
9.16. 9.31 суретте келтірілген спирограммдардан анықтаңыздар:
А) тыныс алу жиілігі;
Б) тыныс алу көлемі
В) өкпе вентеляциясы
Г) оттек қолданылуы.
9.16. 9.31 суретте келтірілген спирограммдардан анықтаңыздар:
А) тыныс алу жиілігі;
Б) тыныс алу көлемі
В) өкпе вентеляциясы
Г) оттек қолданылуы.
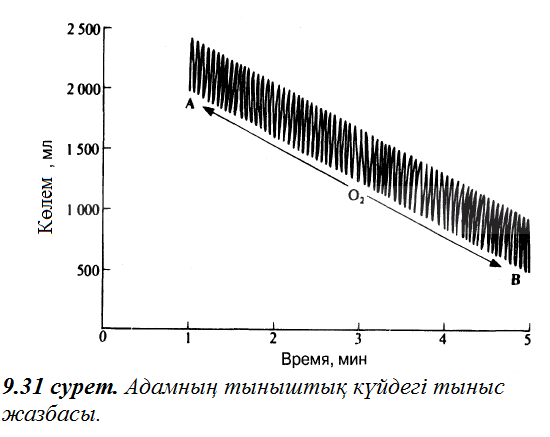
Тәжірибе 9.2. Шағын жер үсті омыртқасыздар оттегі оттегі тұтыну өлшеу,мысалы респирометр көмегімен мокрицтерден
Омыртқасыздардың оттегінің сіңірілуі осы эксперимент жазылады.
Омыртқасыздардың оттегінің сіңірілуі осы эксперимент жазылады.
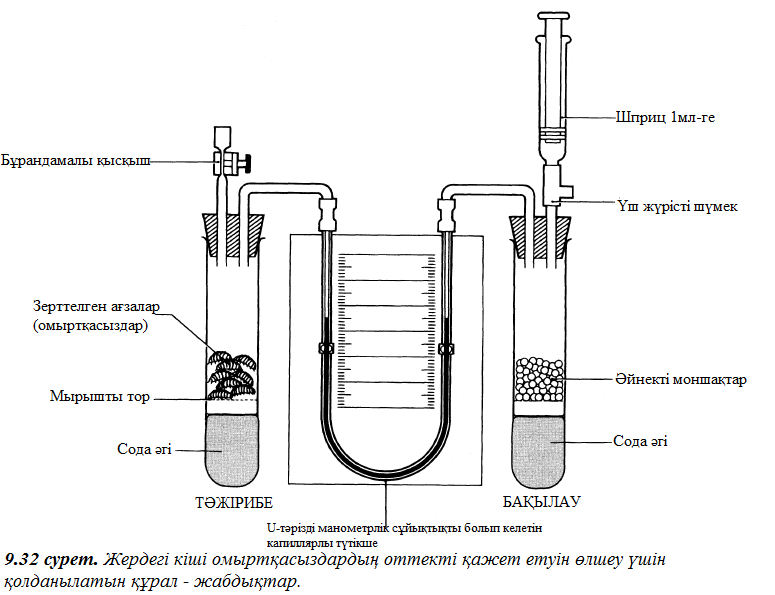
Ескерту
1.Монометриялық сұйықтық ретінде боялған суды,майды немесе сынапты пайдалануға болады.Сұйықтықтың тығыздығы аз болған сайын монометрдегі минискінің ауысу деңгейі артады.
2.Минискінің ауысу деңгейін шкала бойынша анықтайды.Ол үшін U тәрізді монометрдің түтігін шкаласы жазылған қағаз жолағы жабыстырылған тақтайға бекітеміз.Шкаланы монометрдің буындарының біріне жабыстыруға болады.
3.Санауды бастамас бұрын құрылғының нығыздылығын тексеріп алған жөн,ол үшін құрылғыға шприцтің көмегімен ауа жіберіледі,монометриялық сұйықтықтың деңгейі ауысу үшін.Осыдан кейін үш жақты кранды жауып,монометр атмосфералық ауадан өшіреді.Егер құрылғы нығыз болса,монометриялық сұйықтықтың әртүрлілік екі буынында азаймауы қажет.
1.Монометриялық сұйықтық ретінде боялған суды,майды немесе сынапты пайдалануға болады.Сұйықтықтың тығыздығы аз болған сайын монометрдегі минискінің ауысу деңгейі артады.
2.Минискінің ауысу деңгейін шкала бойынша анықтайды.Ол үшін U тәрізді монометрдің түтігін шкаласы жазылған қағаз жолағы жабыстырылған тақтайға бекітеміз.Шкаланы монометрдің буындарының біріне жабыстыруға болады.
3.Санауды бастамас бұрын құрылғының нығыздылығын тексеріп алған жөн,ол үшін құрылғыға шприцтің көмегімен ауа жіберіледі,монометриялық сұйықтықтың деңгейі ауысу үшін.Осыдан кейін үш жақты кранды жауып,монометр атмосфералық ауадан өшіреді.Егер құрылғы нығыз болса,монометриялық сұйықтықтың әртүрлілік екі буынында азаймауы қажет.
9.7. Тыныс мүшелерінің аурулары.
9.7.1. Темекі шегудің өкпе вентиляцияся мен газ алмасуға тікелей әсері.
Шылым шегу тыныс алу мен газ алмасуға әртүрлі әсер етеді.Бұл жерде біз оның қысқа мерзімді салдыры белгілейміз, ал мүмкін салдырды 9.7.2-9.7.5 бөлімдерінде қарастырамыз.
1.Некотин ұсақ бронхиолдарды тарылтады, бұл ауа ағымы мен ауа тасымалдайтын жолға кедергі етеді.
2. Никотин ауа тасымалдайтын жолдан бактериялар мен шаң бөлшектерін кетіретін кірпікшелердің қызметін бұзады. Өзге заттардың тыныс алу жолдарына жиналуы ауаның өту қабілетін ауырлатады.
3. Түтін тітіркендіргіш ретінде әсер етіп, нәтижесінде бокал тәрізді эпителии жасушалары шырышты көбірек мөлшерде секреттейді, ол болса, ауа ағынына кедергі жасайды.
9.7.2. Бронх демікпесі
Демікпе кезінде дем алу қиындығын тегіс бұлшық еттін бронхиолдың қабырғасындағы спазмы туғызады.Бұлшық ет жиырылуынан бронхиолдардын саңылауы тарылады, кейде тіпті мүлдем жабылып қалады.Дем шығару дем алудан қиындай түседі, оған себеп, өкпеден шығатын ауа қысымы одан сайын түтікшелерді қысады. Демікпе шабуылы кезінде тыныс алу ызылдаған дауыс пен ентігу пайда болады. Әсіресе тыныс шығарған кезде айрықша байқалады. Тағы бір жағымсыз белгі – артық жиналған қақырық, қою , жүтел кезінде әрең шығарылады. Ол бронхиолдарда жиналып, одан сайын тыныс алуды қиындата түседі. Қақырықтың ішінде оған түскен бактериялар көбеце алады, ондай жағдайда демікпе бронхитпен күрделене түседі. Демікпе кезіндегі тыныс алу мен бронхтардың шырышты қабығының ісінуін ауырлата түседі.
Демікпе негізіне адам организмінің , сау адамға аса ірі зияны жоқ заттар- түрлі аллергендерге сезгіштігі жоғарлауы жатады. Демікпені жиірек мына аллергендер тудырады: гүл тозаңдары, тұрмыстық шаң(мысалы, оның ішінде кенелер және зең саңырауқұлақтары орналасуы мүмкін), қандай да бір азықтың компоненттері және қаламұшқа толы көпшікте. Приступты эмоцианальді бұзылу тудыруы мүмкін. Сонымен қатар, демікпе приступтарын салқындатылу, физикалық күш немесе шылым шегу тудыруы мүмкін. Біршама қорқатындай жағдай- ол қаладағы балаларда демікпенің шығуының жиілігі. Оның себебі, машиналардың түтіндерінде болатын газдың ауаны ластауы. Ол газдарда демікпе приступтарын тудыратын кейбір заттар болады, сол себепті индустриалды мемлекеттерде заң қаталдана түседі. Бірақ демікпе ол күрделі мәселе, сол себепті оны анықтау үшін жаңа зерттеулер қажет, сол арқылы кімде демікпеге шалдығу қаупі жоғарырақ және оның шығу себебін анықтауға болады. Демікпені емдегенде үлкен пайда тигізетін әдіс-ол қабынуға қарсы дәрі-дәрмектерді қолдану.
9.7.3. Өкпе эмфиземасы
Өкпе эмфиземасы кішкентай альвеолалар қабырғаларының жай жайдан бұзылуымен байланысты. Нəтижесінде өкпеде ауамен толтырылған көлік кеңейеді (9.33 сурет), ал газалмасу жүретіг беттің ауданы тиісінше қысқарады. Науқас одан сайын тұншыға береді. Тіпті ауыр жағдайларда ол үшін бөлменің ішінде жүру қиын болып көрінеді. Креслоға шынжырланған ол қолды дем ала алмай қалмас үшін көтере алатын жағдайда болмайды. Мұндай науқастарда өкпе тіні өзінің иілгіштігін жоғалтады. Сондықтан дем шығару қиындық келтіреді жəне ауаның айтарлықтай үлкен бөлігі дем шығаруда өкпеде қалып қояды. Мұны максимал дем алудан кейін бір уақыт мөлшерінде (əдетте 1с) дем шығаруға болатын ауаның максимал саны – жылдамдатылған көлемді өлшей отырып көрсетіп шығу оңай. (Үлкен бронхиол демікпесінде де бұл көлем қысқартылған). Тыныс алу айтарлықтай щамада өзінің еріксіз сипатын жоғалтады жəне науқас бұл туралы еске алу керек жəне дем алуы үшін күш салуы керек. Сонымен қатар, қабынған үрдіс пен ұсақ бронхиолалардың тарылуы белгіленеді.
9.7.1. Темекі шегудің өкпе вентиляцияся мен газ алмасуға тікелей әсері.
Шылым шегу тыныс алу мен газ алмасуға әртүрлі әсер етеді.Бұл жерде біз оның қысқа мерзімді салдыры белгілейміз, ал мүмкін салдырды 9.7.2-9.7.5 бөлімдерінде қарастырамыз.
1.Некотин ұсақ бронхиолдарды тарылтады, бұл ауа ағымы мен ауа тасымалдайтын жолға кедергі етеді.
2. Никотин ауа тасымалдайтын жолдан бактериялар мен шаң бөлшектерін кетіретін кірпікшелердің қызметін бұзады. Өзге заттардың тыныс алу жолдарына жиналуы ауаның өту қабілетін ауырлатады.
3. Түтін тітіркендіргіш ретінде әсер етіп, нәтижесінде бокал тәрізді эпителии жасушалары шырышты көбірек мөлшерде секреттейді, ол болса, ауа ағынына кедергі жасайды.
9.7.2. Бронх демікпесі
Демікпе кезінде дем алу қиындығын тегіс бұлшық еттін бронхиолдың қабырғасындағы спазмы туғызады.Бұлшық ет жиырылуынан бронхиолдардын саңылауы тарылады, кейде тіпті мүлдем жабылып қалады.Дем шығару дем алудан қиындай түседі, оған себеп, өкпеден шығатын ауа қысымы одан сайын түтікшелерді қысады. Демікпе шабуылы кезінде тыныс алу ызылдаған дауыс пен ентігу пайда болады. Әсіресе тыныс шығарған кезде айрықша байқалады. Тағы бір жағымсыз белгі – артық жиналған қақырық, қою , жүтел кезінде әрең шығарылады. Ол бронхиолдарда жиналып, одан сайын тыныс алуды қиындата түседі. Қақырықтың ішінде оған түскен бактериялар көбеце алады, ондай жағдайда демікпе бронхитпен күрделене түседі. Демікпе кезіндегі тыныс алу мен бронхтардың шырышты қабығының ісінуін ауырлата түседі.
Демікпе негізіне адам организмінің , сау адамға аса ірі зияны жоқ заттар- түрлі аллергендерге сезгіштігі жоғарлауы жатады. Демікпені жиірек мына аллергендер тудырады: гүл тозаңдары, тұрмыстық шаң(мысалы, оның ішінде кенелер және зең саңырауқұлақтары орналасуы мүмкін), қандай да бір азықтың компоненттері және қаламұшқа толы көпшікте. Приступты эмоцианальді бұзылу тудыруы мүмкін. Сонымен қатар, демікпе приступтарын салқындатылу, физикалық күш немесе шылым шегу тудыруы мүмкін. Біршама қорқатындай жағдай- ол қаладағы балаларда демікпенің шығуының жиілігі. Оның себебі, машиналардың түтіндерінде болатын газдың ауаны ластауы. Ол газдарда демікпе приступтарын тудыратын кейбір заттар болады, сол себепті индустриалды мемлекеттерде заң қаталдана түседі. Бірақ демікпе ол күрделі мәселе, сол себепті оны анықтау үшін жаңа зерттеулер қажет, сол арқылы кімде демікпеге шалдығу қаупі жоғарырақ және оның шығу себебін анықтауға болады. Демікпені емдегенде үлкен пайда тигізетін әдіс-ол қабынуға қарсы дәрі-дәрмектерді қолдану.
9.7.3. Өкпе эмфиземасы
Өкпе эмфиземасы кішкентай альвеолалар қабырғаларының жай жайдан бұзылуымен байланысты. Нəтижесінде өкпеде ауамен толтырылған көлік кеңейеді (9.33 сурет), ал газалмасу жүретіг беттің ауданы тиісінше қысқарады. Науқас одан сайын тұншыға береді. Тіпті ауыр жағдайларда ол үшін бөлменің ішінде жүру қиын болып көрінеді. Креслоға шынжырланған ол қолды дем ала алмай қалмас үшін көтере алатын жағдайда болмайды. Мұндай науқастарда өкпе тіні өзінің иілгіштігін жоғалтады. Сондықтан дем шығару қиындық келтіреді жəне ауаның айтарлықтай үлкен бөлігі дем шығаруда өкпеде қалып қояды. Мұны максимал дем алудан кейін бір уақыт мөлшерінде (əдетте 1с) дем шығаруға болатын ауаның максимал саны – жылдамдатылған көлемді өлшей отырып көрсетіп шығу оңай. (Үлкен бронхиол демікпесінде де бұл көлем қысқартылған). Тыныс алу айтарлықтай щамада өзінің еріксіз сипатын жоғалтады жəне науқас бұл туралы еске алу керек жəне дем алуы үшін күш салуы керек. Сонымен қатар, қабынған үрдіс пен ұсақ бронхиолалардың тарылуы белгіленеді.
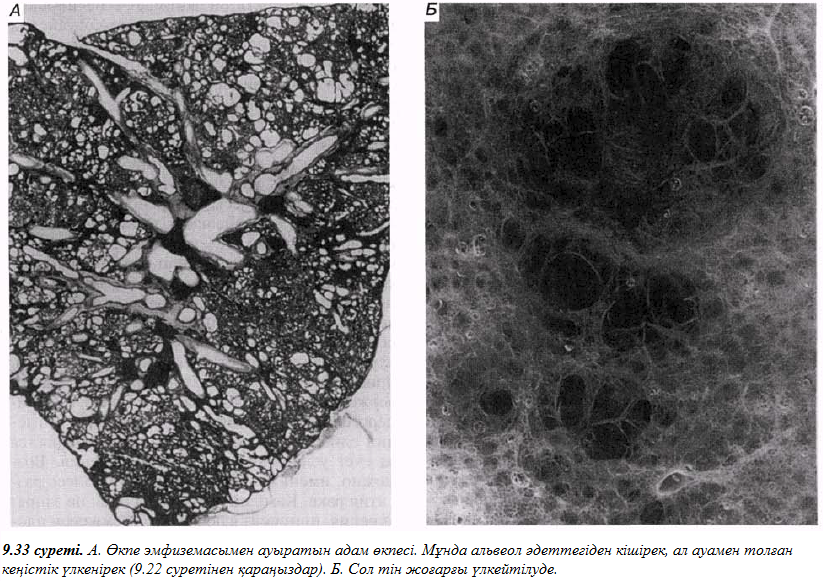
Өкпе эмфиземасының басты себебі-созылмалы тітіркену,өкпеде туындататын темекінің түтіні,атмосфераны ластайтын заттармен немесе шаң бөлшектерімен.Химиялық,сондай-ақ физикалық факторлармен жарақаттайды.Мысалы,темекінің түтініндегі заттар,созылған ұлпаның ыдырау және қайтадан қалпына келудің қалыпты ара қатынасын бұзады.Олар альвеол қабырғасындағы созылғыш ұлпалардың ыдырауын болдырмайтын ферменттерді тежейді және жөндеу процестерін басады.Өкпедегі лейкоциттер күшейіп жатқан стресске жауап ретінде протеолиттік ферменттерді секреттендіреді,сондай-ақ өзінің кезегінде альвеол қабырғасының қиратуына қатысады.Альвеол қабырғасының зақым келуіне мүмкіндік береді және осындай физикалық фактормен-бронхитпен туындаған қатты жөтел.
Ұлыбританияда өкпе эмфиземасы көп таралған.Осы аурумен 100 адамның ішінен 1 адам зардап шегеді,бұған қоса әйелдерден қарағанда,еркектерді 10 есе көп закымдайды.
9.7.4.Бронхит
Бронхит-бастапқы шырышты қабықпен зақымдалған бронхтардың қабыну аурулары. Өткір және созылмалы екі формасын ажыратады. Өткір бронхит қарқынды дамуымен және біршама салыстырмалы тез аяқталуымен сипатталады,әдетте бірнеше күн ішінде. Көбінесе өткір бронхит суық тиюмен қоса болады.
Созылмалы бронхит-ол неғұрлым күрделі мәселе болып табылады. Бұл ауру кеңінен таралған. Ұлыбритания осы созылмалы бронхит ауруынан адамның өлімі жағынан 1 орынды алып отыр. (Жылына 2000нан 1адам,тек шамамен жылына 30000 адам). Науқастар саны шамамен 1 млн-ды құрайды,бұған қоса еркектер әйелдерге қарағанда 3 есе көп. Бронхиттен өлім-жітім шегетіндердің арасында 6 есе жоғары. Бронхит жиі өкпе эмфиземасына алып келеді,әсіресе одан кейінгі сатыларында. Сондай-ақ эмфизема сияқты,ол негізінен темекі шегумен байланысты және ауаның ластануының аз дәрежесімен байланысты. Оған сондай-ақ газ алмасаудың қысқарылуынан ентігу сезімі тән. Темекі түтінінде кездесетін шайырлар ең біріншіден бронхтың шырышты қабықшасының қабынуына жауапты. Бронхиттің белгілерінің бірі-тітіркенуге жауап ретінде бокал тәрізді эпителия жасушасынан күшейтілген шырыштың бөлінуі.Темекі шегетіндерде бронхтардан шырышты айдайтын кірпіктері өледі немесе сал болады. Ең басты себеп жөтел болып табылады,онда артық шырыш бөлінеді және уақыт өте жуан және сары жасылдау түске айналады. Сол шамада ауру барысында жөтел күшейеді және тыныс алу барған сайын қиынға соғады. Тыныс алу функциясы неғұрлым бұзылған сайын,осы тұрғыда басқа да аурулардың пайда болуына ықтималдылығы жоғары,атап айтқанда пневмония.
9.7.5. Өкпе обыры
Ұлыбританияда көбіне обырдың,ер адамдарда-өкпе обыры(әйелдерде сүт безінің обыры) формасы дамыған. Ол Ұлыбританияда өлімнің негізгі себептерінің арасында жүрек-қан тамырлары және инсульттан кейін үшінші орын алады (9.34сурет). Мұнда XX ғасырдың 80 жылдары өкпе обырынан көз жұматындар шамамен 18ден 1і (5-6%)болған. Бақыланбайтын жасушалардың бөлініп, ағзаға түсетін қалыпты сигналдарға жауап қайтаруын тоқтатуынан пайда болады. Соның нәтижесінде дифференцияланбаған салмақ туындайды (ісік;9.35сурет). Кей кезде жасушалар осы салмақтан алыстайды және дененің басқа бөліктеріне(қанға немесе лимфаға) ауысып екінші ісікке бастама болады. Өкпе обыры бастапқыда туындайды,ереже бойынша,эпителий бронхиолындағы бронхиалды карцинома деп аталатын түрінде және содан кейін осы жерден ол өкпеге таралады. Себебі болып көбінесе темекі шегу болып табылады(өкпе обырынан көз жұматындардың 99,75% шылым шеккіштер). Осыған жауапты темекі түтінінде кездесетін- шайыр. Олардың құрамында обырға себеп болатын компоненттер бар(канцерогендер). Шырышты эпителийлі тыныс алу жолдары тітіркеніп,жасушалардың бөлінуінің күшеюі есебінен қалыңдайды. Обырдың даму процесіне мүмкін дәл осы ұшыратады. Шылым шегуге тыйым салуға бағытталған компаниялар соңғы уақытта өз жемісін көрсетуде: өкпе обырынан көз жұматындар саны біртінддеп азаюда (9.34сурет).
Ұлыбританияда өкпе эмфиземасы көп таралған.Осы аурумен 100 адамның ішінен 1 адам зардап шегеді,бұған қоса әйелдерден қарағанда,еркектерді 10 есе көп закымдайды.
9.7.4.Бронхит
Бронхит-бастапқы шырышты қабықпен зақымдалған бронхтардың қабыну аурулары. Өткір және созылмалы екі формасын ажыратады. Өткір бронхит қарқынды дамуымен және біршама салыстырмалы тез аяқталуымен сипатталады,әдетте бірнеше күн ішінде. Көбінесе өткір бронхит суық тиюмен қоса болады.
Созылмалы бронхит-ол неғұрлым күрделі мәселе болып табылады. Бұл ауру кеңінен таралған. Ұлыбритания осы созылмалы бронхит ауруынан адамның өлімі жағынан 1 орынды алып отыр. (Жылына 2000нан 1адам,тек шамамен жылына 30000 адам). Науқастар саны шамамен 1 млн-ды құрайды,бұған қоса еркектер әйелдерге қарағанда 3 есе көп. Бронхиттен өлім-жітім шегетіндердің арасында 6 есе жоғары. Бронхит жиі өкпе эмфиземасына алып келеді,әсіресе одан кейінгі сатыларында. Сондай-ақ эмфизема сияқты,ол негізінен темекі шегумен байланысты және ауаның ластануының аз дәрежесімен байланысты. Оған сондай-ақ газ алмасаудың қысқарылуынан ентігу сезімі тән. Темекі түтінінде кездесетін шайырлар ең біріншіден бронхтың шырышты қабықшасының қабынуына жауапты. Бронхиттің белгілерінің бірі-тітіркенуге жауап ретінде бокал тәрізді эпителия жасушасынан күшейтілген шырыштың бөлінуі.Темекі шегетіндерде бронхтардан шырышты айдайтын кірпіктері өледі немесе сал болады. Ең басты себеп жөтел болып табылады,онда артық шырыш бөлінеді және уақыт өте жуан және сары жасылдау түске айналады. Сол шамада ауру барысында жөтел күшейеді және тыныс алу барған сайын қиынға соғады. Тыныс алу функциясы неғұрлым бұзылған сайын,осы тұрғыда басқа да аурулардың пайда болуына ықтималдылығы жоғары,атап айтқанда пневмония.
9.7.5. Өкпе обыры
Ұлыбританияда көбіне обырдың,ер адамдарда-өкпе обыры(әйелдерде сүт безінің обыры) формасы дамыған. Ол Ұлыбританияда өлімнің негізгі себептерінің арасында жүрек-қан тамырлары және инсульттан кейін үшінші орын алады (9.34сурет). Мұнда XX ғасырдың 80 жылдары өкпе обырынан көз жұматындар шамамен 18ден 1і (5-6%)болған. Бақыланбайтын жасушалардың бөлініп, ағзаға түсетін қалыпты сигналдарға жауап қайтаруын тоқтатуынан пайда болады. Соның нәтижесінде дифференцияланбаған салмақ туындайды (ісік;9.35сурет). Кей кезде жасушалар осы салмақтан алыстайды және дененің басқа бөліктеріне(қанға немесе лимфаға) ауысып екінші ісікке бастама болады. Өкпе обыры бастапқыда туындайды,ереже бойынша,эпителий бронхиолындағы бронхиалды карцинома деп аталатын түрінде және содан кейін осы жерден ол өкпеге таралады. Себебі болып көбінесе темекі шегу болып табылады(өкпе обырынан көз жұматындардың 99,75% шылым шеккіштер). Осыған жауапты темекі түтінінде кездесетін- шайыр. Олардың құрамында обырға себеп болатын компоненттер бар(канцерогендер). Шырышты эпителийлі тыныс алу жолдары тітіркеніп,жасушалардың бөлінуінің күшеюі есебінен қалыңдайды. Обырдың даму процесіне мүмкін дәл осы ұшыратады. Шылым шегуге тыйым салуға бағытталған компаниялар соңғы уақытта өз жемісін көрсетуде: өкпе обырынан көз жұматындар саны біртінддеп азаюда (9.34сурет).
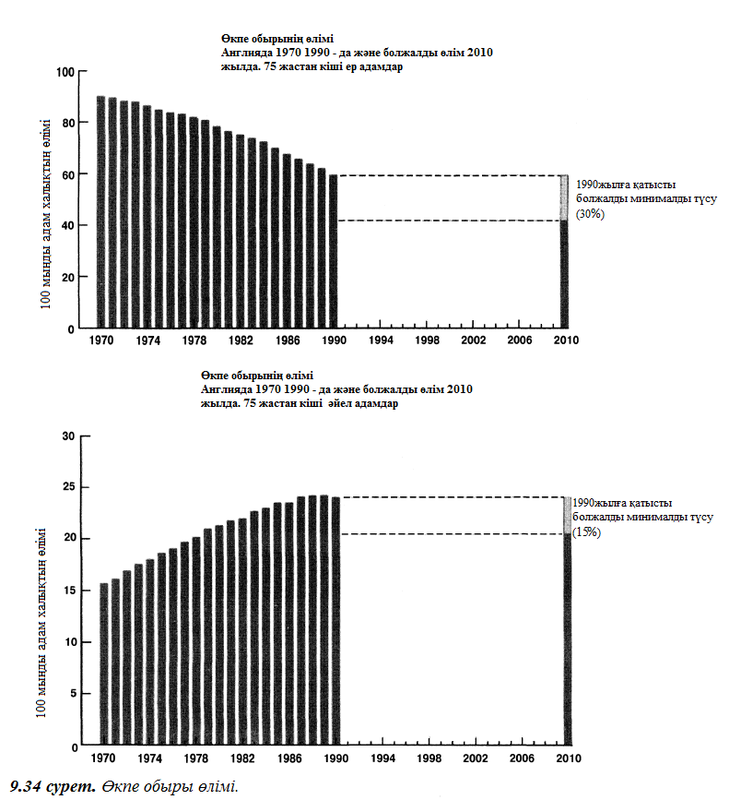
Дем алу жүйесі жұмысының тиімділігі есейе келе құлдырайды. Иілгіш тін жиілеп жоғалады, ал көкірек жасушасы еңбекпен созылады. Нəтижесінде, өкпе сыйымдылығы (максимал дем алудан кейін максимал дем шығаруға болатын максимал ауа көлемі) көрсеткіші түседі.
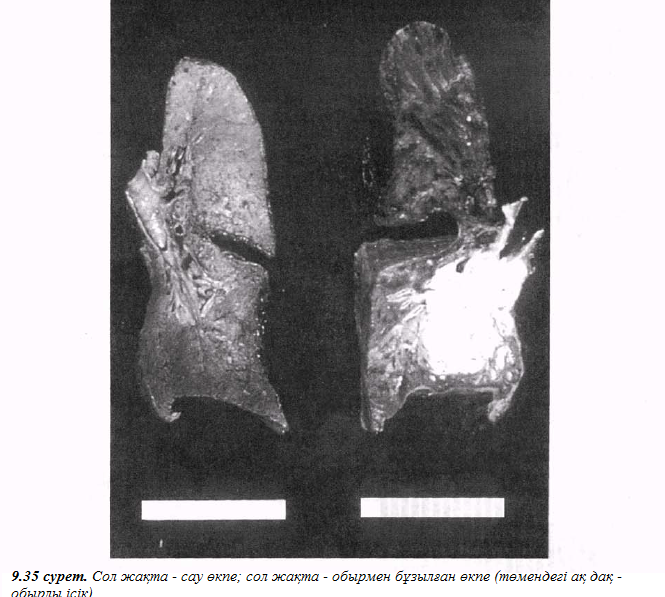
70 жастарда өкпенің тіршілік сыйымдылығы 35% - ға кемуі мүмкін. Барлық дем алу жүйелерінің жұмысында жас іс жүзінде былай айтылады: əлсірейді, атап айтқанда, эпителий кірпікшесінің соққысы мен өзінің лейкоцит қорғаныш функциясын əлсіздеу орындайды. Бұл барлығы тыныс алу жүйесін пневмония, бронзит жəне өкпе эмфиземасы сияқты аурулар шамасын көтере отырып, осал қылып тастайды.
12 бөлім
БИОТЕХНОЛОГИЯ ЖӘНЕ МИКРОБИОЛОГИЯ
Микробиология дегеніміз не?
Микробиология – микроорганизмдер туралы ғылым, яғни олар соншалықты аз және оларды тек микроскоп көмегімен ғана көруге болады. Оған: қарапайым және микроскопиялық балдырлар сияқты бактериялар, вирустар, саңырауқұлақтар жатады. Жіктеу және осы ағзалардың кейбір негізгі сипаттамаларының мысалдары 2 бөлімде келтірілген.
Микроағзалар өте алуан түрлі болып табылады және адам пайдалану үшін зор әлеуеті бар. Қолайлы жағдайында, олар тез химиялық қосылыстардың әсерінен тез өседі және көбейеді. Бұл әмбебап болып табылады және оларға соншалықты пайда әкеледі. Гендік инженерия көмегімен оларға толтырып өзіндік емес пайдалы өнімдер беруге болады, мысалы инсулин. Микроорганизмдерді адамдарға пайдалану енді басталып жатырса да, жетістіктері аз емес. Адамның игілігі үшін микроорганизмдерді қабылдау және басқа да биологиялық объектілерді пайдалану- бұл, бір сөзбен айтқанда – БИОТЕХНОЛОГИЯ
Биотехнология дегеніміз не?
Биотехнология - өнеркәсіптік өндірістерінде ағзалардың және биологиялық жүйелерде немесе биологиялық процестердің пайдаланылуы болып табылады. Биотехнологияда тек микроорганизмдер ғана пайдаланылмайды. Шын мәнінде, бойында биологиялық процестері бар кез келген өнімді биотехнология ретінде қарастыруға болады. Бұған гендік инженерия және ауыл шаруашылық өсімдіктер мен жануарлардың клондау қамтуы мүмкін. Биотехнология мысалдары 12.1. суретте көрсетілген.
Биотехнология тек адамға маңызды өнімдер алуға мүмкіндік бере бермейді, мысалы этил спирті, сыра және гормондық инсулин. Биотехнологияға мысал ретінде биосенсорлары бар биотехнологиялар: ағын суларды тазарту, қатты тұрмыстық қалдықтарды немесе ластануды анықтауды жатқызуға болады. Мұнда, шик3заттан гөрі процесс маңыздырақ болып табылады.
Биотехнологияда тірі организмдерді адамның керегі үшін кеңінен пайдалыналады. Сол себепті биотехнологияны ауыл шаруашылық жануарлардың жаңартылу бөліміне жатқызуға болады.
Мысалы, ірі қара малдарды және шошқаларды, сонымен бірге бидай және картоп өнімдерін жатқызуға болады. Осы бағыттарда маңызды рөлді гендік инженерия дәл және тез 8табады.
Гендік инженерия
Мысалы, адамның бойын өсіретін соматотропин гормонымен, инсулин гормоны, гербицилам пециститіне тұрақты.
Микробиология – микроорганизмдер туралы ғылым, яғни олар соншалықты аз және оларды тек микроскоп көмегімен ғана көруге болады. Оған: қарапайым және микроскопиялық балдырлар сияқты бактериялар, вирустар, саңырауқұлақтар жатады. Жіктеу және осы ағзалардың кейбір негізгі сипаттамаларының мысалдары 2 бөлімде келтірілген.
Микроағзалар өте алуан түрлі болып табылады және адам пайдалану үшін зор әлеуеті бар. Қолайлы жағдайында, олар тез химиялық қосылыстардың әсерінен тез өседі және көбейеді. Бұл әмбебап болып табылады және оларға соншалықты пайда әкеледі. Гендік инженерия көмегімен оларға толтырып өзіндік емес пайдалы өнімдер беруге болады, мысалы инсулин. Микроорганизмдерді адамдарға пайдалану енді басталып жатырса да, жетістіктері аз емес. Адамның игілігі үшін микроорганизмдерді қабылдау және басқа да биологиялық объектілерді пайдалану- бұл, бір сөзбен айтқанда – БИОТЕХНОЛОГИЯ
Биотехнология дегеніміз не?
Биотехнология - өнеркәсіптік өндірістерінде ағзалардың және биологиялық жүйелерде немесе биологиялық процестердің пайдаланылуы болып табылады. Биотехнологияда тек микроорганизмдер ғана пайдаланылмайды. Шын мәнінде, бойында биологиялық процестері бар кез келген өнімді биотехнология ретінде қарастыруға болады. Бұған гендік инженерия және ауыл шаруашылық өсімдіктер мен жануарлардың клондау қамтуы мүмкін. Биотехнология мысалдары 12.1. суретте көрсетілген.
Биотехнология тек адамға маңызды өнімдер алуға мүмкіндік бере бермейді, мысалы этил спирті, сыра және гормондық инсулин. Биотехнологияға мысал ретінде биосенсорлары бар биотехнологиялар: ағын суларды тазарту, қатты тұрмыстық қалдықтарды немесе ластануды анықтауды жатқызуға болады. Мұнда, шик3заттан гөрі процесс маңыздырақ болып табылады.
Биотехнологияда тірі организмдерді адамның керегі үшін кеңінен пайдалыналады. Сол себепті биотехнологияны ауыл шаруашылық жануарлардың жаңартылу бөліміне жатқызуға болады.
Мысалы, ірі қара малдарды және шошқаларды, сонымен бірге бидай және картоп өнімдерін жатқызуға болады. Осы бағыттарда маңызды рөлді гендік инженерия дәл және тез 8табады.
Гендік инженерия
Мысалы, адамның бойын өсіретін соматотропин гормонымен, инсулин гормоны, гербицилам пециститіне тұрақты.
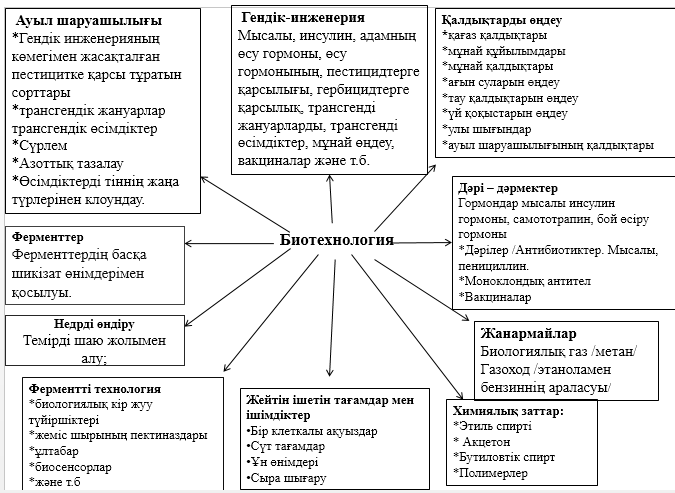
12.1. Даму қажеттіліктері:
Ғалымдар ғылым жолында бірінші рет 19 – ғасырдың отрасында бактериялармен саңырауқұлақтарды бақылап отырып, өсіріп басталды. Бұл жұмыстарды жүргізген Парижден Луи Пастер, Берлиннен Роберт Кох. Ғалымдар өздері ойлап тауып, өсіріп жатқан организмдердің маңызы зор екенін түсінді. Бұл үшін әртүрлі өсіру жолдары және микроорганизмдерің түр – түрінен, бір – бірінен айырып білу керек болды. Мысалы, микроорганизмдерді өсіру жолдарына, соның ішінде паразиттік микроорганизмдерді өсіру үшін қиын /шіріген, ылғалды/ орталар керек болды.
12.1.1. Бізге керекті өнімдер:
Өсу үшін және көбею үшін неше түрлі керекті заттар бар. Сонымен бірге, қуат көзі керек. Микроорганизмдерді өсіру үшін қолайлы жағдай керек. Кез келген қолайлы жағдай мына заттарға толуы қажет:
Көміртек көзі . Көптеген бактериялар мен саңырауқұлақтар гетероторфтарға жатады. Олар көміртек көзімен қанықтырылуы қажет. Мұндай көздерге, әдетте, глюкоза, органикалық тұздар. Мысалы, натрий ацетаты жатады. Бірақ бактериялар жалпы кең спекторлық органикалық заттардан, сонымен бірге, майлы оттектерден, спирттардан, көміртегіден және метаннан тұрады. Кейбір ішегімізде жүрген бактериялар мен саңырауқұлақтар целлюлоза сіңіре алады. Сол себепті, оларды көміртек көзі ретінде пайдалана алады. Барлық ауру туғызатын бактериялар гетеротрофтарға жатады. Кейбір балдырлар мен бактериялар. Мысалы, цианобактерия автотрофтар болып саналады. Сонымен бірге, балдырлар фотосинтездік организмге жатады. Сонда олардың ішінде фотосинтездейтін бактериялар кездеседі.
Азот көзі – бұл органикалық заттар, мысалы, аминқышқылдары, пептидтер және ақуыздар, органикалық емес заттар: аммония тұзымен нитраттар жатады. Аминқышқылдарын қайнатылған ақуызға сұйық түрінде қосылады. Бұлар пептондар деп аталады.
Витаминдер мен бой өсіру микроорганизмдерін өсіру үшін органикалық заттар, азғантай мөлшерде қосылады. Оларға В тобындағы витаминдер жатады(тиамин, немесе В6 рибофлавин, немесе В2 ниоцин, немесе В3 және В6 ) және фалиқышқылы жатады. Пурин, пиримидин сияқты органикалық заттар қосады.
Микрооорганизмдерді жай ғана субстратқа қарағанда синтездік факторына қарап айыруға болады. Егер микроорганизмдер жеткілікті түрде бой жағдайы қажеттіліктеріне қатты бақылайтын болсақ, біз оларға бой қсіру үшіе лабароторияда негізгі субстраттарды дайындай аламыз, осындай жердерде олар қалыпты өседі.
Минералды тұздар
Бой өсіру үшін көбіне иондық кальциялер, калий, натрий, феррум және магний сонымен бирге хлорид, фосфат, сульфат +керек. Жоғарыда айтылғандай, азот аммони немесе нитрат түрінде болады.
Балдырлар өсуі үшін талаптар (қараңыз өсімдіктердің өсуін күшейту үшін бірдей. Кесте туралы.
7.7 және 7.8).
12.1. Осы элементтердің әрқайсысының рөлі бір мысалын келтір.
Гетеротрофтық бактерияларға 1) темір және фосфор;
2) магний оксиді мен ● автотрофты бактериялар. (Егер қажет болса, таблицасын пайдаланыңыз. 7.7).
Қуат көзі
Энергетикалық қоректік жасушалар жайында бастапқы 7 бөлімде талқыладық. Химиялық қуат және жарық қуат ретінде жасаланылады. Химиялық қуат көзін қадет еткен органикалық хемотрофтық деп аталады. Ал жарық қуатын қолданылатын фототрафтық жіне фотосиндездік деп аиталады. /2.3 кесте/
Фотосинтездік микроорганикалық балдырлар мен кейбір бактериялар. Мысалы, цианобактериялар жатады. Егер химиялық қуат күші керек болса, қант түрінде мысалы, глюкозаны қолданады.
Су тура мағынада оның қоректік зат екенін есепке алмайтын болсақ,
Су бүкіләлемге сіңіру үшін тікелей керекті зат. Мысалы, бактерияларға көбіне ылғал керек, ашытқыларға одан да көп керек. Мысалы, қарапайым сіңірілімділік үшін керек /12.1. кестесін қолданамыз./
Ғалымдар ғылым жолында бірінші рет 19 – ғасырдың отрасында бактериялармен саңырауқұлақтарды бақылап отырып, өсіріп басталды. Бұл жұмыстарды жүргізген Парижден Луи Пастер, Берлиннен Роберт Кох. Ғалымдар өздері ойлап тауып, өсіріп жатқан организмдердің маңызы зор екенін түсінді. Бұл үшін әртүрлі өсіру жолдары және микроорганизмдерің түр – түрінен, бір – бірінен айырып білу керек болды. Мысалы, микроорганизмдерді өсіру жолдарына, соның ішінде паразиттік микроорганизмдерді өсіру үшін қиын /шіріген, ылғалды/ орталар керек болды.
12.1.1. Бізге керекті өнімдер:
Өсу үшін және көбею үшін неше түрлі керекті заттар бар. Сонымен бірге, қуат көзі керек. Микроорганизмдерді өсіру үшін қолайлы жағдай керек. Кез келген қолайлы жағдай мына заттарға толуы қажет:
Көміртек көзі . Көптеген бактериялар мен саңырауқұлақтар гетероторфтарға жатады. Олар көміртек көзімен қанықтырылуы қажет. Мұндай көздерге, әдетте, глюкоза, органикалық тұздар. Мысалы, натрий ацетаты жатады. Бірақ бактериялар жалпы кең спекторлық органикалық заттардан, сонымен бірге, майлы оттектерден, спирттардан, көміртегіден және метаннан тұрады. Кейбір ішегімізде жүрген бактериялар мен саңырауқұлақтар целлюлоза сіңіре алады. Сол себепті, оларды көміртек көзі ретінде пайдалана алады. Барлық ауру туғызатын бактериялар гетеротрофтарға жатады. Кейбір балдырлар мен бактериялар. Мысалы, цианобактерия автотрофтар болып саналады. Сонымен бірге, балдырлар фотосинтездік организмге жатады. Сонда олардың ішінде фотосинтездейтін бактериялар кездеседі.
Азот көзі – бұл органикалық заттар, мысалы, аминқышқылдары, пептидтер және ақуыздар, органикалық емес заттар: аммония тұзымен нитраттар жатады. Аминқышқылдарын қайнатылған ақуызға сұйық түрінде қосылады. Бұлар пептондар деп аталады.
Витаминдер мен бой өсіру микроорганизмдерін өсіру үшін органикалық заттар, азғантай мөлшерде қосылады. Оларға В тобындағы витаминдер жатады(тиамин, немесе В6 рибофлавин, немесе В2 ниоцин, немесе В3 және В6 ) және фалиқышқылы жатады. Пурин, пиримидин сияқты органикалық заттар қосады.
Микрооорганизмдерді жай ғана субстратқа қарағанда синтездік факторына қарап айыруға болады. Егер микроорганизмдер жеткілікті түрде бой жағдайы қажеттіліктеріне қатты бақылайтын болсақ, біз оларға бой қсіру үшіе лабароторияда негізгі субстраттарды дайындай аламыз, осындай жердерде олар қалыпты өседі.
Минералды тұздар
Бой өсіру үшін көбіне иондық кальциялер, калий, натрий, феррум және магний сонымен бирге хлорид, фосфат, сульфат +керек. Жоғарыда айтылғандай, азот аммони немесе нитрат түрінде болады.
Балдырлар өсуі үшін талаптар (қараңыз өсімдіктердің өсуін күшейту үшін бірдей. Кесте туралы.
7.7 және 7.8).
12.1. Осы элементтердің әрқайсысының рөлі бір мысалын келтір.
Гетеротрофтық бактерияларға 1) темір және фосфор;
2) магний оксиді мен ● автотрофты бактериялар. (Егер қажет болса, таблицасын пайдаланыңыз. 7.7).
Қуат көзі
Энергетикалық қоректік жасушалар жайында бастапқы 7 бөлімде талқыладық. Химиялық қуат және жарық қуат ретінде жасаланылады. Химиялық қуат көзін қадет еткен органикалық хемотрофтық деп аталады. Ал жарық қуатын қолданылатын фототрафтық жіне фотосиндездік деп аиталады. /2.3 кесте/
Фотосинтездік микроорганикалық балдырлар мен кейбір бактериялар. Мысалы, цианобактериялар жатады. Егер химиялық қуат күші керек болса, қант түрінде мысалы, глюкозаны қолданады.
Су тура мағынада оның қоректік зат екенін есепке алмайтын болсақ,
Су бүкіләлемге сіңіру үшін тікелей керекті зат. Мысалы, бактерияларға көбіне ылғал керек, ашытқыларға одан да көп керек. Мысалы, қарапайым сіңірілімділік үшін керек /12.1. кестесін қолданамыз./

Қоршаған ортаның өзгерістері
Сіңірімді сіңу үшін қажетті ортадан басқа бактерияларды өсіру үшін қоршаған ортаның қажетті жағдайын туғызу керек. Микроорганизм белгілі бір температурада жақсы өседі. Дәл сол жағдайда оларға мөлшерлі көміртегімен жарығы түсуі керек. Әртүрлі микроорганизмдерге әртүрлі қоршаған орта керек.
Температура
Температуралық жағдайдың әртүрлі организмдерге керектігі әртүрлі температураның ферменттерге тигізетін әсерін оңай қарап отыруға болады . Барлық жасушалар ферменттерден тұрады, ферментердің жасушаларының жұмыс жылдамдығы жасушалар өсіру жылдамдығы температураға байланысты. Әрбір микроорганизмге қажетті әртүрлі диапозондағы температура қажет.
Ең жоғарғы температура денатурациядан басталады. Денатурация ақуыздардың өмірлік жолын бастауының бақылауы. Оптималдық температураның өзгеру 0-5 °С аралығында болады, кейбір бактериялар үшін мұхиттарда 70°С дейін керек болады. Кейбір терең су астындағы бактрериялар вулкандық жағдайда өсетін бактериялар 118°С тірі болады.
Температураға қарай 4 топқа бөлуге болады.
Термофилдер, бұлар 45°С та өседі, мысалы, лактобацилустың кейбр түрлері(сүтте кездеседі).
Мезофилдер, өсу үшін қажетті температура 25°С-45°С. Сүтқоректілердің есебінен өмір сүретін бактериялар үшін қолайлысы 37°С, ал адам ішектерінде мекендеуші, мысалы Escherichiacoli(Е. coli)үшін максимум 42—43 °С қажет. Сонымен қатар, ашытқылар немесе саңырауқұлақтар да мезофилдерге жатады.
Психрофильдер, температура 15°С тан төмен, және олар 20°Стан жоғары температурада өспейді.
Бұл микроорганизмдерді тоңазытқышта сақтау өте қиын. Әсіресе, осы микроорганизмдер тамақтардың бұзылуына әкеліп соқтырады.
Психротрофтар, температуралық ортасы 15°С, бірақ олар одан төмен температурада өседі, мысалы, 5°С немесе одан да төмен. Candida түрінен келетін кейбір ашытқылар, кейбір көгерген саңырауқұлақтар, міне дәл осылар тоңазытқышты сақталатын тамақтарды бүлдіреді.
рН
Температура жағдайындағыдай әрбір түрі өзінің төменгі, жоғарғы және дәл температурасында өседі. Барлық бактериялар өзінің орта температурасында болады. Қышқыл ортада жақсы өсетін бактериялар микроорганизмдер ацидифиломдар деп аталады. Ал ылғал, дымқыл ауада бактериялар алколофилдер деп аталалды. Көптеген ашытқылар 4,5-5,0 рҺ орта ауасында өседі. Ал көктейтін саңырауқұлақтарға аз қышқыл орта керек.
Ауаның концентрациясы
Ауаның құрамынан жұту құрамына қарай микроорганизмдер келесі топқа бөлінеді:
*Облигатты аэробы. Мұнда микроорганизмдер тек көміртегімен өседі бұған туберкулездің бактериялары жатады, балдырлар мен қарапайымдылар тобындағы микроорганимдер жатады.
*Факультативтық аэробтар. Мұндай топтарға көміртекпен өсетін микроорганизм көміртегі жоө кезде анаэробты өсуін айтады. Оған бактериялар, ашытқылар жатады.
* Қамту анаэробы. Бұған ішек жолындағы қыртыстарда өсетін микроорганизмдер жатады. Оған сіреспе, ботулизм ауруын туғызуы мүмкін. Сонымен бірге уланып ауруы мүмкін.
*Микроаэрофилдер. Бұған өте төмен температурада өсетін микроорганизмдер жатады. Мысаы, сүтте кездесетін лактобоцилоз
4 бөлік түтіктің әр бөлімінде
*аэробтар
*анаэробтар
*факультативті аэробтар
*микроаэрофилдер
Иондық және осматикалық теңдік
Кейде жасушалардың ішкі құрамын концентрацияда ол жаймен әрі қарай осмос жолымен суын жоғалтады. Соған қарамастан, голофил деген организмдер өте жоғары концентрациялық тұздарда мысалы, үлкен Тұзды көл Америкада, Өлі көл Израилда кездеседі. Және оған қарама – қарсы суды кетіре алмайтын сулы ортада өсетін жасушаларда жоқ микроорганизмдде кездеседі. Соған қарамастан, олар дұрыс тепе – теңдік құрайды.
Дақылды орталары
Орта қатты және сұйық субстанцияны айтады. Дарақ деп осы микроб жасушаларының жиынтығының орта ішіндегі немесе орта сыртындағы өсуін айтады.
12.2.1. Қатты және сұйық орталар
Микроорганизмдерді қатты ортада немесе сұйық ортада өсіреді.
Қатты орта
Қатты орта өсіп тұрған өсімдіктер мен саңырауқұлақтар үшін өте ыңғайлы. Оларды гель образдағы 1-2 % концентрациясы бар сұйық қоректік ерітіндімен араластыру арқылы дайындайды. Сөйтіп, қоректік агарды алады. Агар 90—100 С температурада балқиды, ал 44°С қатады. Микроорганизмдерді беріктендіру үшін агар қосады.
Агар ашық түсті. Кейін полисахаридте айналған соң микроорганизм бұщылмау үшін төзімді болып табылады. Қатты және сұйық түріндегі мысалдар төмендегі бөлімдерде көрсетіледі.
Сұйық орта
Сұйық ортаны көбіне жарық санының өсуін зерттеу үшін қолданылады. жасушаларды қақпағы мықты жабылған пробиркаға салады. Қолдану үшін жасушалар алдын – ала спиральдинген болуы керек. Азғантай жасушалардың қосылуын бұл орталарда өсім дейді. Одан кейін өсіндерді термостат арқылы ыңғайлы ортада кездейсоқ жағдайлас отмргедилл береді. Ортаны көп шамада қолданған кезде тұнба қалмас үшін магнит арқылы араластырады. Ауа тазалануы үшін ауа тазалағыш түтіктерді тазалап тұрады. Ауа шашыратқыш құрал арқылы көптеген кішкентай түтіктерден еркін өтеді.
Ортаны көп шамада қолданған кезде тұнба қалмас үшін магнит арқылы отырады. Одан басқа оттектің норма жүйесіндегі концентрациясын сақтау үшін, орта арқылы тап - таза ауа кіреді. Б ұ л үшін әртүрлі құралдар , яғни сүзбе және әйнекті сүзбе , немесе сіңірілген мақталы вата қолданылады. Ауа тозаңдатқыш , я ғни көптеген кішкетай тесіктер арқылы өтеді. Олар мінсіз көпіршіктер алуға мүмкіндік береді. Оны дақыл ыдысына бара жатқан кекзінен, түтіктің соңына қарай бекітеді.
12.2.2. Байытылған селективтік орта
Байытылған орта бұл белгілі бір микроорганизмді қажетті қорлармен толтырырылуы. Сол себепті басқа микроорганизмдерге қарағанда, бұлардың өсімі жоғары болады. Ортаны түрлі дақылмен толтыру барысында табиғатта керекті микроорганизмдердің неше түрлерін шығаруға болады. Және олар үшін рН, оттек көзі, қуат көзі, температура барлығы жоғары нәтижеде қаралу керек. Мысалы, ТИФ ауруын туғызатын Salmonella фехалий ортасымен араласа отырып, селендік микроорганизмді құрайды. Фехалийде кәдімгі ішек бактериялары да болады, олар селенмен жойылады.
Селективті орта – бірен саран, ортадан басқа ортаның көбінің өсуін тежейді. Бұған мысал келтіретін болсақ, дақылға пеницилиннің тұрақты сұрыпта организмдерін қосу немесе моноклондық антиденелерді шығару барысындағы гибридтық жасушаларды қосу.
12.2.3. Индикаторлық орта
Бұлар индикаторлық ортаны құрайды. Өз мақсатында олар бір организмді келесі бір организмнен айыру үшін қолданылады. Мысалы, кейбір бактериялар лактозаны қышқыл қылып бұзады. Агар маккони ортасы лактозаны бүлдірмеу үшін ашық – қызыл ферменттер бөледі. Бұл ішімдік суды тазалау жолында пайдалынылады. Суда лактозаны бұзатын ішек бактериялары болады.
12.2.4. Дайын орта
Агарды құрайтын құрғақ орта сатылымда кездеседі. Оларды суда 15 минут ұстайды, сосын тазалау үшін колбаларға құйып 121 °С 15 мин ұстайды. Автоклариволдық араласу барысында еріп кетеді. Автоклавир ішінде су қайнайды. Автоклав қорғаныш қақпағымен жабылады. Қысым көтерілген сайын судың қайнауы да көтеріледі. Осылайша барлық бактерияларды өлтіру үшін тазалайды. Кәдімгі жағдайда барлық бактерияны өлтіру үшін 103кП (кН/м3) қысымда 15 – 20мин ұстайды. Ал ішкі қысым 121 ° жетеді.
Сіңірімді сіңу үшін қажетті ортадан басқа бактерияларды өсіру үшін қоршаған ортаның қажетті жағдайын туғызу керек. Микроорганизм белгілі бір температурада жақсы өседі. Дәл сол жағдайда оларға мөлшерлі көміртегімен жарығы түсуі керек. Әртүрлі микроорганизмдерге әртүрлі қоршаған орта керек.
Температура
Температуралық жағдайдың әртүрлі организмдерге керектігі әртүрлі температураның ферменттерге тигізетін әсерін оңай қарап отыруға болады . Барлық жасушалар ферменттерден тұрады, ферментердің жасушаларының жұмыс жылдамдығы жасушалар өсіру жылдамдығы температураға байланысты. Әрбір микроорганизмге қажетті әртүрлі диапозондағы температура қажет.
Ең жоғарғы температура денатурациядан басталады. Денатурация ақуыздардың өмірлік жолын бастауының бақылауы. Оптималдық температураның өзгеру 0-5 °С аралығында болады, кейбір бактериялар үшін мұхиттарда 70°С дейін керек болады. Кейбір терең су астындағы бактрериялар вулкандық жағдайда өсетін бактериялар 118°С тірі болады.
Температураға қарай 4 топқа бөлуге болады.
Термофилдер, бұлар 45°С та өседі, мысалы, лактобацилустың кейбр түрлері(сүтте кездеседі).
Мезофилдер, өсу үшін қажетті температура 25°С-45°С. Сүтқоректілердің есебінен өмір сүретін бактериялар үшін қолайлысы 37°С, ал адам ішектерінде мекендеуші, мысалы Escherichiacoli(Е. coli)үшін максимум 42—43 °С қажет. Сонымен қатар, ашытқылар немесе саңырауқұлақтар да мезофилдерге жатады.
Психрофильдер, температура 15°С тан төмен, және олар 20°Стан жоғары температурада өспейді.
Бұл микроорганизмдерді тоңазытқышта сақтау өте қиын. Әсіресе, осы микроорганизмдер тамақтардың бұзылуына әкеліп соқтырады.
Психротрофтар, температуралық ортасы 15°С, бірақ олар одан төмен температурада өседі, мысалы, 5°С немесе одан да төмен. Candida түрінен келетін кейбір ашытқылар, кейбір көгерген саңырауқұлақтар, міне дәл осылар тоңазытқышты сақталатын тамақтарды бүлдіреді.
рН
Температура жағдайындағыдай әрбір түрі өзінің төменгі, жоғарғы және дәл температурасында өседі. Барлық бактериялар өзінің орта температурасында болады. Қышқыл ортада жақсы өсетін бактериялар микроорганизмдер ацидифиломдар деп аталады. Ал ылғал, дымқыл ауада бактериялар алколофилдер деп аталалды. Көптеген ашытқылар 4,5-5,0 рҺ орта ауасында өседі. Ал көктейтін саңырауқұлақтарға аз қышқыл орта керек.
Ауаның концентрациясы
Ауаның құрамынан жұту құрамына қарай микроорганизмдер келесі топқа бөлінеді:
*Облигатты аэробы. Мұнда микроорганизмдер тек көміртегімен өседі бұған туберкулездің бактериялары жатады, балдырлар мен қарапайымдылар тобындағы микроорганимдер жатады.
*Факультативтық аэробтар. Мұндай топтарға көміртекпен өсетін микроорганизм көміртегі жоө кезде анаэробты өсуін айтады. Оған бактериялар, ашытқылар жатады.
* Қамту анаэробы. Бұған ішек жолындағы қыртыстарда өсетін микроорганизмдер жатады. Оған сіреспе, ботулизм ауруын туғызуы мүмкін. Сонымен бірге уланып ауруы мүмкін.
*Микроаэрофилдер. Бұған өте төмен температурада өсетін микроорганизмдер жатады. Мысаы, сүтте кездесетін лактобоцилоз
4 бөлік түтіктің әр бөлімінде
*аэробтар
*анаэробтар
*факультативті аэробтар
*микроаэрофилдер
Иондық және осматикалық теңдік
Кейде жасушалардың ішкі құрамын концентрацияда ол жаймен әрі қарай осмос жолымен суын жоғалтады. Соған қарамастан, голофил деген организмдер өте жоғары концентрациялық тұздарда мысалы, үлкен Тұзды көл Америкада, Өлі көл Израилда кездеседі. Және оған қарама – қарсы суды кетіре алмайтын сулы ортада өсетін жасушаларда жоқ микроорганизмдде кездеседі. Соған қарамастан, олар дұрыс тепе – теңдік құрайды.
Дақылды орталары
Орта қатты және сұйық субстанцияны айтады. Дарақ деп осы микроб жасушаларының жиынтығының орта ішіндегі немесе орта сыртындағы өсуін айтады.
12.2.1. Қатты және сұйық орталар
Микроорганизмдерді қатты ортада немесе сұйық ортада өсіреді.
Қатты орта
Қатты орта өсіп тұрған өсімдіктер мен саңырауқұлақтар үшін өте ыңғайлы. Оларды гель образдағы 1-2 % концентрациясы бар сұйық қоректік ерітіндімен араластыру арқылы дайындайды. Сөйтіп, қоректік агарды алады. Агар 90—100 С температурада балқиды, ал 44°С қатады. Микроорганизмдерді беріктендіру үшін агар қосады.
Агар ашық түсті. Кейін полисахаридте айналған соң микроорганизм бұщылмау үшін төзімді болып табылады. Қатты және сұйық түріндегі мысалдар төмендегі бөлімдерде көрсетіледі.
Сұйық орта
Сұйық ортаны көбіне жарық санының өсуін зерттеу үшін қолданылады. жасушаларды қақпағы мықты жабылған пробиркаға салады. Қолдану үшін жасушалар алдын – ала спиральдинген болуы керек. Азғантай жасушалардың қосылуын бұл орталарда өсім дейді. Одан кейін өсіндерді термостат арқылы ыңғайлы ортада кездейсоқ жағдайлас отмргедилл береді. Ортаны көп шамада қолданған кезде тұнба қалмас үшін магнит арқылы араластырады. Ауа тазалануы үшін ауа тазалағыш түтіктерді тазалап тұрады. Ауа шашыратқыш құрал арқылы көптеген кішкентай түтіктерден еркін өтеді.
Ортаны көп шамада қолданған кезде тұнба қалмас үшін магнит арқылы отырады. Одан басқа оттектің норма жүйесіндегі концентрациясын сақтау үшін, орта арқылы тап - таза ауа кіреді. Б ұ л үшін әртүрлі құралдар , яғни сүзбе және әйнекті сүзбе , немесе сіңірілген мақталы вата қолданылады. Ауа тозаңдатқыш , я ғни көптеген кішкетай тесіктер арқылы өтеді. Олар мінсіз көпіршіктер алуға мүмкіндік береді. Оны дақыл ыдысына бара жатқан кекзінен, түтіктің соңына қарай бекітеді.
12.2.2. Байытылған селективтік орта
Байытылған орта бұл белгілі бір микроорганизмді қажетті қорлармен толтырырылуы. Сол себепті басқа микроорганизмдерге қарағанда, бұлардың өсімі жоғары болады. Ортаны түрлі дақылмен толтыру барысында табиғатта керекті микроорганизмдердің неше түрлерін шығаруға болады. Және олар үшін рН, оттек көзі, қуат көзі, температура барлығы жоғары нәтижеде қаралу керек. Мысалы, ТИФ ауруын туғызатын Salmonella фехалий ортасымен араласа отырып, селендік микроорганизмді құрайды. Фехалийде кәдімгі ішек бактериялары да болады, олар селенмен жойылады.
Селективті орта – бірен саран, ортадан басқа ортаның көбінің өсуін тежейді. Бұған мысал келтіретін болсақ, дақылға пеницилиннің тұрақты сұрыпта организмдерін қосу немесе моноклондық антиденелерді шығару барысындағы гибридтық жасушаларды қосу.
12.2.3. Индикаторлық орта
Бұлар индикаторлық ортаны құрайды. Өз мақсатында олар бір организмді келесі бір организмнен айыру үшін қолданылады. Мысалы, кейбір бактериялар лактозаны қышқыл қылып бұзады. Агар маккони ортасы лактозаны бүлдірмеу үшін ашық – қызыл ферменттер бөледі. Бұл ішімдік суды тазалау жолында пайдалынылады. Суда лактозаны бұзатын ішек бактериялары болады.
12.2.4. Дайын орта
Агарды құрайтын құрғақ орта сатылымда кездеседі. Оларды суда 15 минут ұстайды, сосын тазалау үшін колбаларға құйып 121 °С 15 мин ұстайды. Автоклариволдық араласу барысында еріп кетеді. Автоклавир ішінде су қайнайды. Автоклав қорғаныш қақпағымен жабылады. Қысым көтерілген сайын судың қайнауы да көтеріледі. Осылайша барлық бактерияларды өлтіру үшін тазалайды. Кәдімгі жағдайда барлық бактерияны өлтіру үшін 103кП (кН/м3) қысымда 15 – 20мин ұстайды. Ал ішкі қысым 121 ° жетеді.

2. Асептикалық шаралар
Бұл шаралар стирилденген құралдарды ластанудан сақтау үшін қолданылады. Бактериялар мен саңырауқұлақ споралары қоршаған ортада кеңінен пайдалынылады. Сонымен бірге лабаротрияларда да. Микробиология ылғи қоршаған ортаның тазалығы үшін жұмыстанады. Мектеп және колледж лабароторияларында сақтану әдістері үшін қолданады. Дұрыс жұмыс жүргізілуі үшін арнайы құралдармен жабдықталған зертхана қажет.
12.3.1. Ыдыс толтыру.
Бұл ең негізгі микробиологиялық әдістеме. Қоректі затты құрайтын агарды Петри – ыдысы дейді. Петри – ыдысы арнайы терең емес дөңгелек контейнерден жасалған. Оларды диаметрі 9 см. Олар бактериялар мен саңырау құлақтың немесе дақыл тіндерінің қатты ортада бейімделуіне қолайлы. Пластикалық чашкалар қолданыстан кейін лақтырылады. Оларды әдетте автоклавтайды, себебі дақылды жою үшін. Бұл үшін оларды малтады. Тостағанды тек қана стерильдік гамма емдеу жолымен мөр басылған қораптардан ғана сатып алады. Қақпақтар тостақанның ластануынан сақтайды. Бірақ, газ молекулалары тостақанның ішкі көлемімен араласады және қоршаған ортасы микроскопиялық тегіс емес орын қақпақпен жанасады. Сондықтан дақылға оттек қолжетімді болады, ал қостотықты көміртек сыртқа шығарылады.
Бұл шаралар стирилденген құралдарды ластанудан сақтау үшін қолданылады. Бактериялар мен саңырауқұлақ споралары қоршаған ортада кеңінен пайдалынылады. Сонымен бірге лабаротрияларда да. Микробиология ылғи қоршаған ортаның тазалығы үшін жұмыстанады. Мектеп және колледж лабароторияларында сақтану әдістері үшін қолданады. Дұрыс жұмыс жүргізілуі үшін арнайы құралдармен жабдықталған зертхана қажет.
12.3.1. Ыдыс толтыру.
Бұл ең негізгі микробиологиялық әдістеме. Қоректі затты құрайтын агарды Петри – ыдысы дейді. Петри – ыдысы арнайы терең емес дөңгелек контейнерден жасалған. Оларды диаметрі 9 см. Олар бактериялар мен саңырау құлақтың немесе дақыл тіндерінің қатты ортада бейімделуіне қолайлы. Пластикалық чашкалар қолданыстан кейін лақтырылады. Оларды әдетте автоклавтайды, себебі дақылды жою үшін. Бұл үшін оларды малтады. Тостағанды тек қана стерильдік гамма емдеу жолымен мөр басылған қораптардан ғана сатып алады. Қақпақтар тостақанның ластануынан сақтайды. Бірақ, газ молекулалары тостақанның ішкі көлемімен араласады және қоршаған ортасы микроскопиялық тегіс емес орын қақпақпен жанасады. Сондықтан дақылға оттек қолжетімді болады, ал қостотықты көміртек сыртқа шығарылады.
|
|
Қоректік агардық балқыту тәртібі 12.3. суретте көрсетілген.
3. Егу әдісі
Микроорганизмді қоректену ортасына енгізу үшін асептикалық әдістемелерді қолдану қажет. Бұл рәсімдер көлемі бойынша қолданылады.
12.4.1. Қатты ортаны егу
Штрих пен егу немесе өсімді көбейту әдістері жасау
Бұл әдіс 12.4 суретте көрсетілген. Ол бактериялардан таза бактерия бөліп шығару. Егу үшін бұл жерде сымды ілгіштерді пайдаланады. Ал ол үшін алдымен, 12.4,А суреттегідей, тазарту үшін қыздыру керек. Содан кейін ілмек арқылы жұқа пленка микроорганизмін алады. Ілмектерді жаймен орта үстімен штрих жасай отырып, жүргізеді. Әрбір штрих сайын құтыны сәл қозғайды.(инкубация біткенге дейінгі уақыттағы соңғы штрихтауды көреміз деп, тіпті үміттенбей – ақ қойыңдар!). әдіс жасалған кезде штрихтар тез –тез істелініп кетілінір кетуі мүмкін. Бұл әдісті үйренген соң тез жасауға болады. Осы әдіс бойынша бактерияларды қорек ортасынан шығаруға болады. Мысалы, сүттен, судан. Әдістеме жүргізбес бұрын қауіпсіздік нұсқаулармен танысу керек.
Агар бетін егу
Бұл әдіс 12.5.суретінде көрсетілген. Бұл микроорганизмді сұйық суспензиядан қатты ортаға аудару үшін ыңғайлы. Жасушалардың өмірінің көптігін анықтау үшін керек.12.6.1.суреттен қарайсыңдар. Бұл әдісті түгел газондау жұмысында қолдануға болады.Сонымен қатар , бұл әдіс ингибиттердің жылдамдығы үшін антибиотиктер мен дизенфекциялар әдістерінде пайдаланылады. Ингибитор агар арқылы дифуздалады. Сен бірге,ол жан-жағының өсу зонасының аумағын көрсетеді. Зона сыйлығының диометрі ингиборлық бағасын білуге болады.
Толтыру егісі
Бұл әдіс,агар бетіне егу әдісінің жасушаның өмірінің ұзақтығын санау үшін және сұйық мәдениетін пайдалану болып табылады. Жасушалар барлық ортада бөлінетіндіктен оларды көбірек санауға болады. Жобамен 1000 қатар ыдыста. Бірақ өскен қатардың саны аздау.
3. Егу әдісі
Микроорганизмді қоректену ортасына енгізу үшін асептикалық әдістемелерді қолдану қажет. Бұл рәсімдер көлемі бойынша қолданылады.
12.4.1. Қатты ортаны егу
Штрих пен егу немесе өсімді көбейту әдістері жасау
Бұл әдіс 12.4 суретте көрсетілген. Ол бактериялардан таза бактерия бөліп шығару. Егу үшін бұл жерде сымды ілгіштерді пайдаланады. Ал ол үшін алдымен, 12.4,А суреттегідей, тазарту үшін қыздыру керек. Содан кейін ілмек арқылы жұқа пленка микроорганизмін алады. Ілмектерді жаймен орта үстімен штрих жасай отырып, жүргізеді. Әрбір штрих сайын құтыны сәл қозғайды.(инкубация біткенге дейінгі уақыттағы соңғы штрихтауды көреміз деп, тіпті үміттенбей – ақ қойыңдар!). әдіс жасалған кезде штрихтар тез –тез істелініп кетілінір кетуі мүмкін. Бұл әдісті үйренген соң тез жасауға болады. Осы әдіс бойынша бактерияларды қорек ортасынан шығаруға болады. Мысалы, сүттен, судан. Әдістеме жүргізбес бұрын қауіпсіздік нұсқаулармен танысу керек.
Агар бетін егу
Бұл әдіс 12.5.суретінде көрсетілген. Бұл микроорганизмді сұйық суспензиядан қатты ортаға аудару үшін ыңғайлы. Жасушалардың өмірінің көптігін анықтау үшін керек.12.6.1.суреттен қарайсыңдар. Бұл әдісті түгел газондау жұмысында қолдануға болады.Сонымен қатар , бұл әдіс ингибиттердің жылдамдығы үшін антибиотиктер мен дизенфекциялар әдістерінде пайдаланылады. Ингибитор агар арқылы дифуздалады. Сен бірге,ол жан-жағының өсу зонасының аумағын көрсетеді. Зона сыйлығының диометрі ингиборлық бағасын білуге болады.
Толтыру егісі
Бұл әдіс,агар бетіне егу әдісінің жасушаның өмірінің ұзақтығын санау үшін және сұйық мәдениетін пайдалану болып табылады. Жасушалар барлық ортада бөлінетіндіктен оларды көбірек санауға болады. Жобамен 1000 қатар ыдыста. Бірақ өскен қатардың саны аздау.
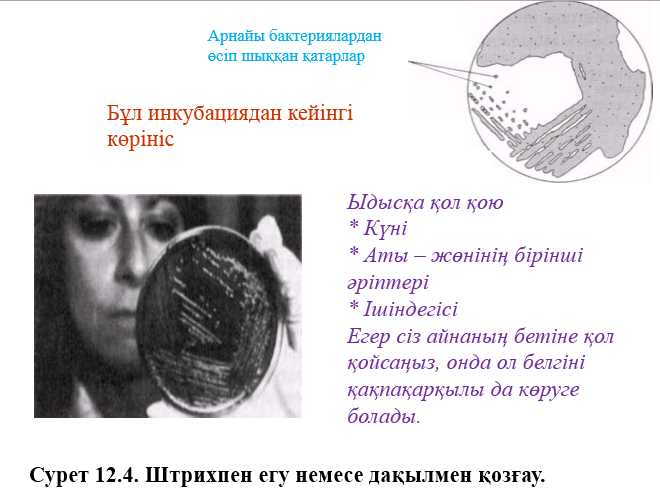
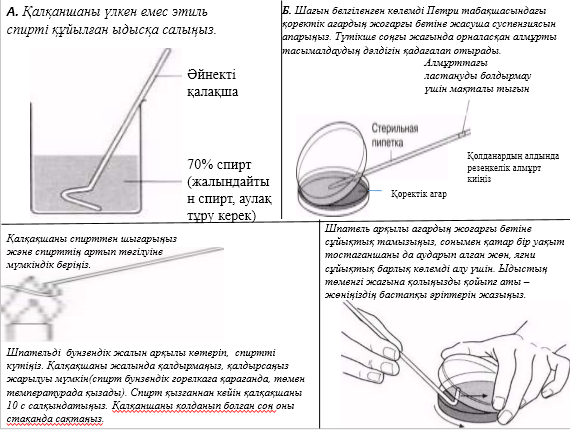
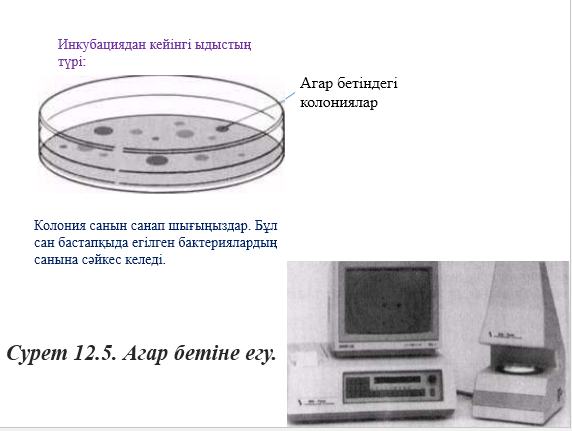
ЕГУ
Бұл әдіс анаэробтық организмнің төменгі оттегі концентрациясында өсу жолын қолданылады. Әдетте, қоректік агарлық ортада қолданады. Агардың тереңдігінің арқасында, ыдысқа қарағанда оттегінің құрамы азаяды. Егілу тіке ине арқылы ілгішпен жүргізіледі. Дақылдың азғантай мөлшері қатты және сұйық жағдайда иненің ұшымен алынып, агарды вертикальды теседі. 12.7. суретінде көрсетілген. Дақыл агардың барлық жағынан өседі.
Бұл әдіс анаэробтық организмнің төменгі оттегі концентрациясында өсу жолын қолданылады. Әдетте, қоректік агарлық ортада қолданады. Агардың тереңдігінің арқасында, ыдысқа қарағанда оттегінің құрамы азаяды. Егілу тіке ине арқылы ілгішпен жүргізіледі. Дақылдың азғантай мөлшері қатты және сұйық жағдайда иненің ұшымен алынып, агарды вертикальды теседі. 12.7. суретінде көрсетілген. Дақыл агардың барлық жағынан өседі.

12.5. Бактериялардың дамуы
12.5.1. Өсу популяциясы
Бактериялардың өсу популлияциясы белгілі бір өлшемге жеткенде, олар жыныссыз көбейеді. Оны қарапайым бөліну дейді. Сонымен бірге, жасушалық екі гендік жасушаға бөлінеді. Егер жалғыз бактерияны қоршаған ортаға орнатсақ, бактерия әрбір 30мин сайын көбейеді. Кесте 12.2.
12.5. егер сіз 2 бөлімдегі 2.1. сұраққа жауап бере алмасаңыз 12.2. кестесінде бактериялардың саны рет – ретімен тізіліп тұрғанына қарай берілген графикті қолданыңыз (график А) және lg уақытына қарай бактерия саны (график Б). Мұндай пішіндегі бактериялар туралы не айтуға болады?
Клеткалардың таблица 12.2. суретінде көрсеткендей лагоретті немесе экспонентті өсуі белгілі жағдай.
12.2 суретіне қарап оны оңай түсіндіруге болады. Көрсеткіштердің лагарифимдік немесе экспоненттік көрсеткітерді экспоненттік деп атайды.
Олардың қатары 0,1,2,3.... сол сияқты өсу қатарында болады.
12.2 А таблицасында лагорофимді 2 Г және В жолдарымен салыстырыңдар. А графасындағы қисық сызық лагорофимдік және экспоненттік қисық деп аталады. Мұндай сызықтар тіке және қисықтар график сыза отырып, жартылай графикті масштабта болады.
Бактериялар жақсы жағдайда өсуі экспонциалды болады.
Бактериялардың екі есе өсуі қараластырады. Бақылап қарасақ, екі қисық сызықтың біреуі өлгендерімен қоса бактериялардың санын көрсетеді. Екі сызық бактериялардың өмір сүргіштігін көрсетеді. Бұл қисық сызық 4 фазалы:
Лагфаза. Бактериялар жаңа ортаға бейімделеді. Үйренгенде аса қатты өсуге жетпейді.
Лагарифиялық. Мұнда бактериялар аса жылдамдықпен өсіп, бактерия саны өсіп, экспонциалды тіке жол жасайды. Бұл фазада 2ге бөліну қалыпты жағдай болып саналады. Бара -бара қатарлардың өсуі жайлайды. Екіге бөлінуі көбейеді.
Дақыл стационарлық фазаға өтеді. Жаңа жасушалардың өсуі жайланады. Содан соң тоқталады. Кез келген жасушалардың өсуі басқа жасушалардың өлуіне әкеледі. Сондықтан саны жағынан тірі жасушалар бұрынғы қалпында болады. Мұндай қалыпқа енуі қоректік зат азаюынан, улы заттардың көбеюуінен,оттегінің жетіспеуінен болады. Бактериялардың өсуі рҺ өзгерісінің пайдалануынан да болады.
Соңғы өсу фазасының жайлануынан жасушаның өлу жылдамдығы өседі. Бара – бара жасушалар көбеюін тоқтатады. Бактериялардың санының өсуі келесі кестеде көрсетілген 12.8
12.5.1. Өсу популяциясы
Бактериялардың өсу популлияциясы белгілі бір өлшемге жеткенде, олар жыныссыз көбейеді. Оны қарапайым бөліну дейді. Сонымен бірге, жасушалық екі гендік жасушаға бөлінеді. Егер жалғыз бактерияны қоршаған ортаға орнатсақ, бактерия әрбір 30мин сайын көбейеді. Кесте 12.2.
12.5. егер сіз 2 бөлімдегі 2.1. сұраққа жауап бере алмасаңыз 12.2. кестесінде бактериялардың саны рет – ретімен тізіліп тұрғанына қарай берілген графикті қолданыңыз (график А) және lg уақытына қарай бактерия саны (график Б). Мұндай пішіндегі бактериялар туралы не айтуға болады?
Клеткалардың таблица 12.2. суретінде көрсеткендей лагоретті немесе экспонентті өсуі белгілі жағдай.
12.2 суретіне қарап оны оңай түсіндіруге болады. Көрсеткіштердің лагарифимдік немесе экспоненттік көрсеткітерді экспоненттік деп атайды.
Олардың қатары 0,1,2,3.... сол сияқты өсу қатарында болады.
12.2 А таблицасында лагорофимді 2 Г және В жолдарымен салыстырыңдар. А графасындағы қисық сызық лагорофимдік және экспоненттік қисық деп аталады. Мұндай сызықтар тіке және қисықтар график сыза отырып, жартылай графикті масштабта болады.
Бактериялар жақсы жағдайда өсуі экспонциалды болады.
Бактериялардың екі есе өсуі қараластырады. Бақылап қарасақ, екі қисық сызықтың біреуі өлгендерімен қоса бактериялардың санын көрсетеді. Екі сызық бактериялардың өмір сүргіштігін көрсетеді. Бұл қисық сызық 4 фазалы:
Лагфаза. Бактериялар жаңа ортаға бейімделеді. Үйренгенде аса қатты өсуге жетпейді.
Лагарифиялық. Мұнда бактериялар аса жылдамдықпен өсіп, бактерия саны өсіп, экспонциалды тіке жол жасайды. Бұл фазада 2ге бөліну қалыпты жағдай болып саналады. Бара -бара қатарлардың өсуі жайлайды. Екіге бөлінуі көбейеді.
Дақыл стационарлық фазаға өтеді. Жаңа жасушалардың өсуі жайланады. Содан соң тоқталады. Кез келген жасушалардың өсуі басқа жасушалардың өлуіне әкеледі. Сондықтан саны жағынан тірі жасушалар бұрынғы қалпында болады. Мұндай қалыпқа енуі қоректік зат азаюынан, улы заттардың көбеюуінен,оттегінің жетіспеуінен болады. Бактериялардың өсуі рҺ өзгерісінің пайдалануынан да болады.
Соңғы өсу фазасының жайлануынан жасушаның өлу жылдамдығы өседі. Бара – бара жасушалар көбеюін тоқтатады. Бактериялардың санының өсуі келесі кестеде көрсетілген 12.8
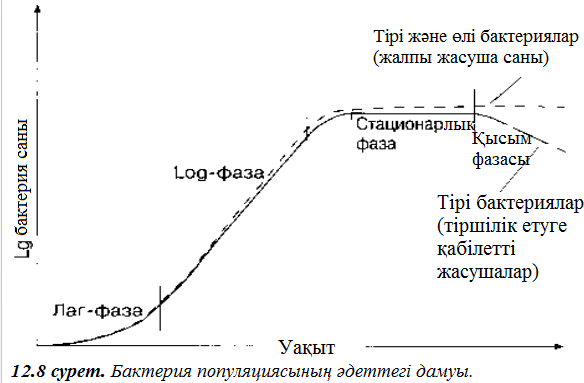
12.6.1. Өмір сүргіш жасушалардың саны.
Кейде өмір сүргіш жасушалардың санын білген маңызды. Мысалы, сүтті постеризация жасағанда тірі қалған бактериялардың санын білу және өлген жасушалардың санын білуімізге болады. Шикізат өндіруде тек тірі жасушалар жұмыс жасайды. Сондықтан қаншасы өлі, қаншасы тірі екенін білу маңызды.
Тірі жасушалардың санын ыдыстан санауға болады. Ал өмір сүргіш жасушаларды гемоцитометрин әдісімен табуға болады.
12.5.2. Диауксия
Диауксия дегеніміз – бір немесе бірнеше өткізгіш өсу дақылының көбеюі. Бұл жағдай бактериялардың бір немесе екі альтернативтық қоректену көзі болған жағдайда болады.
12.5.3. Бірінші және екінші метаболиттің пайда болуы
Бірінші метаболиттер, бұл метоболизм өнімі. Олар өсумен өмір сүруге қажет.
Екінші метаболиттер метаболизм өнімі. Бұлардың өсуге де, көбеюге де қатысы жоқ. Бірақ олар кейбір микроболиттерден қорғану үшін және көбейтуін тоқтату үшін өте қажетті. Бұлардың кейбіреулері тірі организмдерге улы, сондықтан оны химиялық қару ретінде қолданады.
Екінші метаболиттердің ішінде керекті, маңызды антибиотиктер алынады.
12.6. Дақыл саласындағы бактериялар мен саңырауқұлақтарды өлшеу.
Өткен бөлімде біз бактериялардың қисық өсу жолын талқыладық. Бактериялардың өсуін бақылаған кезде біз қанша жасуша барын анықтай алдық. Олардың өзгеріс параметрлерін тексердік және лайлығын, я болмаса, газының бөлінуін көрдік.
Әдетте, азғантай мөлшердегі микроорганизмдерді стирильденген ортада өседі немесе оптималдық температурада, инкубаторда өсіреді. Олардың өсімін иннапуляция басталғаннан өлшейді. Кейбір ғылыми тексерулер бірнеше рет эксперимент жүзінде қайталанылады. Бақылау жүйелерінен өтеді.
Ал кейбір әдіс түрлері белгілі бір маман иелерінің қолдарында жасалады. Олар ондай дәл болмайды. Тексеруді екі жүйеде бірдей өткізіп, әрқайсысын қатаң бақылауда тексеру қажет. Сол кезде өсу ортасына ешқандай микроорганизмдерді қоспай жасап, шынымен стирилді жұмыс жасап жатқанын тексеру қажет. Жасушалардың өсу жолдарын 2 жолмен табуға болады. Ал соның ішінде өмір сүргіш жасушалардың (бұл жерде тек қана тірі жасушаларды санайды). Сол жерде барлық жасушалардың санын анықтау қажет.
12.6.1. Өмір сүргіш жасушалардың саны.
Кейде өмір сүргіш жасушалардың санын білген маңызды. Мысалы, сүтті постеризация жасағанда тірі қалған бактериялардың санын білу және өлген жасушалардың санын білуімізге болады. Шикізат өндіруде тек тірі жасушалар жұмыс жасайды. Сондықтан қаншасы өлі, қаншасы тірі екенін білу маңызды.
Тірі жасушалардың санын ыдыстан санауға болады. Ал өмір сүргіш жасушаларды гемоцитометрин әдісімен табуға болады.
Агар бетінде егу
Бұл әдіс 12.4.1 суретінде берілді. Азғантай белгілі көлемдегі дақылды сіңімді агар бетіне жағады. Бұл әдістің келіспейтін жері дақылдың жартысы шпательдің жартысының бетінде қалып қояды. Сондықтан жасушалардың дәл санын білу мүмкін емес. Бірақ көбіне бұл аза маңызды емес.
Бұл әдіс әрбір бактерия белгілі бір уақыттан кейін, мысалы, екі күннен кейін (калония) құрайды. Сөйтіп, бактериялар саны дәл сол санда баған құрады. Жай көзбен бізге 100000 үстінде жасушалар көрінеді. Ал негізінде бағанда бірнеше млн бактерияларды құрайды.
Кейде өмір сүргіш жасушалардың санын білген маңызды. Мысалы, сүтті постеризация жасағанда тірі қалған бактериялардың санын білу және өлген жасушалардың санын білуімізге болады. Шикізат өндіруде тек тірі жасушалар жұмыс жасайды. Сондықтан қаншасы өлі, қаншасы тірі екенін білу маңызды.
Тірі жасушалардың санын ыдыстан санауға болады. Ал өмір сүргіш жасушаларды гемоцитометрин әдісімен табуға болады.
12.5.2. Диауксия
Диауксия дегеніміз – бір немесе бірнеше өткізгіш өсу дақылының көбеюі. Бұл жағдай бактериялардың бір немесе екі альтернативтық қоректену көзі болған жағдайда болады.
12.5.3. Бірінші және екінші метаболиттің пайда болуы
Бірінші метаболиттер, бұл метоболизм өнімі. Олар өсумен өмір сүруге қажет.
Екінші метаболиттер метаболизм өнімі. Бұлардың өсуге де, көбеюге де қатысы жоқ. Бірақ олар кейбір микроболиттерден қорғану үшін және көбейтуін тоқтату үшін өте қажетті. Бұлардың кейбіреулері тірі организмдерге улы, сондықтан оны химиялық қару ретінде қолданады.
Екінші метаболиттердің ішінде керекті, маңызды антибиотиктер алынады.
12.6. Дақыл саласындағы бактериялар мен саңырауқұлақтарды өлшеу.
Өткен бөлімде біз бактериялардың қисық өсу жолын талқыладық. Бактериялардың өсуін бақылаған кезде біз қанша жасуша барын анықтай алдық. Олардың өзгеріс параметрлерін тексердік және лайлығын, я болмаса, газының бөлінуін көрдік.
Әдетте, азғантай мөлшердегі микроорганизмдерді стирильденген ортада өседі немесе оптималдық температурада, инкубаторда өсіреді. Олардың өсімін иннапуляция басталғаннан өлшейді. Кейбір ғылыми тексерулер бірнеше рет эксперимент жүзінде қайталанылады. Бақылау жүйелерінен өтеді.
Ал кейбір әдіс түрлері белгілі бір маман иелерінің қолдарында жасалады. Олар ондай дәл болмайды. Тексеруді екі жүйеде бірдей өткізіп, әрқайсысын қатаң бақылауда тексеру қажет. Сол кезде өсу ортасына ешқандай микроорганизмдерді қоспай жасап, шынымен стирилді жұмыс жасап жатқанын тексеру қажет. Жасушалардың өсу жолдарын 2 жолмен табуға болады. Ал соның ішінде өмір сүргіш жасушалардың (бұл жерде тек қана тірі жасушаларды санайды). Сол жерде барлық жасушалардың санын анықтау қажет.
12.6.1. Өмір сүргіш жасушалардың саны.
Кейде өмір сүргіш жасушалардың санын білген маңызды. Мысалы, сүтті постеризация жасағанда тірі қалған бактериялардың санын білу және өлген жасушалардың санын білуімізге болады. Шикізат өндіруде тек тірі жасушалар жұмыс жасайды. Сондықтан қаншасы өлі, қаншасы тірі екенін білу маңызды.
Тірі жасушалардың санын ыдыстан санауға болады. Ал өмір сүргіш жасушаларды гемоцитометрин әдісімен табуға болады.
Агар бетінде егу
Бұл әдіс 12.4.1 суретінде берілді. Азғантай белгілі көлемдегі дақылды сіңімді агар бетіне жағады. Бұл әдістің келіспейтін жері дақылдың жартысы шпательдің жартысының бетінде қалып қояды. Сондықтан жасушалардың дәл санын білу мүмкін емес. Бірақ көбіне бұл аза маңызды емес.
Бұл әдіс әрбір бактерия белгілі бір уақыттан кейін, мысалы, екі күннен кейін (калония) құрайды. Сөйтіп, бактериялар саны дәл сол санда баған құрады. Жай көзбен бізге 100000 үстінде жасушалар көрінеді. Ал негізінде бағанда бірнеше млн бактерияларды құрайды.
Егер ыдыста онша көп те емес, аз да емес бактерияларды Петридің қақпағын ашпастан оңай санауға болады. Осы мысалды дәлдік үшін қайалауға болады. 12.9. сурет бойынша.
12.3. бөліміндегі 12.9 бөліміндегі бактериялар саны сүтті тексерудегі әдіспен тексерілді. Кейбір болмайтын заттар шектеулі бөлімінің соңында талқыланады.
Ашытқылардың ашытқыш жасушаларының көбеюуін гемецотометрмен және көк метиленмен санауға болады. Бұл әдіс келесі бөлімде жазылған, тірі жасушаларды санау жүзіндегі қиындықтар мұндай санау кезінде көптеген ауыртпашылықтар әкеледі.
Ашытқылар
Ашытқылардың ашытқыш жасушаларының көбеюуін гемецотометрмен және көк метиленмен санауға болады. Бұл әдіс келесі бөлімде жазылған, тірі жасушаларды санау жүзіндегі қиындықтар мұндай санау кезінде көптеген ауыртпашылықтар әкеледі.
Жасушалардың өмір сүру бейімділігінде туындайтын проблемалар:
12.6.2. Жалпы жасуша сандары
Гэмоцитометр
Жалпы жасуша сандары бактериялармен ашытқыш жасушардың сұйық дақылдардың мысалы, сорпада микроскоп арқылы тіке санауға болады. Бұл әдіс ірі ашытқыш жасушаларды санауға ыңғайлы. Санаған кезде міндетті түрде иммерцион майы болу керек.
5.11.2. суреттерінде қараңыз.
Негізгі, кәдімгі гемоцитометрмен маңызды терезешелерді қолданады.
12.10 сурет қараңыз. Бұл әдіс гемоциото-метрия әдісімен атақты. Екі терезенің арасындағы дақылды терезешені сындырып алмай, қысу керек. Содан соң терезені жайлап қозғап, одан шыққан түрлі түсті сызықшаны қараймыз. 12.10 суретін қааймыз.
Тексеру ылғалдылығы пипеткамен терезеге тамызылады. Осыдан каппилярлы шығынды анықтауға болады. Камераға 2 – 3 мин тастаймыз. Сол кезде жасушалар санауға ыңғайлы болып жатады. Содан соң микроскоп тордан барлық жасушаны санайды. 600 жасушадан кем санамау керек. Егер жасушалар шетке кетсе, мысалы (жоғары немесе төмен)кетсе, ол жасушалардың жартысын санау қажет емес. Әрбір шаршы, өзінің белгілі ауданын, соған кететін сұйықтығы ,ылғи бірдей есептеледі(көбіне 0,02 немесе 0,1мл). Егер шаршыдағы жасуша 30дан асса, ылғалды араластыру керек. Әрбір кішкентай шаршының өзінің көлемі болады.
0,05 мм * 0.05мм=0,0025мм2.
Жалпы ауданның біршаршысына бір мм2 егер терезешелердің арасындағы ашықтық 0.02 мм құрса онда жалпы ауданы
1*0,02мм=0,02мм3 болады. Осылай жалпы аудандағы жасушалардың санын санауға болады. Естеріңде болса, 1000 мм3 = 1 см3қа, ал 1000см3=1дм3. Жасушаларды санаған кезде көк метиллин түсін аламыз. Бұл түске тек өлі жасушалар боялады. Тірі жасушалар түрлі түсті жасушаларды шығарып тастайды. Осылайша, тірі жасушаларды санауға болады.
12.3. бөліміндегі 12.9 бөліміндегі бактериялар саны сүтті тексерудегі әдіспен тексерілді. Кейбір болмайтын заттар шектеулі бөлімінің соңында талқыланады.
Ашытқылардың ашытқыш жасушаларының көбеюуін гемецотометрмен және көк метиленмен санауға болады. Бұл әдіс келесі бөлімде жазылған, тірі жасушаларды санау жүзіндегі қиындықтар мұндай санау кезінде көптеген ауыртпашылықтар әкеледі.
Ашытқылар
Ашытқылардың ашытқыш жасушаларының көбеюуін гемецотометрмен және көк метиленмен санауға болады. Бұл әдіс келесі бөлімде жазылған, тірі жасушаларды санау жүзіндегі қиындықтар мұндай санау кезінде көптеген ауыртпашылықтар әкеледі.
Жасушалардың өмір сүру бейімділігінде туындайтын проблемалар:
- Кейбір бактериялар жасуша топтарымен жасуша тізбектерін құрайды. Мысалы, стрептококтармен стафилококтер(Сурет 2.10.). Әрбір жасуша тобы бір бағанның басталуын көрсетеді. Сондықтан тірі жасушалардың есептеу бактериялардың саны емес, құрылған тізбегіне қарай есептейміз.
- Егер бактеряилардың бірнеше түрі қатысса, мысалы, су, топырақ, жер, сүт қатысса онда өсу жағдайы бірдей типте болмайды. Сондықтан кейбір бактериялар тез өседі. Ал тізбек бактериялары түсініксіз қабат құрайды.
12.6.2. Жалпы жасуша сандары
Гэмоцитометр
Жалпы жасуша сандары бактериялармен ашытқыш жасушардың сұйық дақылдардың мысалы, сорпада микроскоп арқылы тіке санауға болады. Бұл әдіс ірі ашытқыш жасушаларды санауға ыңғайлы. Санаған кезде міндетті түрде иммерцион майы болу керек.
5.11.2. суреттерінде қараңыз.
Негізгі, кәдімгі гемоцитометрмен маңызды терезешелерді қолданады.
12.10 сурет қараңыз. Бұл әдіс гемоциото-метрия әдісімен атақты. Екі терезенің арасындағы дақылды терезешені сындырып алмай, қысу керек. Содан соң терезені жайлап қозғап, одан шыққан түрлі түсті сызықшаны қараймыз. 12.10 суретін қааймыз.
Тексеру ылғалдылығы пипеткамен терезеге тамызылады. Осыдан каппилярлы шығынды анықтауға болады. Камераға 2 – 3 мин тастаймыз. Сол кезде жасушалар санауға ыңғайлы болып жатады. Содан соң микроскоп тордан барлық жасушаны санайды. 600 жасушадан кем санамау керек. Егер жасушалар шетке кетсе, мысалы (жоғары немесе төмен)кетсе, ол жасушалардың жартысын санау қажет емес. Әрбір шаршы, өзінің белгілі ауданын, соған кететін сұйықтығы ,ылғи бірдей есептеледі(көбіне 0,02 немесе 0,1мл). Егер шаршыдағы жасуша 30дан асса, ылғалды араластыру керек. Әрбір кішкентай шаршының өзінің көлемі болады.
0,05 мм * 0.05мм=0,0025мм2.
Жалпы ауданның біршаршысына бір мм2 егер терезешелердің арасындағы ашықтық 0.02 мм құрса онда жалпы ауданы
1*0,02мм=0,02мм3 болады. Осылай жалпы аудандағы жасушалардың санын санауға болады. Естеріңде болса, 1000 мм3 = 1 см3қа, ал 1000см3=1дм3. Жасушаларды санаған кезде көк метиллин түсін аламыз. Бұл түске тек өлі жасушалар боялады. Тірі жасушалар түрлі түсті жасушаларды шығарып тастайды. Осылайша, тірі жасушаларды санауға болады.
Лайлануды өлшеу
Бұл әдіс неффилометр әдісі негізінде қарапайым. Суспензияда қанша көп жасуша болса, оның лайлылығы сонша жоғары болады. Сол қарайған жерден жасушаларды оңай санауға болады. Негізгі сатылымда бар броуновтық түтікшелерде қолданған ыңғайлы. Олардың суспензияларында сульфат, барий әртүрлі қоюлықта сұйықтан (түтікше №1) лайланғанға дейін (түтікше №10) болады.
Түтікшелердің жинағына кесте қосылады. Бұл қоблықтардың майлылығын кең арнада, әр түрлі микроорганизмде өлшейді. Сонымен бірге лайдан жарықтың өткізу қалыңдығын көруге болады .
Бұл кезде калорийметр мен спектро фотометрді қолданады. Негігі қызыл түсті қолдан,ған жақсы. Себебі ол дақылдың ортасында сары түспен араласқанда шағылыспайлы. Лайды өлшегенде, өсіп жатқан жасушалар үйіліп, жиналып қалса онда тексеру кезінде қате кетуі мүмкін.
12.6.3. Сапасыз әдістер
Микроорнаизмдерің өсуінің өлшеуін басқа әдістермен де өлшеуге болады. Мысалы, саңырауқұлақтарды уақыт өтісуімен мецилиде диаметрінің өсуімен өлшеуге болады.
Бұл әдіс температураның саңырауқұлақтарға қандай өзгеріс беретінін тексереді.
Егер саңырауқұлақты сұйық ортада өсірсе, белгілі уақыттан кейін оның дақылын сүзіп алуға болады.
12.7. Боялған бактериялар – Грамм бойынша бояу
Фазалық микроскоптың көмегімен тірі бактерияларды боялған түрінде анықтап көруге болады. Бұл әдісті Грамм бойынша бояу деп аталады. Бұл әдіс 1884 жылы даттық дәріге Кристшен Граммның табуымен аталды. Боялғанға дейін бұл бактериялар түссіз, боялғаннан кейін Грамм кәдімгі бактериялары күлгін түске айналды. Ал Граммға қарама – қарсы бактериялар қызыл түске боялады. Осы екі типтегі бактериялардың айырмашылығы 2.5.1. суретінде көрсетілген.
Түске бояу әдістері 12.2., 12.9. бөлімдерінде көрсетілген.
12.8 Вирустарды дақылдау
Вирустарды өсіру, бактерияларды, дақылдарды өсіруге қарағанда қиындау болады. Себебі вирустар тірі организмдер ішінде өседі. Вирусты тексеру арқылы бір бүтін өсімдікті немесе бір бүтін денені бүлдіріп алуы мүмкін. Сондықтан дақылдың белгілі бір жасушасын немесе тінін алып зерттейді. 12.11 суретінде көрсетілген.
Ертеде тауықтың ішінде өсіп жатқан жұмыртқаның дақылын алып зерттеген. Бұған ұқсас вирустар шошқа мойын мен тұмау ауруларын болдырпмау үшін егетін вакцина дайындаған. Жасуша дақылы бұл жасуша суспензиясының сұйық ортада өсуі. Ал тін дақылы бұл өсімдік немесе тірі организмнен алынған сұйық немесе қатты ортаны айтады. Жасуша дақылы мерристеманы қолданады. Бұл дегеніміз жасушалардың белсенді бөлінетін жері, материалды бүлінген өсімдіктен алады немесе дақылды керекті вируспен ауыртады. Тірі дақылдың жасушаларынан жұқа жасуша дайындап, соны зерттейді. Полиовирус 1-ші маймылдың бөтекесінен алынды. Тірі организмнің дақылын қолдану үшін өкпе жасушаларын обыр ауруымен ауырған адамның амнинонын тексеру арқылы зерттеген.
Бұл әдіс неффилометр әдісі негізінде қарапайым. Суспензияда қанша көп жасуша болса, оның лайлылығы сонша жоғары болады. Сол қарайған жерден жасушаларды оңай санауға болады. Негізгі сатылымда бар броуновтық түтікшелерде қолданған ыңғайлы. Олардың суспензияларында сульфат, барий әртүрлі қоюлықта сұйықтан (түтікше №1) лайланғанға дейін (түтікше №10) болады.
Түтікшелердің жинағына кесте қосылады. Бұл қоблықтардың майлылығын кең арнада, әр түрлі микроорганизмде өлшейді. Сонымен бірге лайдан жарықтың өткізу қалыңдығын көруге болады .
Бұл кезде калорийметр мен спектро фотометрді қолданады. Негігі қызыл түсті қолдан,ған жақсы. Себебі ол дақылдың ортасында сары түспен араласқанда шағылыспайлы. Лайды өлшегенде, өсіп жатқан жасушалар үйіліп, жиналып қалса онда тексеру кезінде қате кетуі мүмкін.
12.6.3. Сапасыз әдістер
Микроорнаизмдерің өсуінің өлшеуін басқа әдістермен де өлшеуге болады. Мысалы, саңырауқұлақтарды уақыт өтісуімен мецилиде диаметрінің өсуімен өлшеуге болады.
Бұл әдіс температураның саңырауқұлақтарға қандай өзгеріс беретінін тексереді.
Егер саңырауқұлақты сұйық ортада өсірсе, белгілі уақыттан кейін оның дақылын сүзіп алуға болады.
12.7. Боялған бактериялар – Грамм бойынша бояу
Фазалық микроскоптың көмегімен тірі бактерияларды боялған түрінде анықтап көруге болады. Бұл әдісті Грамм бойынша бояу деп аталады. Бұл әдіс 1884 жылы даттық дәріге Кристшен Граммның табуымен аталды. Боялғанға дейін бұл бактериялар түссіз, боялғаннан кейін Грамм кәдімгі бактериялары күлгін түске айналды. Ал Граммға қарама – қарсы бактериялар қызыл түске боялады. Осы екі типтегі бактериялардың айырмашылығы 2.5.1. суретінде көрсетілген.
Түске бояу әдістері 12.2., 12.9. бөлімдерінде көрсетілген.
12.8 Вирустарды дақылдау
Вирустарды өсіру, бактерияларды, дақылдарды өсіруге қарағанда қиындау болады. Себебі вирустар тірі организмдер ішінде өседі. Вирусты тексеру арқылы бір бүтін өсімдікті немесе бір бүтін денені бүлдіріп алуы мүмкін. Сондықтан дақылдың белгілі бір жасушасын немесе тінін алып зерттейді. 12.11 суретінде көрсетілген.
Ертеде тауықтың ішінде өсіп жатқан жұмыртқаның дақылын алып зерттеген. Бұған ұқсас вирустар шошқа мойын мен тұмау ауруларын болдырпмау үшін егетін вакцина дайындаған. Жасуша дақылы бұл жасуша суспензиясының сұйық ортада өсуі. Ал тін дақылы бұл өсімдік немесе тірі организмнен алынған сұйық немесе қатты ортаны айтады. Жасуша дақылы мерристеманы қолданады. Бұл дегеніміз жасушалардың белсенді бөлінетін жері, материалды бүлінген өсімдіктен алады немесе дақылды керекті вируспен ауыртады. Тірі дақылдың жасушаларынан жұқа жасуша дайындап, соны зерттейді. Полиовирус 1-ші маймылдың бөтекесінен алынды. Тірі организмнің дақылын қолдану үшін өкпе жасушаларын обыр ауруымен ауырған адамның амнинонын тексеру арқылы зерттеген.

12.9. Лабараториялық жұмыс
Практика жұмыстарының мақсаты: кейбір бактериялық әдістерге арналған, сапасы қауіпсіз болғандықтан бактерия көзі-- сүтті пайдаланбақ болдық. Сүтқоректілер мен көптеген бактериялар үшін сүт керемет тағам.
12.9.1. Сүттегі бактерия құрамы
Қанша үлкен гигиеналық сақтықпен сауғанның өзінде сүтке бактериялар түседі. Балғын сүтті сауған бойда салқындатады. Сөйтіп бактериялардың көбеюуән азайтады. Өңделген сүтті пастерлейді. Ол дегенімі қайнатады. Сонда потогендік бактериялар өледі, ал пактогендік емес бактериялар тірі қалады.
Сүттегі бактериялар
Стрептококтар сүттің ашуы үшін қажет. Олардың формасы тізбек тәрізді болады.
Лактобалицус жасушасы таяқша пішінде болады. Олар оңай жабысып, ұзын тізбек жасайды. Сүтте бұдан басқа ішек бактерияларын кездестіруге болады. Соның ішінде ішек таяқшасын, алкогиминес. Бұл бактериялар жалғанған таяқша бола алады және тізбек құрайды.
Сүттен басқа да бактериялардың қатарын табуымызға болады, яғни оған ішекте өмір сүретін Alcaligenes таяқщасы(грамофты жағымсыз), бұл жасушалар бір өздері немесе тізбектп тұра алады. Олар әрбір колония айналасындағы сарғыш (сілтілік) аймағында агар Mакконки арқылы танылуы мүмкін.
Практика жұмыстарының мақсаты: кейбір бактериялық әдістерге арналған, сапасы қауіпсіз болғандықтан бактерия көзі-- сүтті пайдаланбақ болдық. Сүтқоректілер мен көптеген бактериялар үшін сүт керемет тағам.
12.9.1. Сүттегі бактерия құрамы
Қанша үлкен гигиеналық сақтықпен сауғанның өзінде сүтке бактериялар түседі. Балғын сүтті сауған бойда салқындатады. Сөйтіп бактериялардың көбеюуән азайтады. Өңделген сүтті пастерлейді. Ол дегенімі қайнатады. Сонда потогендік бактериялар өледі, ал пактогендік емес бактериялар тірі қалады.
Сүттегі бактериялар
- 15 – 30 °С та стрептокосус ( грамафонды тиімді болып келетін микрооргнаизмдер) және көптеген стрептококтар лактиздеу және тағы басқа бунақденелі бактериялар болады(мысалы, Microbacterium, Brevibacterium), олар Lactobacillusке ұқсайды. Алайда, таяқшаның арты жуандалады.
- 30-40°Ста әртүрлі лактасис, ішек бактериялары мысалы, Е,коли тірі болады.
Стрептококтар сүттің ашуы үшін қажет. Олардың формасы тізбек тәрізді болады.
Лактобалицус жасушасы таяқша пішінде болады. Олар оңай жабысып, ұзын тізбек жасайды. Сүтте бұдан басқа ішек бактерияларын кездестіруге болады. Соның ішінде ішек таяқшасын, алкогиминес. Бұл бактериялар жалғанған таяқша бола алады және тізбек құрайды.
Сүттен басқа да бактериялардың қатарын табуымызға болады, яғни оған ішекте өмір сүретін Alcaligenes таяқщасы(грамофты жағымсыз), бұл жасушалар бір өздері немесе тізбектп тұра алады. Олар әрбір колония айналасындағы сарғыш (сілтілік) аймағында агар Mакконки арқылы танылуы мүмкін.

12.9.2. Бактерияларға тәжірибе жұмыс
Бактериялық тәжірибелер бойынша төменгі 3әдіс микробиологиялық әдіс қолданылады.
1-ші әдіс сүт бактерияларының дақылын агар ыдысында өсіру. Бактерия өнімін штрихпен белгілеу.
2-ші әдіс Граммдау бойынша бояу. 12.1. суретке қараңдар
Бактериялардың бағаналарын қолдану мен бірге оларды неше түрге көбейтіміз.
Тәжірибе 12.1. Бактериялардың жаңа және ескі сүтте болуы
Тексеру барысында сүтті тәулігіне үй температурасында сақтағанда сүт неге ашып кететінін білу. Сүт адамдар үшін тағам, ал тексеру барысында сүт – көптеген бактериялардың қорегі болып шықты.
Материалдармен құралдар
*4тазаланған Петри ыдысы
*қоректік агары мен бірге
*микробиологиялық ілмек
*бунзеновтік қыздырғыш
*өшірілмейтін маркер
*балауыздан жасалған қарындаштар
*таза қайнатылған сүт
*бір тәулік бойы үй температурасында тұрған ескі сүт
*термостат 35°С
Әдістері:
*ілмекті бунзеновтік қыздырғышпен қыздырып, қыздырылғаншы ұстаймыз 12.4.
*ілмекті суытып содан соң жаңа сүтке батыру керек
*сүтке батырар алдында сүтті жақсылап былғап алу керек. Бос қолыңмен тазаланған агар қақпағын жайлап көтеріп ысыру керек.
* ал ілмек ішіндегісін агардың үстіне жаю керек.
12.4 суретін қара. Қақпағын жабамыз, сосын қайтадан ілмекті тесіп, қызған отқа қоямыз.
*Ыдыстың түбін кетпейтін маркермен қол қоямыз.
* немесе балауыыз қарындашымен
* осы жұмысты екінші ыдыспен келесі екінші сүтке де қайталаймыз.
*қайталадан ілмекті тесеміз, суытып ыдысқа көне сүтті құясыз.
*ілмек ішіндегіні 3 ші ыдыстың бетіне жағып, қақпағын жабамыз.
* түбін кетпейтін маркермен қол қоямыз. Дәл осы әдісті 4ші ыдыспен және әші ыдыспен көне сүтте қайталаймыз.
*барлық ыдысты термостатқа салып, 35°С қа қойып, 3 күнге инкубациялау қажет. Термостатқа ыдысты салмас бұрын, түбін жоғарыға аудару керек. Ол ыдыстың түбінде концентрация қалмас үшін жасалады. Инкубациядан кейін екі жарты жабыстырғыш лентаны дақыл жоғалмас үшін юапсырамыз.
* колонияның сыртқы сипатын жазыңыз.
Содан соң шыққан нәтижесімен салыстырыңыз.
Ескертулер:
*студенттер өздігінен стирильденген Маккортни ыдысын ерітілген агармен құя алады.
*Штрихтаған кезде бактериялардың саны жаймен азаяды.
*Бұл әдіспен жобадағы сүттің қанша бактерияны көп екенін айту керек. * Ыдыстар керек болмаса инкубациядан кейін тоңазытқышқа сақтауға болады. Салқындаған кезде бактериямыз өсуін тоқтатады. Ыдыстар керек болмаған жағдайда оларды 1 рет қолданатынпакетке салып, оны қатты қысымға ұстап, 15 мин ұстайды. Содан соң барып лақтырады.
* сүтпен тағы басқа да тәжірибелер жасауға болады. Ал істеген тәжірибеміз ең қарапайым. Сүттің құрамын сауған бойда тексеруге болады. Бұл үшін шикі сүтті стирильдегеныдыстарға құйып мақта пробкасымен жабады. Содан кейін 35 минуттай 63°С сулы буда ұстайды. Тәрірибенің үшінші нұсқасы кейбір ыдыстарды 10 °С-та инкубациялап, ұстау. Ал 35°С-та ұстаса стрептоуоккоуслактиз өседі. Ал лактабацилус өспейді.
Тәрірибе 12.2. бактерияларды бояу арқылы микроскоппен зерделеу.
Контрастық микроскоптың көмегімен тірі бактерияларды тексеруге болады.бірақ оларды алдын – ала бояп бітіру керек. Біз білетін бояу әдісі ГРАММ бойынша бояу қажет. Боялғанға дейін бактериялар түссіз, боялғаннан кейін қарапайым бактериялар күлгін түске айналады. Ал қарапайым емес бактериялар қызыл түске боялады. Осы 2 түрдегі бактериялардың айырмашылығы 2.3.1.суретінен қараңыз.
Материалдар мен құрал – жабдықтар:
Әдістер: Бірінші және алтыншы бөлімдері 5 минуттан артық бола алмайды. (Бактериология кітабынан алынды, Хамприес, Джон Мэррай, 1974ж. * Дайындалған әйнектің бетіне бактерия бояуларын дайындаңдар. Ол үшін сым ілгішін тесіп, қызған жалынға жалыннан өткіземіз және оны суытыңдар. 1-2тамшы суды әйнектің ортасына тамшылатыңдар. Жаңағы сым ілгішпен өздерің таңдаған бактериялық колонияны қозғаңдар. Ыдыстың қақпағын жай алыңдар. Себебі, дақылды бұзып алмау үшін. Жасушаны алып, бүлдіріп алмай шынының үстіне құямыз. Сонан соң ілгекпен жаймен араластырамыз. Жасушаларды шыны бетіне жаймен жағамыз, олардың аудандары 3*1см. Содан соң қайтадан ілмек қадаймыз. Сөйтіп, біз онша қалың да емес, онша жұқа да емес өзімізге керекті қалыңдықтағы жақпажасауымыз керек.оны бірнеше минутқа кептіріп қоямыз.
*бактерияларды бекітіңдер. Бактерияларды бекіту шыныны кішкентай қысқышпен алып, горизантальды түрде бунзендік жанғыштан өткіземіз. Шыны ысып кетпеу керек. Бактерияларды бекіткен кезде жасушалар коагуляция цитаплазмасы болып, өледі. Және шыныға жабысады.
*бактерияларды бояу. Бояу кезінде үстелді оңай бүлдіріп аламыз. Сол себепті бұл тәжірибені штативтің үстінде өткізген жақсы. Штативті 2 шыныдан немесе металлды таяқшадан горизанталь түрінде қойып, бір – біріне 5 см қашықтықта болады. Таяқшаларды пластилинмен бекітеді. Шыныларға криссталдың күлгін су құйып, 30секундқа қоямыз. Сөйтіп, бактериялар күлгін түске боялады.
*бояғыштарды Люголья ерітіндісімен шаямыз. Ол үшін шыныны осы ерітіндісімен 30 секундқа қоямыз. Йод жасуша ішіндегі бояғышты мықты жалғайды. *шыныларды ацетон – спирт ерітіндісімен бояуы кеткенше шаю керек. Егер бұл әдісті қайталау керек болса қайталаңыз. Істей бере – істей бере қолыңыз үйренеді. Жуғаннан кейін қарапайым емес бактериялар түссізденеді
*шыныға сафранин құйып, 3 минутқа құйыңыз. Бояцды сумен құямыз. Шыныны абайлап кептіріп, ауаға қоямыз. Сафранин қосымша бояғыш болып саналады. Ол криссталды күлгін түстен кейін қолданылады және бүкіл қарапайым емес бактерияларды қызыл түске бояйды.
*1 тамшы майды иммирсион майына тамызыңыз. Сосын объектив арқылы қараңыз.
Тәжірибе 12-3. Жаңа және ескірген сүт бактериялардың санын салыстыру
Егер таяқша бактерияларын агарға қондырсақ, сол кезде олар колония құрып бастайды. Оны жай көзбен қарауға болады. Құралдарды тазартамыз. 1-ші әдіс көбейту әдісіне арналған. Бактериялар саны сүтте көп. Сондықтан оны санау үшін ыңғайлы көлемдегі ыдыс алып жасау керек. 2-ші әдісте әр жасалған тәжірибеден өнімін алу керек. Бактерияларды санау үшін қай ыдыста инкубацияланды, қайсысында көптеген баған санын аламыз.
Материалдар мен құрал – жабдықтар
Қорек агарымен 6 стирильді тостаған
Сегіз дәйектелген 1 см3 тамшуыр
10см3 дәйектелген тамшуыр
6 түтік және арнайы штатив
Мақта
160 °С – тағы пеш
Өшпейтін маркер
Бунзендік оттық
100 см3 дистильденген су
Жаңа сүт
Ескірген сүт
70% спирт
Алюминий фольгасы
Шыны қалақшасы
Жабдықтарды залалсаздындыру
*әрбір 6шы ыдыстың қақпағын мақтамен жабыңыз.
*әрбір 6шы түтікшенің мақталы қақпағын жауып, алюминий фольгасымен орау керек. Әрбір 8 тамшуырдың ұшына 10 см3 кішкентай мақтаны бөлек орап, оны алюминий фольгаға орау.
*қақпақ пен мақтаны қатты ысытылған пешке 160 °С қа қыздырып, сол жерде 60 минут қоямыз.
*қолданар алдында құралдарды суытып алу керек.
Сүтті бағандап бөлу оған агар ыдысы арқылы жасуша егу.
*бір қолымызда қақпақты ұстап тұрып, екінші қолдарымызбен стирильденген түтікшені 10 см3 кольбадан 9,9см 3 ке толтырып, дистирильденген суды аламыз.
*кольбаны қақпақ пен жабамыз.
* осындай жолмен бірінші түтікшеден де қақпағын аламыз.
*9,9см3 суды түтікшеге аударамыз.
*Қақпақты жабамызқалған 5 түтікшемен де дәл осыны қайталаймыз.
* Жақсылап ыдыстағы даңа сүтті жақсылап шайқаймыз. Одан 1см3 түтікшемен аламыз. Оны 0,1см3 ыдыстағы сүтке құямыз С1. Қақпақты шешіп, қайтадан алдындағыдай толтырамыз. Осылай болу 100 рет қайталанады. Түтікшеден жайлап шайқаңыз. Сонда оның ішіндегісі дұрыстап араласады. * Жаңадан стирильденген түтікшені алып, 10 см3 С1 түтікшесіне сорамыз. Қақпағын сәл ашып қоямыз.
12.1. 3 және 4 тәжірибелеріндегі ескертпелерді қараңыз
12.9.3. Саңырауқұлақтармен жасалынатын практикалық жұмыс
Төменгі сападағы саңырауқұлақтармен жұмыс жасағанда стандраттрық микробиологиялық бактериямен жұмысындағыдай қолданылады. Көптеген сапраофиттік саңырауқұлақтар бактерия сияқты қоректік агарда өсіріледі. Ал егер таза саңырауқұлақтардың дақылдары керек болса, онда стирильденген жағдайда жұмыс жасалынады. 12.3 ,12.4 бөлімдерінде көрсетілген осындай долдармен мукор физопус пен пенициллинді өсіреді. Бұларға екі пайыздық агар жарайды. Олар петри ыдысына құйылған болу керек. Дақылдардың тағы бір бөлігін өсіруге болады
Олар : нанда, жемістерде, тағы басқа ылғал өнімдерде. Спораларды дәндә дақылдар өсетін ортаға ауыстырады. Дәнді дақылдарды сетраскоспиялық микроскоппен қараған жақсы.
12.10 Ауқымды өндірістікті
Өткен бөлімдерде микроорганизмдермен бірнеше әдістермен жұмыс жасадық.. лабаротриялық масштабтан өнеркәсіп масштабына өту үшін көптеген саллаларды қиындықтарын меңгеріміз керек. Оған биоинженерия, химия, биология жатады. Өндірістік бактерияларды өсіру кезінде оның экономикалық социалдық және этикалық жағын қарастыру керек. Бұл бөлімде ірі масттабты өндірістердің түрлі салаларына мысал келтіреміз.
Микроорганизмдерді өндірісті келесі себептер бойынша пайдалыналады.
12.10.1. Қарау
12.13 суретінде биотехнологиялық типтегі жұмыстар үшін диаграмма мен көрсетілген.
12.10.2. Скрининг
Микроорганизмдердің алуан түрлілігі химиялық реакцияла пай -даланып, өнімдерді жасай аламыз. Бірақ оның азғантай бөлімі өнеркәсіп өндірісінде пайдалыналады. Дәрі дәрмек шығаратын коммерциялық компаниялар микроорганизмдердің пайдалы жағын тапқысы келіп, ылғи ізденіс үстінде болады. Сонымен олар жаңа өнім тапқысы келіп, жаңа дақылдарды жинап жұмыстанады. Көп жағдайда эмпирикалық жұмыс жаңалықтар ашқанда маңызды рөл ойнайды. Микроорганизмидерді тексеруді Скрининг деп атайды. Жаңа антибиотик іздеп табу үшін жасалынатын Скрининг бұған жақсы мысал бола аладаы. Алғашқы аинтибиотик 1928 жылы Александар Флемингінің атымен аталып, пенициллин атын алып ашылған. Табиғаттық антибиотиктер бұлар химиялық заттар. Микроорганизмдермен синтезделеді. Бірақ басқа микроорганизм тоқтатып өлтіреді. 1928 жылдан бастап, микроорганиздерден 5000 астам түрлі антибиотиктер соның ішінде әр түрлі топтағы пенициллиндер шығарылды. Бұлардың көбі медициналық мақсатқа жарамсыз болып табылды. Себебі, олар жоғарғы улы зат еді. Бірақ, стрептомицин өкілдерінің көздері антибиотиктің өте бай көзі болып табылды.
Бактериялық тәжірибелер бойынша төменгі 3әдіс микробиологиялық әдіс қолданылады.
1-ші әдіс сүт бактерияларының дақылын агар ыдысында өсіру. Бактерия өнімін штрихпен белгілеу.
2-ші әдіс Граммдау бойынша бояу. 12.1. суретке қараңдар
Бактериялардың бағаналарын қолдану мен бірге оларды неше түрге көбейтіміз.
Тәжірибе 12.1. Бактериялардың жаңа және ескі сүтте болуы
Тексеру барысында сүтті тәулігіне үй температурасында сақтағанда сүт неге ашып кететінін білу. Сүт адамдар үшін тағам, ал тексеру барысында сүт – көптеген бактериялардың қорегі болып шықты.
Материалдармен құралдар
*4тазаланған Петри ыдысы
*қоректік агары мен бірге
*микробиологиялық ілмек
*бунзеновтік қыздырғыш
*өшірілмейтін маркер
*балауыздан жасалған қарындаштар
*таза қайнатылған сүт
*бір тәулік бойы үй температурасында тұрған ескі сүт
*термостат 35°С
Әдістері:
*ілмекті бунзеновтік қыздырғышпен қыздырып, қыздырылғаншы ұстаймыз 12.4.
*ілмекті суытып содан соң жаңа сүтке батыру керек
*сүтке батырар алдында сүтті жақсылап былғап алу керек. Бос қолыңмен тазаланған агар қақпағын жайлап көтеріп ысыру керек.
* ал ілмек ішіндегісін агардың үстіне жаю керек.
12.4 суретін қара. Қақпағын жабамыз, сосын қайтадан ілмекті тесіп, қызған отқа қоямыз.
*Ыдыстың түбін кетпейтін маркермен қол қоямыз.
* немесе балауыыз қарындашымен
* осы жұмысты екінші ыдыспен келесі екінші сүтке де қайталаймыз.
*қайталадан ілмекті тесеміз, суытып ыдысқа көне сүтті құясыз.
*ілмек ішіндегіні 3 ші ыдыстың бетіне жағып, қақпағын жабамыз.
* түбін кетпейтін маркермен қол қоямыз. Дәл осы әдісті 4ші ыдыспен және әші ыдыспен көне сүтте қайталаймыз.
*барлық ыдысты термостатқа салып, 35°С қа қойып, 3 күнге инкубациялау қажет. Термостатқа ыдысты салмас бұрын, түбін жоғарыға аудару керек. Ол ыдыстың түбінде концентрация қалмас үшін жасалады. Инкубациядан кейін екі жарты жабыстырғыш лентаны дақыл жоғалмас үшін юапсырамыз.
* колонияның сыртқы сипатын жазыңыз.
Содан соң шыққан нәтижесімен салыстырыңыз.
Ескертулер:
*студенттер өздігінен стирильденген Маккортни ыдысын ерітілген агармен құя алады.
*Штрихтаған кезде бактериялардың саны жаймен азаяды.
*Бұл әдіспен жобадағы сүттің қанша бактерияны көп екенін айту керек. * Ыдыстар керек болмаса инкубациядан кейін тоңазытқышқа сақтауға болады. Салқындаған кезде бактериямыз өсуін тоқтатады. Ыдыстар керек болмаған жағдайда оларды 1 рет қолданатынпакетке салып, оны қатты қысымға ұстап, 15 мин ұстайды. Содан соң барып лақтырады.
* сүтпен тағы басқа да тәжірибелер жасауға болады. Ал істеген тәжірибеміз ең қарапайым. Сүттің құрамын сауған бойда тексеруге болады. Бұл үшін шикі сүтті стирильдегеныдыстарға құйып мақта пробкасымен жабады. Содан кейін 35 минуттай 63°С сулы буда ұстайды. Тәрірибенің үшінші нұсқасы кейбір ыдыстарды 10 °С-та инкубациялап, ұстау. Ал 35°С-та ұстаса стрептоуоккоуслактиз өседі. Ал лактабацилус өспейді.
Тәрірибе 12.2. бактерияларды бояу арқылы микроскоппен зерделеу.
Контрастық микроскоптың көмегімен тірі бактерияларды тексеруге болады.бірақ оларды алдын – ала бояп бітіру керек. Біз білетін бояу әдісі ГРАММ бойынша бояу қажет. Боялғанға дейін бактериялар түссіз, боялғаннан кейін қарапайым бактериялар күлгін түске айналады. Ал қарапайым емес бактериялар қызыл түске боялады. Осы 2 түрдегі бактериялардың айырмашылығы 2.3.1.суретінен қараңыз.
Материалдар мен құрал – жабдықтар:
- Негізгі бояғыштар – криссталды күлгін (0,5% су ерітіндісі)
- Бояуды жылтыратушы зат - Люголь ерітіндісі
- Ацетон – түссіздену үшін қолданылатын спирт(50 де 50 абсолюттік спирт болатын ацетон)
- Қосымша бояғыш - сафранин (1 пайыздық су ерітіндісі)
- Сымды Бунзендік оттықтың ілгегі
- Әйнектерді жақсылап жуады (спиртпен сүртеді)
- Қысқыш
- Бояу үшін штативті астауша немесе тостағанша орналастырамыз
- Сүзгіш қағазды дистильденген сумен шаю
- Иммерсионды маймен және иммерсионды объективті микроскоп.
Әдістер: Бірінші және алтыншы бөлімдері 5 минуттан артық бола алмайды. (Бактериология кітабынан алынды, Хамприес, Джон Мэррай, 1974ж. * Дайындалған әйнектің бетіне бактерия бояуларын дайындаңдар. Ол үшін сым ілгішін тесіп, қызған жалынға жалыннан өткіземіз және оны суытыңдар. 1-2тамшы суды әйнектің ортасына тамшылатыңдар. Жаңағы сым ілгішпен өздерің таңдаған бактериялық колонияны қозғаңдар. Ыдыстың қақпағын жай алыңдар. Себебі, дақылды бұзып алмау үшін. Жасушаны алып, бүлдіріп алмай шынының үстіне құямыз. Сонан соң ілгекпен жаймен араластырамыз. Жасушаларды шыны бетіне жаймен жағамыз, олардың аудандары 3*1см. Содан соң қайтадан ілмек қадаймыз. Сөйтіп, біз онша қалың да емес, онша жұқа да емес өзімізге керекті қалыңдықтағы жақпажасауымыз керек.оны бірнеше минутқа кептіріп қоямыз.
*бактерияларды бекітіңдер. Бактерияларды бекіту шыныны кішкентай қысқышпен алып, горизантальды түрде бунзендік жанғыштан өткіземіз. Шыны ысып кетпеу керек. Бактерияларды бекіткен кезде жасушалар коагуляция цитаплазмасы болып, өледі. Және шыныға жабысады.
*бактерияларды бояу. Бояу кезінде үстелді оңай бүлдіріп аламыз. Сол себепті бұл тәжірибені штативтің үстінде өткізген жақсы. Штативті 2 шыныдан немесе металлды таяқшадан горизанталь түрінде қойып, бір – біріне 5 см қашықтықта болады. Таяқшаларды пластилинмен бекітеді. Шыныларға криссталдың күлгін су құйып, 30секундқа қоямыз. Сөйтіп, бактериялар күлгін түске боялады.
*бояғыштарды Люголья ерітіндісімен шаямыз. Ол үшін шыныны осы ерітіндісімен 30 секундқа қоямыз. Йод жасуша ішіндегі бояғышты мықты жалғайды. *шыныларды ацетон – спирт ерітіндісімен бояуы кеткенше шаю керек. Егер бұл әдісті қайталау керек болса қайталаңыз. Істей бере – істей бере қолыңыз үйренеді. Жуғаннан кейін қарапайым емес бактериялар түссізденеді
*шыныға сафранин құйып, 3 минутқа құйыңыз. Бояцды сумен құямыз. Шыныны абайлап кептіріп, ауаға қоямыз. Сафранин қосымша бояғыш болып саналады. Ол криссталды күлгін түстен кейін қолданылады және бүкіл қарапайым емес бактерияларды қызыл түске бояйды.
*1 тамшы майды иммирсион майына тамызыңыз. Сосын объектив арқылы қараңыз.
Тәжірибе 12-3. Жаңа және ескірген сүт бактериялардың санын салыстыру
Егер таяқша бактерияларын агарға қондырсақ, сол кезде олар колония құрып бастайды. Оны жай көзбен қарауға болады. Құралдарды тазартамыз. 1-ші әдіс көбейту әдісіне арналған. Бактериялар саны сүтте көп. Сондықтан оны санау үшін ыңғайлы көлемдегі ыдыс алып жасау керек. 2-ші әдісте әр жасалған тәжірибеден өнімін алу керек. Бактерияларды санау үшін қай ыдыста инкубацияланды, қайсысында көптеген баған санын аламыз.
Материалдар мен құрал – жабдықтар
Қорек агарымен 6 стирильді тостаған
Сегіз дәйектелген 1 см3 тамшуыр
10см3 дәйектелген тамшуыр
6 түтік және арнайы штатив
Мақта
160 °С – тағы пеш
Өшпейтін маркер
Бунзендік оттық
100 см3 дистильденген су
Жаңа сүт
Ескірген сүт
70% спирт
Алюминий фольгасы
Шыны қалақшасы
Жабдықтарды залалсаздындыру
*әрбір 6шы ыдыстың қақпағын мақтамен жабыңыз.
*әрбір 6шы түтікшенің мақталы қақпағын жауып, алюминий фольгасымен орау керек. Әрбір 8 тамшуырдың ұшына 10 см3 кішкентай мақтаны бөлек орап, оны алюминий фольгаға орау.
*қақпақ пен мақтаны қатты ысытылған пешке 160 °С қа қыздырып, сол жерде 60 минут қоямыз.
*қолданар алдында құралдарды суытып алу керек.
Сүтті бағандап бөлу оған агар ыдысы арқылы жасуша егу.
- Алты стирильденген жабық түтікпен қымталған түтікті белгілейміз : С1,С2,С2,Н1,Н2,Н3. Алюминий қақпағын шешеміз.
- Қорек агары бар стирильденген ыдыстардың түбіне қасына қоямыз С1,С2,С2,Н1,Н2,Н3.
- Әрбір алты түтікшеге тазаланған дистильденген суды құйыңыз. Ол үшін келесі мысалға қараңыз:
*бір қолымызда қақпақты ұстап тұрып, екінші қолдарымызбен стирильденген түтікшені 10 см3 кольбадан 9,9см 3 ке толтырып, дистирильденген суды аламыз.
*кольбаны қақпақ пен жабамыз.
* осындай жолмен бірінші түтікшеден де қақпағын аламыз.
*9,9см3 суды түтікшеге аударамыз.
*Қақпақты жабамызқалған 5 түтікшемен де дәл осыны қайталаймыз.
* Жақсылап ыдыстағы даңа сүтті жақсылап шайқаймыз. Одан 1см3 түтікшемен аламыз. Оны 0,1см3 ыдыстағы сүтке құямыз С1. Қақпақты шешіп, қайтадан алдындағыдай толтырамыз. Осылай болу 100 рет қайталанады. Түтікшеден жайлап шайқаңыз. Сонда оның ішіндегісі дұрыстап араласады. * Жаңадан стирильденген түтікшені алып, 10 см3 С1 түтікшесіне сорамыз. Қақпағын сәл ашып қоямыз.
- Шпательді қайнатыңыз. Ол үшін оны 70 пайыз спиртке батырамыз. Алғасын спирттің аққванын күтеміз. Одан соң оны вертикалды ұстап тұрып, бунзендік қыздырғыштың жалынына ұстаймыз.
- * шпательдің сууын күту керек. Суыған шпательдің шпательмен сүттің бетін кесіңіз.
- Қайтадан Шпательді тазалаңыз.
- Жоғарыдағы түтікшені қолдана отырып, 0,1см3 С1 түтікшемен алып, С2 түтікшесіне салып жоғарыдағы көрсетілгендей мақта тығынымен орнына қоямыз. * С2 түтікшесін дұрыстап шайқаймыз. Одан 10000 рет бөлінеді.
- 10- сы мен 11-шідегі С2нің орнына С3ті пайдаланып, жоғарыдағы тәжірибені қайталаңыз. Ол 1 000000 рет бөлінеді.
- 6-шы пунктегі С3тің орнына С1ді қайталаңыз.
- Көне сүттен Н1, Н2,Н3 ыдыстарын алып, жоғарыдағы тәжірибелерді қайталаңыз. Алдыңғы 6 ыдысты түбімен жоғарыдағыдай айырып, термостатқа 35 °Сқа қойып, жобамен 3 тәулік тастаймыз.
- * потогенді микроорганизмдердің жойылып кетуінен сақтану үшін қақпағын жабыңыз, лентамен жабыстырыңыз.
- Бүкіл ыдыстағы бактерияның өсуін бағалаңыз. Әр колонияның санын бөлек бағалаңыз. Шыққан нәтижелерін санап, жазыңыз. Бактерия санын 1см3 араласпаған сүтпен есептейміз және кесте түрінде жазамыз.
12.1. 3 және 4 тәжірибелеріндегі ескертпелерді қараңыз
12.9.3. Саңырауқұлақтармен жасалынатын практикалық жұмыс
Төменгі сападағы саңырауқұлақтармен жұмыс жасағанда стандраттрық микробиологиялық бактериямен жұмысындағыдай қолданылады. Көптеген сапраофиттік саңырауқұлақтар бактерия сияқты қоректік агарда өсіріледі. Ал егер таза саңырауқұлақтардың дақылдары керек болса, онда стирильденген жағдайда жұмыс жасалынады. 12.3 ,12.4 бөлімдерінде көрсетілген осындай долдармен мукор физопус пен пенициллинді өсіреді. Бұларға екі пайыздық агар жарайды. Олар петри ыдысына құйылған болу керек. Дақылдардың тағы бір бөлігін өсіруге болады
Олар : нанда, жемістерде, тағы басқа ылғал өнімдерде. Спораларды дәндә дақылдар өсетін ортаға ауыстырады. Дәнді дақылдарды сетраскоспиялық микроскоппен қараған жақсы.
12.10 Ауқымды өндірістікті
Өткен бөлімдерде микроорганизмдермен бірнеше әдістермен жұмыс жасадық.. лабаротриялық масштабтан өнеркәсіп масштабына өту үшін көптеген саллаларды қиындықтарын меңгеріміз керек. Оған биоинженерия, химия, биология жатады. Өндірістік бактерияларды өсіру кезінде оның экономикалық социалдық және этикалық жағын қарастыру керек. Бұл бөлімде ірі масттабты өндірістердің түрлі салаларына мысал келтіреміз.
Микроорганизмдерді өндірісті келесі себептер бойынша пайдалыналады.
- Микроорганизмдер қарапайым қоректік талаптары бар
- Ферменттерде үлкен ыдыстағы өсіп атқан микроорганизмдеріңң өсі сапасын діл бақылауға болады.
- Микроорганизмдер тез өсу әдетімен ерекшеленнеді.
- Реакцияларын төменгі температурада жүргізуге
- Микроорганизмдер тағамның өте жоғарғы сапада болуын кірістіреді.
- Химиялық қосындылардың кең аумағын пайдалануға болады.
- Қиын химиялық қосындылары бар гармондар мен антибиотиктерді шығаруға болады.
- Микроорганизмнің генетикасы қарапайым сондықтан генетикалық монипулияциялық әдістері ылғи дамып отырады
12.10.1. Қарау
12.13 суретінде биотехнологиялық типтегі жұмыстар үшін диаграмма мен көрсетілген.
12.10.2. Скрининг
Микроорганизмдердің алуан түрлілігі химиялық реакцияла пай -даланып, өнімдерді жасай аламыз. Бірақ оның азғантай бөлімі өнеркәсіп өндірісінде пайдалыналады. Дәрі дәрмек шығаратын коммерциялық компаниялар микроорганизмдердің пайдалы жағын тапқысы келіп, ылғи ізденіс үстінде болады. Сонымен олар жаңа өнім тапқысы келіп, жаңа дақылдарды жинап жұмыстанады. Көп жағдайда эмпирикалық жұмыс жаңалықтар ашқанда маңызды рөл ойнайды. Микроорганизмидерді тексеруді Скрининг деп атайды. Жаңа антибиотик іздеп табу үшін жасалынатын Скрининг бұған жақсы мысал бола аладаы. Алғашқы аинтибиотик 1928 жылы Александар Флемингінің атымен аталып, пенициллин атын алып ашылған. Табиғаттық антибиотиктер бұлар химиялық заттар. Микроорганизмдермен синтезделеді. Бірақ басқа микроорганизм тоқтатып өлтіреді. 1928 жылдан бастап, микроорганиздерден 5000 астам түрлі антибиотиктер соның ішінде әр түрлі топтағы пенициллиндер шығарылды. Бұлардың көбі медициналық мақсатқа жарамсыз болып табылды. Себебі, олар жоғарғы улы зат еді. Бірақ, стрептомицин өкілдерінің көздері антибиотиктің өте бай көзі болып табылды.
Антибиотиктер адам мен жануарлардың бактериалдық немесе саңырауқұлақтық ауруларын емдеуге қолданды. Олардың кейбіреулері рак ауруларын тоқтатты. Сондықтан антибиотиктер екінші қатардағы метаболизм өнімдеріне жатады
12.10.3 Өндіріс ауқымын кеңейту
Кез келген жаңа биотехнологиялық өндіріс алдымен лабараториялық масштабта өткізілген дұрыс. Кәдімгі лабараторияда зерттелгенннен кейін өндірістік әдістеме құрал – жабдықтарымен қояды. Бұл жерде көлемі 2 ден 200 дм3 –ге дейін болатын ферменттер қолданылады. Ферментер дегеніміз жұмыс жүретін резеруар. Жасалынған өнімнің көп болуы үшін оған физикалық қажеттіктерін және өнімдік қажеттіліктерін қамтамасыз ету керек. Жұмыс көлемін үлкейтіп әдістеме өндірістен үлкен масштабтағы өндіріске өткізу үшін бірнеше дм3 аймақ керек. 12.15. суретін қараңыз.
12.10.3 Өндіріс ауқымын кеңейту
Кез келген жаңа биотехнологиялық өндіріс алдымен лабараториялық масштабта өткізілген дұрыс. Кәдімгі лабараторияда зерттелгенннен кейін өндірістік әдістеме құрал – жабдықтарымен қояды. Бұл жерде көлемі 2 ден 200 дм3 –ге дейін болатын ферменттер қолданылады. Ферментер дегеніміз жұмыс жүретін резеруар. Жасалынған өнімнің көп болуы үшін оған физикалық қажеттіктерін және өнімдік қажеттіліктерін қамтамасыз ету керек. Жұмыс көлемін үлкейтіп әдістеме өндірістен үлкен масштабтағы өндіріске өткізу үшін бірнеше дм3 аймақ керек. 12.15. суретін қараңыз.


Негізгі проблема – тазық жағдайының ұсталуы. Ферментерді ластап лау өте оңай, сондықтан оны үнемі бақылап отрыу керек. Инженерлік технологтар асептикалық әдістерді қолдана отырып, прутин деп аталатын бір жасушалы нәруыздарды іздеп, тәжірибелер арқылы өте жақсы жеңістіктерге жетті.
Ферментер және биореактор сұйық және қатты ортада өсірілетін микроорганхмдерді қадағалайтын жұмысы бар. Ферменттерде жүргізілетін жұмыс ферментация деп аталады. Алғашқыда ферментация тек анаэробтық жұмыстарға арналған, бірақ кейін ол анаэробпен қоса аэробқа да арналған кеңінен қолданатын жұмыс болып табылды. 12.16 – да ферментер көрсетілген. Бұл өте техниканың қиын құрылыс, сондықтан бұні жасату үшін көп уақыт керек. Жұмыс кезінде ферменттердің құрамы әр түрлі жағдайда орналасады. Мысалы, бір жасушалы прутина уызын араластыру ауа арқылы және жоғалу жылдамдықта жасалады. Өнім болып жасушалардың биомассасы немесе пайдалы жасуша метоболиты болады. Барлық операциялар стириьлденген жағдайда дән – дақылды ластамай жүргізеді.
Фермент құрал жабдықтары стирильденген болу керек. Сондықтан оны жұмысқа қосар алдында бөлшектеп стирильдейді. Ферментация кезінде басқа организмнен қосылатын дәнді дақыл белсенді емес түрінде болады(мысалы, қатырған түрде). Ыдыстағы затты белсенділігін арттырып, керекті мөлшерде өсіреді. Одан кейі оған фермент қосады. Ферменттерде оганизм қоректі ортадан өсіп көбейеді.
Негізі, ферменттерді даттанбайтын, сапасы жоғары металлдан жасайды. Себебі, жұмыс барысында тұзды орталарда, улы заттар бөлініп, темірді шерітпес үшін барлық қолданатын құралдар материалдар ауада стирильдеу керек. Құралды жоғарғы қысымдағы бумен стирильді, бу құралдары барлық жеріне тиюі керек. Жұмысқа қосар алдында ортаны салқынмен энокулияциялайды.
12.10.5. субстратты дақылға қосумен үздіксіз жұмыс жасау
Ферменттің 2 негізгі түрлер бар. Үздіксіз ферментациялау, пакетті ферментация кезде керек қоспалар жұмыс басында қосылады. Бұл кезде қоректік затта ашыту барысында қосылмайды. Сондықтан фермент параметрлері өлшенеді. Керекті өнімдер қосылып болғаннан кейін, жұмысты тоқтатады. Содан соң фермент босатады. Өнім алады, қолданылған микроорганизмдерді тастайды. Ферменттерді тазалап, келесі жұмысқа дайындайды. Ал үздіксіз ферменттерді өнімдеу жұмысы ұзақ уақыт алады, көптеген апта уақыт кетеді. Бұл кезде қоректік заттарды, керегінше ортаға жұмсайды,ал дақылдың артығын алып тастайды. Қиындығына қарамай бұл тәсіл негізгі жағдайға жақын келеді. Мысалы, бір компания бір жасушалы прутинаны бактериялардан алып, осылай дайындайды. Фермент тоқтамастан 100 кун жұмыс жасайды. Ал күніне 150 000т өнім алады. Өнімге субстрат қосу арқылы үздіксіз жұмыс дасау, жұмыстың бақылауын жеңілдетеді, сіңімді заттарды төменгі қоблықтарды қосу арқылы өсі уақыты ұзарады. Негізгі тез өсіру әдісі үшін оттегінің кең көлемде жеткізілуінің маңызы зор. Бұндай жұмыс ашытқылы жұмысын қолданғанда пайдаланады. Егер ашытқылы жасушаға қантты көп қосса анаэробаны демалады. Сөйтіп биологиялық массаның орнына спирт жиналады. Үздіксіз субстраттарды қосу арқылы дақылдау пенициллин шығарылуда қолданады. 12.11.1 бөлімдерінде көрсетілген.
Үздіксіз және пакеттеп өнімдеудің пайдасы мен зияны
- Бір саладан екінші масштабтағы салаға көшу үлкен физикалық факторларға мысалы, араласуға аэрацияға әкеліп, ортаны қызып кетуден сақтайды. Бұл проблемаларды инженер химиктер шешеді.
- Дақылдарға жақсы оттегі жету үшін ортаны аэриллеп, қағу, жеткіліксіз. Кішкентай көпіршіктер өте тиімді. Сондықтан ауа беру үшін трубкадан шашатын шашқыштарды пайдаланады. Турвалентіктің жоғару болуы үшін отегінің бүкіл ортаға тарауы үшін көпіршік шашатын бөлімдерін көбейтеді.
- Көпіршіктерді азайту үшін көпіршік бастаныды қолданады. Үлкен масштабтағы өндірістерде микроорганизмдер белсенді жұмыс жасауы үшін жылу бөлінеді. Сондықтан ферменттеррді салқындату үшін су серклияциясы болу қажет.
- Үлкен масштабты өндірісте қолайлы орта жағдайын ондағы қоректік заттардың отегінің, рҺ ты сақтап тұру өте қиын. Бұл қиындықтарды мән бермей өту көптеген қолайсыздықтарға әкеліп соқтырады. Прутина өндірісінде алғашқы өнімдерінің шығуы берген келісіміне қарамастан өте аз шығарылды. Бұл жағдайды ферментттерге метанол қоса отырып, жөндеді. Метанол оттегі мен қуат көзі болып саналды. Метанолды қосқаннан кейін өнімдер ойдағыдай жақсы өсті. Микрооргнаизмдер қоршаған жағдайға байланысты үлкен масштабтарда бақылау қиын. Кез келген байқаусыз өзгерістер адамға немесе жануарларға зиянды болуы мүмкін.
- Құрал жабдықтармен жасалу әдістері қиын болған сайын олардың апаттарымен қателері көп болады. Сондықтан шешім қабылдамас бұрын экономикалық жобасын дұрыс бақылап алу керек.
- Улкен масштабты өндірістерде стирильдеу үшін көп көлемде су кетеді.
- Көп мөлшерде пайдалынылатын материалдар жақын жерде болуы керек. Олардың қоры үзілмеуі керек, ыңғайлы жағдайда саақталуы керек. Химиялық өзгеріске ұшырайтын материалдар аз болуы керек. Қолданатын реогенттердің бастапқы бағасы аррзан болуы қажет бұл ең керек фактордардың бірі. Кейбір таңдалған микроорганизмдер ферментативті пайдалану ісінде төмен өнім шығарады. Үлкен масштабты өндірістерде мундай жағдайлар көп болады.
- Түйіршік түрінде жасалынған заттар жұмыс кезінде жұмысшылар үшін өміріне қауіа туғызуы мүмкін.
- Ферменттер жұмысындағы материалдар карозияға төзімді болу керек. Сонымен бірге микроорганизмдер болу үшін улы емес тазалауға төзімді болу керек.
- Үлкен мастабтағы ферментация кезінде гендік мадифицирленген организмдер қосымша қауіп туғызады. Олар адамдар үшінде өсірілген микроорганизм үшінде биологиялық белсенді өнім болып табылады.
- Кейде микробтық штаммдар ферментативті процесінде пайдалану үшін таңдалған, мутация арқылы оңай қайтаруға және бұрынғыдан да тез өсетіндей төмегі өнім штаммына айнала алады. Кеңейту кезінде осындай оқиғалардың ықтималдығы өндірістікте айтарлықтай арттырады.
- көп мөлшерде қолданылатын ұнтақ заттар, қызметкерлердің денсаулығына қауіпті болуы мүмкін.
- Ферментер құрылысына пайдаланылатын материалдар, металл із көлемімен ластануын болдырмау үшін коррозияға төзімді болуы тиіс. Сонымен қатар, олар микроорганизмдерге улы болмау керек және жоғарғы қысымнан залалсыздандыру керек
- Ауқымды ашыту 1қосымша тәуекел, әсіресе пайдаланылатын генетикалық түрлендiрiлген организмдер. Олар кәдімгі зертханалық ауқымда салыстырғанда, дақыл микроорганизмдердің ретінде адамдарға неғұрлым қуатты әсер, немесе биологиялық белсенді өнім мүмкіндігі туғызған.
Ферментер және биореактор сұйық және қатты ортада өсірілетін микроорганхмдерді қадағалайтын жұмысы бар. Ферменттерде жүргізілетін жұмыс ферментация деп аталады. Алғашқыда ферментация тек анаэробтық жұмыстарға арналған, бірақ кейін ол анаэробпен қоса аэробқа да арналған кеңінен қолданатын жұмыс болып табылды. 12.16 – да ферментер көрсетілген. Бұл өте техниканың қиын құрылыс, сондықтан бұні жасату үшін көп уақыт керек. Жұмыс кезінде ферменттердің құрамы әр түрлі жағдайда орналасады. Мысалы, бір жасушалы прутина уызын араластыру ауа арқылы және жоғалу жылдамдықта жасалады. Өнім болып жасушалардың биомассасы немесе пайдалы жасуша метоболиты болады. Барлық операциялар стириьлденген жағдайда дән – дақылды ластамай жүргізеді.
Фермент құрал жабдықтары стирильденген болу керек. Сондықтан оны жұмысқа қосар алдында бөлшектеп стирильдейді. Ферментация кезінде басқа организмнен қосылатын дәнді дақыл белсенді емес түрінде болады(мысалы, қатырған түрде). Ыдыстағы затты белсенділігін арттырып, керекті мөлшерде өсіреді. Одан кейі оған фермент қосады. Ферменттерде оганизм қоректі ортадан өсіп көбейеді.
Негізі, ферменттерді даттанбайтын, сапасы жоғары металлдан жасайды. Себебі, жұмыс барысында тұзды орталарда, улы заттар бөлініп, темірді шерітпес үшін барлық қолданатын құралдар материалдар ауада стирильдеу керек. Құралды жоғарғы қысымдағы бумен стирильді, бу құралдары барлық жеріне тиюі керек. Жұмысқа қосар алдында ортаны салқынмен энокулияциялайды.
12.10.5. субстратты дақылға қосумен үздіксіз жұмыс жасау
Ферменттің 2 негізгі түрлер бар. Үздіксіз ферментациялау, пакетті ферментация кезде керек қоспалар жұмыс басында қосылады. Бұл кезде қоректік затта ашыту барысында қосылмайды. Сондықтан фермент параметрлері өлшенеді. Керекті өнімдер қосылып болғаннан кейін, жұмысты тоқтатады. Содан соң фермент босатады. Өнім алады, қолданылған микроорганизмдерді тастайды. Ферменттерді тазалап, келесі жұмысқа дайындайды. Ал үздіксіз ферменттерді өнімдеу жұмысы ұзақ уақыт алады, көптеген апта уақыт кетеді. Бұл кезде қоректік заттарды, керегінше ортаға жұмсайды,ал дақылдың артығын алып тастайды. Қиындығына қарамай бұл тәсіл негізгі жағдайға жақын келеді. Мысалы, бір компания бір жасушалы прутинаны бактериялардан алып, осылай дайындайды. Фермент тоқтамастан 100 кун жұмыс жасайды. Ал күніне 150 000т өнім алады. Өнімге субстрат қосу арқылы үздіксіз жұмыс дасау, жұмыстың бақылауын жеңілдетеді, сіңімді заттарды төменгі қоблықтарды қосу арқылы өсі уақыты ұзарады. Негізгі тез өсіру әдісі үшін оттегінің кең көлемде жеткізілуінің маңызы зор. Бұндай жұмыс ашытқылы жұмысын қолданғанда пайдаланады. Егер ашытқылы жасушаға қантты көп қосса анаэробаны демалады. Сөйтіп биологиялық массаның орнына спирт жиналады. Үздіксіз субстраттарды қосу арқылы дақылдау пенициллин шығарылуда қолданады. 12.11.1 бөлімдерінде көрсетілген.
Үздіксіз және пакеттеп өнімдеудің пайдасы мен зияны
Сурет 12.16. Типті ферменттер. Сауыт мөлшері 1 дм3-500000дм3 дейін болад немесе өседі. Берілген құрал – материалдармен олардың әртүрлі болады.
Фермент көбіне тат баспайтын темірден цилиндр түрінде жатады.
Шартты белгілер:
Ферментация болғаннан кейін, өнімді ортадан шығарылу тазалау керек. Негізінде, ферменттердің құрал алдыңғы бөлшектер, сұйық және қатты жасушалары бар бөлшектерді бөледі. Бұл фильтрациямен центрифугация көмегімен жүреді. Керекті өнім сұйық түрінде болады не жасуша ішінде болады. тазалағанның биохимиялық тазалау неше түрі болады: бұған, кептіру, хромотография, еріткіш экстракциямен дестициялау, ары қарай өңдеу жұмысын инсулин шығатын завод 90% - ы 200 адамнан құралған жұмысымен жасайды. Пенициллинмен микропротеин өңдеу жұмысын жасалады. 12.11.1 және 12.12.3 суреттерінде.
12.4. Кесте. Үздіксіз және периодтық өңдеудің пайдасы мен зияны.
12.11.1. Пеницилмен шығарылуы
Пенициллин антибиотиктерін шығару үздіксіз шығару жақсы үлгіс.
Антибиотик дегеніміз негізгі
Пенициллин саңырауқұлағын дерттеу барысында, алғашқы болып АҚШ тың Дүниежүзілік соғыстарында қолданбақ болды. Соған қарамастан Флеминг пенициллинді Англияда ашты. Бұл ізденіс Оксфордуниверситетінің сэрі Говардом Флори қолдауымен болды. Бұл жерде өнімдердің неше түрлері үлгідегі сапасы тексерілдң. Бірақ, пенициллин жоғарғы орын алды. Содан бері қарай жоғарғы өнімді мутаттық штамптарда, сонымен бірге, өнімді өсіру, оларды бөліп шығару және тазалағанда өнімі 2000 есе өсті.ортаның шешуші мәнін ферментация жұмысы шешті. Керек көміртегімен глюкоза бұларға, ең керек көзі болып табылады. Глюкоза саңырауқұлақтың өсуін қамтамасыз етеді. Бірақ, ферменттін өнуін басады. Сондықтан, алдымен 30 – 40 сағ саңырауқұлақтың тез өсуін қамтамасыз етеді. Ал содан сң глюкоза ең төменгі концентрацияда бақылай отырып қосады. Азот көзі болып, арзан ақуыз алынады. Мысалы,соялық және балық ұны. Сонымен бірге, фенилуксустық қышқыл қажет. Пеницилин екіншіші метаболит болғандығтан ол бірінші тез өсетін фазадан кейін қосады. Бірінші метаболитке демалу өнімдері көмірмен этиль спирті жатады. Өсетін өнімді саңырауқұлақ жиналғаннан алады. 50000дм3 аймақтағы иникулияция ферменттер алу үшін 3-5 мецилия қажет. Пенициллин мол көлемде керек болғандықтан оны шығару үшін 200000дм3 аймақтағы ферменттер құрылды. Күнделікті 500 ку – дай глюкоза қолданылад. Пеницилиннің жасалуы температураға бағынышты. Температура сағатына 2°С болуын қатаң қадағалайды. Сонымен қатар, рҺ – тың мәнін тексеріп, қадағалап тұрады.бұл жұмыс 15 тәулікке созылады. Одан арғы жұмыс мицелияны фильтрация жұмысының айыру жолымен басталады. Пенициллин ерітіндісін сұйық түрінде қалады. Әрбір экстракцияланған сайын жарты қосындысын алып отырады. Сөйтіп, пенициллин барған сайын тазара түседі. Әбден таусылғанша осы жол қайталана береді. Одан соң ваккумды кептіріп,барлығында судан тазартады. Ал пенициллин натрий немесе калий тұзы ретінде криссталданып қалады.
12.11.2. Моноклондық антиденелер
Адамдар бөтен молекулалардың құралуынан сақтанатын системалы болады. антидененің болуын қамтамасыз ету молекуланы антиген дейміз. Негізі бұлар уызды мен грикопротеиндардан тұрады. Әрбір антиген спецификалық антидененің құрылуын туғызады. Егер белгілі бір өзінің бактерияларымен бөтен дене жабыстыратын болса, иммундық система. Оның антигенін қарап танып, оған қарама – қарсы антидене жасалады. Антиденелер ерекше лимфоциттерден құралып, В – жасушалар деп аталады.
1970 жылы Цезарь Милыитейн және Георг Кёлер Кембриджда жұмыс жасап жүріп,м антиденелердің бір түрін жасап шығарды. Бұған дейін антиденелер жануарлардың қанынан алынып отырды. Милыитейн және Кёлер 1977 жылы Моноклондық антидене тауып, Нобель сыйлығын алды. Моноклондық деген бір клонға жатады. Әрбір антидене В1 – жасушамен бегіленеді. Олар өздерін клондап, басқа сөзбен айтқанда өздеріне ұқсастығын антигенді көбейтеді. Теориялы жағдайларда бегілі бір В жасушаларын өнімдеп, көптеп шығаруға болады. Оны моноклоналдық деп атайды. Бірақ дәнді дақылдар орталарында В жасушасы азғантай күн тұрақтай алады. Милыитейн мен Кёлер В жасушасын обыр жасушасымен араластырып, гибридома жасушасын алды. Бұл жасушалар көбейіп, клондалады және көптеген көлемде антидене алуға болады.
Моноклонды антиденелерін алу
Моноклонды антиденелерін алу үшін белгілі бір антигенді алып, тышқанға егеді. Белгілі бір уақыттан кейін жануардың өтінен лимфоцит бөлінеді. Оларды обыр ауруларының ерекше түрімен араластырғаннан лиэтиленгликоль ортасынан қосады. Бұл жерде тек гибрид жасушасы өсе алатын жағдайларды алады. Бұлардың өздерінің ішінен антидене бөлетін жасушасын бөлектеп алады. Бұл жасушалар бөлінеді, және ылғи таза антидене көзі болып табылады.
Моноклондық антидене қолдану
Моноклондық антидене белгілі бір антигенге арналға. Оларды антигенді табу үшін немесе жою үшін пайдаланады. Моноклондық антиденелер клиникалық лабароторияда кеңінен пайдаланады. Әзірше олар диагностика мақсатында қолданылады.
Жүктілік тесті
150 диагностикалық моноклондық антиденеден жүктілік кезін тексеру үшін пайдаланнылады. Жатырдағы эмбрион 4 күн болғаннан бастап, ол жатырдың қабығына орналасады. Бұл процесс имплантация деп аталады. Имплантация әйелдің жұмыртқасынан тексеріледі. Мұндай жағдайда жұмытқалар жатырдың ішкі қабатындағы жағдайларды жөніненгормон бөледі. Сол кезде етеккір тоқтайды, ал эмбрион сақталады. Жұмытқаға харион түтікшелері бар гормон сигнал береді. Бұл гормондардың аты хариондық гонадотропин. Ол ананың қанында болып, жұмыртқаға түседі. Әйелдің етеккірі қалыпты жағдайда болғанда ХГЧ (қаннық құрамы) мөлшерден жоғары көтереді. Оны зәрден біледі. ХГЧ глюкопротеин болып саналады. Оны жоғарыдағы антидененің көмегімен алады. Заманауи жүктілік тексеру тесті моноклондық антидене арқылы жүреді. 12.17 суретке қараңыз.
5 минутқа үй жағдайында тексеруге болатын тест дәріханаларда сатылады.
Ауру диагностикасы
Ауру диагностикасы жыныс жолымен жұғатын кең таралған ауру хламидиоз. Бұл ауруларды туғызатын грамотрицательді бактериялар хламадиоздың ерігіштігі, ол жасуша ішіндег паразит. Оның симптомдарын білу өте қиын. Сондықтн оны көбіне гонореямен шатастырады. Бұл 2 ауруда әйелдің жатыры арқылы өтіп, фаллопиевті трубкаларын залалсыздандырады. Бұл ауру түрлі ауруларға әкеліп, бедеулікке соқтырады. Моноклоналдық антидене табылғанға дейін, хламодиозды табу өте қиын болып табылады. Антидене табылғаннан кейін ауруларды емдеу тез әрі үмітті болады.
Ауруду табу жылдамдығы 15 -20минуттай болады.
Моноклоналдық антиденелердің тағы бір түрі стрептококтық диагностикасына қолданылады. Бұндай құралдың арқасында ауруды тез тауып,ем жасайды.
Қазіргі кезде моноклоналдық антиденелер вирус герпесінің 1 –ші типін тауып отыр. 10 -12 % генитальдық герпестің 1 –ш типтегі топтардан болады.әрбір инфекцияның емі әртүрлі болғандықтан,олардың вирустардан дәл айырып табу маңызды. Сонымен бірге кейбір обыр ауруларының алдын алу мақсаттарда, мысалы, өкпе ауруы, кеуде обыры, тікішек және тоқішек обырларын емдеу үшін әдістер ойлап табылуда. Адамның денесінде обырларды емдеу әдістері ойластырылуда. Мысалы, йод В1жатады. Антиденелер денеге сіңгесін денедегі ауру жасушаны іздеп табады. Бұл әдістер обыр жасушасын жою үшін пайдалынылады.
Фермент көбіне тат баспайтын темірден цилиндр түрінде жатады.
Шартты белгілер:
- Ауа шыңатын тесік. Көп жағдайда ферменттің аэробты болатындықтан ауаны тазалу үшін б.рку әдісін пайдаланып, көпіршіктер зат құрамын араластырып, оттегінің кіруін қамтамасыз етеді.
- Фермент көбіне айналдырғыш болады. Айналдыру жылдамдығы арасында оттегі өнімдерінің жақсы араласып,жасушлардың бір –біріне жабысуынан соңғы және ортада жылу алмасу жүреді.
- Айналу қалақтарында көбіне тіке, вертикалды орналасады.
- У көпіршікті басу үшін кіргізгіш түтіктер. Олар бақылау датчиктерімен жалғанған. Аэрация мен араластыру барысынд акөпіршік пайда болады. бұлар өнімнің кедергінің тесіктен шығуына кедергі жасайды. Сол кезде кпіршік басатын датчикпасы рольін ойнайды.
- Разетка ферменттер құрамы қысымда болады. сондықтан олар монометрмен қауіпсіз клапанына қосылады.
- Ферменттердің жоғарыдағы жағында тесік болады. ферменттерді тазалау соның жұмысы.
- Сөндіргіш бұл вертикалды қабырға, ферменттің ішкі қабырғасына орналастырады, бұл араластыруды көшейтін, өнімнің оттегін алуына тиімділігін тигізеді.
- Салқындатқыш құрал температура түсіреді. Сол кезде өнімнің өсуінің арқасында өздігінен жылу бөлінеді.
- Ферментация кезіндегі жұмыстан разетка арқылы бақылау үшін өнімнен үлгі алады.
Ферментация болғаннан кейін, өнімді ортадан шығарылу тазалау керек. Негізінде, ферменттердің құрал алдыңғы бөлшектер, сұйық және қатты жасушалары бар бөлшектерді бөледі. Бұл фильтрациямен центрифугация көмегімен жүреді. Керекті өнім сұйық түрінде болады не жасуша ішінде болады. тазалағанның биохимиялық тазалау неше түрі болады: бұған, кептіру, хромотография, еріткіш экстракциямен дестициялау, ары қарай өңдеу жұмысын инсулин шығатын завод 90% - ы 200 адамнан құралған жұмысымен жасайды. Пенициллинмен микропротеин өңдеу жұмысын жасалады. 12.11.1 және 12.12.3 суреттерінде.
12.4. Кесте. Үздіксіз және периодтық өңдеудің пайдасы мен зияны.
12.11.1. Пеницилмен шығарылуы
Пенициллин антибиотиктерін шығару үздіксіз шығару жақсы үлгіс.
Антибиотик дегеніміз негізгі
- Антибиотик
- Үздіксіз өнімдеу арқылы жасалған субтрат
- Екінші метоболит? Дәнді дақылдан субстрат. Екінші метоболиттерді алу үшін
Пенициллин саңырауқұлағын дерттеу барысында, алғашқы болып АҚШ тың Дүниежүзілік соғыстарында қолданбақ болды. Соған қарамастан Флеминг пенициллинді Англияда ашты. Бұл ізденіс Оксфордуниверситетінің сэрі Говардом Флори қолдауымен болды. Бұл жерде өнімдердің неше түрлері үлгідегі сапасы тексерілдң. Бірақ, пенициллин жоғарғы орын алды. Содан бері қарай жоғарғы өнімді мутаттық штамптарда, сонымен бірге, өнімді өсіру, оларды бөліп шығару және тазалағанда өнімі 2000 есе өсті.ортаның шешуші мәнін ферментация жұмысы шешті. Керек көміртегімен глюкоза бұларға, ең керек көзі болып табылады. Глюкоза саңырауқұлақтың өсуін қамтамасыз етеді. Бірақ, ферменттін өнуін басады. Сондықтан, алдымен 30 – 40 сағ саңырауқұлақтың тез өсуін қамтамасыз етеді. Ал содан сң глюкоза ең төменгі концентрацияда бақылай отырып қосады. Азот көзі болып, арзан ақуыз алынады. Мысалы,соялық және балық ұны. Сонымен бірге, фенилуксустық қышқыл қажет. Пеницилин екіншіші метаболит болғандығтан ол бірінші тез өсетін фазадан кейін қосады. Бірінші метаболитке демалу өнімдері көмірмен этиль спирті жатады. Өсетін өнімді саңырауқұлақ жиналғаннан алады. 50000дм3 аймақтағы иникулияция ферменттер алу үшін 3-5 мецилия қажет. Пенициллин мол көлемде керек болғандықтан оны шығару үшін 200000дм3 аймақтағы ферменттер құрылды. Күнделікті 500 ку – дай глюкоза қолданылад. Пеницилиннің жасалуы температураға бағынышты. Температура сағатына 2°С болуын қатаң қадағалайды. Сонымен қатар, рҺ – тың мәнін тексеріп, қадағалап тұрады.бұл жұмыс 15 тәулікке созылады. Одан арғы жұмыс мицелияны фильтрация жұмысының айыру жолымен басталады. Пенициллин ерітіндісін сұйық түрінде қалады. Әрбір экстракцияланған сайын жарты қосындысын алып отырады. Сөйтіп, пенициллин барған сайын тазара түседі. Әбден таусылғанша осы жол қайталана береді. Одан соң ваккумды кептіріп,барлығында судан тазартады. Ал пенициллин натрий немесе калий тұзы ретінде криссталданып қалады.
12.11.2. Моноклондық антиденелер
Адамдар бөтен молекулалардың құралуынан сақтанатын системалы болады. антидененің болуын қамтамасыз ету молекуланы антиген дейміз. Негізі бұлар уызды мен грикопротеиндардан тұрады. Әрбір антиген спецификалық антидененің құрылуын туғызады. Егер белгілі бір өзінің бактерияларымен бөтен дене жабыстыратын болса, иммундық система. Оның антигенін қарап танып, оған қарама – қарсы антидене жасалады. Антиденелер ерекше лимфоциттерден құралып, В – жасушалар деп аталады.
1970 жылы Цезарь Милыитейн және Георг Кёлер Кембриджда жұмыс жасап жүріп,м антиденелердің бір түрін жасап шығарды. Бұған дейін антиденелер жануарлардың қанынан алынып отырды. Милыитейн және Кёлер 1977 жылы Моноклондық антидене тауып, Нобель сыйлығын алды. Моноклондық деген бір клонға жатады. Әрбір антидене В1 – жасушамен бегіленеді. Олар өздерін клондап, басқа сөзбен айтқанда өздеріне ұқсастығын антигенді көбейтеді. Теориялы жағдайларда бегілі бір В жасушаларын өнімдеп, көптеп шығаруға болады. Оны моноклоналдық деп атайды. Бірақ дәнді дақылдар орталарында В жасушасы азғантай күн тұрақтай алады. Милыитейн мен Кёлер В жасушасын обыр жасушасымен араластырып, гибридома жасушасын алды. Бұл жасушалар көбейіп, клондалады және көптеген көлемде антидене алуға болады.
Моноклонды антиденелерін алу
Моноклонды антиденелерін алу үшін белгілі бір антигенді алып, тышқанға егеді. Белгілі бір уақыттан кейін жануардың өтінен лимфоцит бөлінеді. Оларды обыр ауруларының ерекше түрімен араластырғаннан лиэтиленгликоль ортасынан қосады. Бұл жерде тек гибрид жасушасы өсе алатын жағдайларды алады. Бұлардың өздерінің ішінен антидене бөлетін жасушасын бөлектеп алады. Бұл жасушалар бөлінеді, және ылғи таза антидене көзі болып табылады.
Моноклондық антидене қолдану
Моноклондық антидене белгілі бір антигенге арналға. Оларды антигенді табу үшін немесе жою үшін пайдаланады. Моноклондық антиденелер клиникалық лабароторияда кеңінен пайдаланады. Әзірше олар диагностика мақсатында қолданылады.
Жүктілік тесті
150 диагностикалық моноклондық антиденеден жүктілік кезін тексеру үшін пайдаланнылады. Жатырдағы эмбрион 4 күн болғаннан бастап, ол жатырдың қабығына орналасады. Бұл процесс имплантация деп аталады. Имплантация әйелдің жұмыртқасынан тексеріледі. Мұндай жағдайда жұмытқалар жатырдың ішкі қабатындағы жағдайларды жөніненгормон бөледі. Сол кезде етеккір тоқтайды, ал эмбрион сақталады. Жұмытқаға харион түтікшелері бар гормон сигнал береді. Бұл гормондардың аты хариондық гонадотропин. Ол ананың қанында болып, жұмыртқаға түседі. Әйелдің етеккірі қалыпты жағдайда болғанда ХГЧ (қаннық құрамы) мөлшерден жоғары көтереді. Оны зәрден біледі. ХГЧ глюкопротеин болып саналады. Оны жоғарыдағы антидененің көмегімен алады. Заманауи жүктілік тексеру тесті моноклондық антидене арқылы жүреді. 12.17 суретке қараңыз.
5 минутқа үй жағдайында тексеруге болатын тест дәріханаларда сатылады.
Ауру диагностикасы
Ауру диагностикасы жыныс жолымен жұғатын кең таралған ауру хламидиоз. Бұл ауруларды туғызатын грамотрицательді бактериялар хламадиоздың ерігіштігі, ол жасуша ішіндег паразит. Оның симптомдарын білу өте қиын. Сондықтн оны көбіне гонореямен шатастырады. Бұл 2 ауруда әйелдің жатыры арқылы өтіп, фаллопиевті трубкаларын залалсыздандырады. Бұл ауру түрлі ауруларға әкеліп, бедеулікке соқтырады. Моноклоналдық антидене табылғанға дейін, хламодиозды табу өте қиын болып табылады. Антидене табылғаннан кейін ауруларды емдеу тез әрі үмітті болады.
Ауруду табу жылдамдығы 15 -20минуттай болады.
Моноклоналдық антиденелердің тағы бір түрі стрептококтық диагностикасына қолданылады. Бұндай құралдың арқасында ауруды тез тауып,ем жасайды.
Қазіргі кезде моноклоналдық антиденелер вирус герпесінің 1 –ші типін тауып отыр. 10 -12 % генитальдық герпестің 1 –ш типтегі топтардан болады.әрбір инфекцияның емі әртүрлі болғандықтан,олардың вирустардан дәл айырып табу маңызды. Сонымен бірге кейбір обыр ауруларының алдын алу мақсаттарда, мысалы, өкпе ауруы, кеуде обыры, тікішек және тоқішек обырларын емдеу үшін әдістер ойлап табылуда. Адамның денесінде обырларды емдеу әдістері ойластырылуда. Мысалы, йод В1жатады. Антиденелер денеге сіңгесін денедегі ауру жасушаны іздеп табады. Бұл әдістер обыр жасушасын жою үшін пайдалынылады.
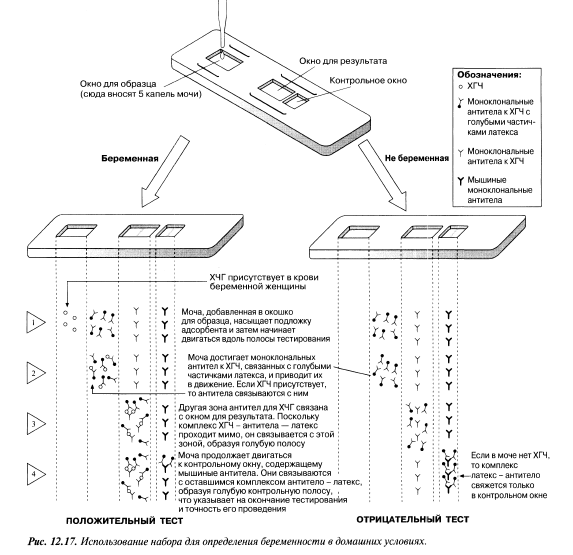
Аурудың емі – сиқырлы оқ
Сиқырлы оқтар - аурулардың емі. Бұл жай ғана олардың диагностикасы үшін емес , ұзақ уақыт бойы, моноклоналды антиденелер ауруларды емдеу үшін пайдалануға болады деген үміт бар. Арнайы моноклоналды антиденелер мақсатты табу және олармен байланыстыруға қабілеті бар,мысалы, ісік жасушалары сияқты, мұндай антиденелер «сиқырлы оқтар» ретінде белгілі болды. Егер антиденелерге улы химиялық зат немесе радиоактивті изотопы байланысса, онда олар жасушаны жояды деген үміт бар. Тағы Бір мәселе, антигендер ісік жасушаларына тән, сондай-ақ олар белгілі бір қалыпты жасушаларының,яғни олар да құритынына міндетті екенін ашқан. Тағы басқа бір проблема, антидене обыр жасушаның беткі жағымен байланысады, яғни оның ішіне енбейді, себебі антидене межелі жерге уыттар жеткізу керек. Дегенмен, бұл салада бұл кейбір прогресс болды. Көшірілген органдардың қабылдамауынан сақтап қалу Трансплантацияның ең маңызды проблемалардың бірі, ол организмнің иммундық жүйесі бөтен сияқты жаңа органды танып қоюуында және оған қарсы шабуыл жасауында. Бұл проблемалардың шешуінің бір жолы, ол науқастың иммундық жүйесін жою болып табылады. Антидене көшірілген бүйректің қабылдамауын сақтап қалу үшін өте тиімді қызмет етеді. Бұндай антиденелер антигендермен әрекеттеседі, барлық Т жасушаларын анықтайды. Т-лимфоциттер - әдетте вирус жұқтырған немесе қатерлі жасушаларға шабуыл жасайтын лимфоциттің бір түрі. Бұл жасушалар көшірілген органдарды сақтап қалу үшін қатысады.. қолданылған иммундық жүйені басып тастау кезінде, Моноклонды антиденелер, кәдімгі дәрілерге қарағанда өте тиімді. Олар иммундық жүйенің барлығын емес, дәрі дәрмектер сияқы соның ішіндегі Т жасушаларын ғана басып тастай алады. Осылайша, науқаста ауруға қарсы күресі сақталады.
Трансплантациялау үшін тіндерге талдау жүргізу. Қабылдамаудың қауіп қатерлерін азайту үшін, органдардың бір біріне көшірілмес бұрын оларға сәйкес донор табу керек. Бұл дегеніміз, донордың антигендері науқастың антигендерімен сәйкес келуі қажет. Антигендердің сәйкес келмеуі, қауіа қатердің өсуіне әкеліп соқтырады. Донордың антигенінің шығу тегін қарастыру үшін, моноклоналдық антиденелер қолданылады.
12.11.3. Инсулин және адам бойын өсіретін гормон
Қазіргі таңда гендік инженериямен бұл өнімді бактериядан түрлендіретін әдіс арқылы алуға болады. гендік инженерияны биотехнологияның бір бөлігі ретінде қарастыруға болады, біз оны 25 бөлімде басқа да заттармен қалай қолданылатыны туралы мәселесін талқылаймыз. Және сол бөлімде инсулин және бой өсіру гормоны туралы айтылады.
12.11. Тамақ өнімдері және сусындар
Тамақ өнімдерін дайындалуы және сусындар сонау биотехнологияның атасымен байланысты болып келеді. Нан, ірімшік, йогурт, сірке су және сыра, шарап сияқты алькогольдық ішімдіктер олар 1000 жылдан бері пайда болып келе жатқан және тарихтан бұрынғы замандардан әйгілі. Бір бұрынғы өсиеттерде вино туралы ескертулер бар(римдіктер сыра мен шарап ішкен). Біздің заманымызға дейінгі 4000 жылдықтан бері көне египпеттіктер шарап қайнатқан және нанды көтеріп пісіру үшін ашытқылар қолданған. 10000 жылдан бері есептеуге болатын ой биотехнологияда жануарларды будандастыруға және өсімдіктерден тамақ өнімдерін алуға және ауыл шаруашылығы болады. Біз бұл тақырыпта тек ескі технологияларды қарастырмаймыз, тамақ өнімдерінің өндірілуінің жаңа әдістерін де қарастырамыз.
12.12.1. Ашытылған – нан, сыра және шарап
Ашытқылар – біржасушалы саңырауқұлақ. Ашытқылар табиғатта кеңінен жайылған және олардың дайындалу әдістерінің тестісі немесе сахардың пайда болған спитрпен ашытылуы туралы елестету қиын емес, бірақ процестің шын мәнінде түсініксіз түрде болып қалады. 19ғасырда тек қана Луи Пастер ашытқының мкроорганизмдердің белсенділігімен пайда болатынын көрсетті.
Сиқырлы оқтар - аурулардың емі. Бұл жай ғана олардың диагностикасы үшін емес , ұзақ уақыт бойы, моноклоналды антиденелер ауруларды емдеу үшін пайдалануға болады деген үміт бар. Арнайы моноклоналды антиденелер мақсатты табу және олармен байланыстыруға қабілеті бар,мысалы, ісік жасушалары сияқты, мұндай антиденелер «сиқырлы оқтар» ретінде белгілі болды. Егер антиденелерге улы химиялық зат немесе радиоактивті изотопы байланысса, онда олар жасушаны жояды деген үміт бар. Тағы Бір мәселе, антигендер ісік жасушаларына тән, сондай-ақ олар белгілі бір қалыпты жасушаларының,яғни олар да құритынына міндетті екенін ашқан. Тағы басқа бір проблема, антидене обыр жасушаның беткі жағымен байланысады, яғни оның ішіне енбейді, себебі антидене межелі жерге уыттар жеткізу керек. Дегенмен, бұл салада бұл кейбір прогресс болды. Көшірілген органдардың қабылдамауынан сақтап қалу Трансплантацияның ең маңызды проблемалардың бірі, ол организмнің иммундық жүйесі бөтен сияқты жаңа органды танып қоюуында және оған қарсы шабуыл жасауында. Бұл проблемалардың шешуінің бір жолы, ол науқастың иммундық жүйесін жою болып табылады. Антидене көшірілген бүйректің қабылдамауын сақтап қалу үшін өте тиімді қызмет етеді. Бұндай антиденелер антигендермен әрекеттеседі, барлық Т жасушаларын анықтайды. Т-лимфоциттер - әдетте вирус жұқтырған немесе қатерлі жасушаларға шабуыл жасайтын лимфоциттің бір түрі. Бұл жасушалар көшірілген органдарды сақтап қалу үшін қатысады.. қолданылған иммундық жүйені басып тастау кезінде, Моноклонды антиденелер, кәдімгі дәрілерге қарағанда өте тиімді. Олар иммундық жүйенің барлығын емес, дәрі дәрмектер сияқы соның ішіндегі Т жасушаларын ғана басып тастай алады. Осылайша, науқаста ауруға қарсы күресі сақталады.
Трансплантациялау үшін тіндерге талдау жүргізу. Қабылдамаудың қауіп қатерлерін азайту үшін, органдардың бір біріне көшірілмес бұрын оларға сәйкес донор табу керек. Бұл дегеніміз, донордың антигендері науқастың антигендерімен сәйкес келуі қажет. Антигендердің сәйкес келмеуі, қауіа қатердің өсуіне әкеліп соқтырады. Донордың антигенінің шығу тегін қарастыру үшін, моноклоналдық антиденелер қолданылады.
12.11.3. Инсулин және адам бойын өсіретін гормон
Қазіргі таңда гендік инженериямен бұл өнімді бактериядан түрлендіретін әдіс арқылы алуға болады. гендік инженерияны биотехнологияның бір бөлігі ретінде қарастыруға болады, біз оны 25 бөлімде басқа да заттармен қалай қолданылатыны туралы мәселесін талқылаймыз. Және сол бөлімде инсулин және бой өсіру гормоны туралы айтылады.
12.11. Тамақ өнімдері және сусындар
Тамақ өнімдерін дайындалуы және сусындар сонау биотехнологияның атасымен байланысты болып келеді. Нан, ірімшік, йогурт, сірке су және сыра, шарап сияқты алькогольдық ішімдіктер олар 1000 жылдан бері пайда болып келе жатқан және тарихтан бұрынғы замандардан әйгілі. Бір бұрынғы өсиеттерде вино туралы ескертулер бар(римдіктер сыра мен шарап ішкен). Біздің заманымызға дейінгі 4000 жылдықтан бері көне египпеттіктер шарап қайнатқан және нанды көтеріп пісіру үшін ашытқылар қолданған. 10000 жылдан бері есептеуге болатын ой биотехнологияда жануарларды будандастыруға және өсімдіктерден тамақ өнімдерін алуға және ауыл шаруашылығы болады. Біз бұл тақырыпта тек ескі технологияларды қарастырмаймыз, тамақ өнімдерінің өндірілуінің жаңа әдістерін де қарастырамыз.
12.12.1. Ашытылған – нан, сыра және шарап
Ашытқылар – біржасушалы саңырауқұлақ. Ашытқылар табиғатта кеңінен жайылған және олардың дайындалу әдістерінің тестісі немесе сахардың пайда болған спитрпен ашытылуы туралы елестету қиын емес, бірақ процестің шын мәнінде түсініксіз түрде болып қалады. 19ғасырда тек қана Луи Пастер ашытқының мкроорганизмдердің белсенділігімен пайда болатынын көрсетті.

Астық тұқымдастартар тұқымынан ұнтақтап нанды сол ұннан алады. Ұн – бұл крахмалдың ең негізгі түрі (дәннің ақ түрі), қоректік заттың қосалқысы болып табылады және көктеудің қалыпты жағдайында жұмсайды. Ферменттердің тұқымына ішінара кіретін мальтоза мен глюкоза крахмалды қантқа дейін сіңіріп алады. Қанттың маңызын арттыру үшін, саңырауқұлақтан алынған амилазаны крахмалды жаңқалау үшін де қосуға болады. ашытқылар қантты тыныс алудың энергия көзі ретінде пайдаланады. Нәтижесәнде, аэробты және анаэробты дем алу көмірқышқыл газы болып табылады. Газ көпіршіктерін көтеру үшін жылы тестіде ұстайды. Бұл кезең қамыр ашыту деп аталады. Ашытқы штаммаларының бөлінуі Saccharomycescerevisiae, олар өте көп көмірқышқыл газын құрайды. Анаэробты ашытқысы процесі кезінде спирт құралады, келесі ашытудан ол спирт пісіру процесінде буланады.
Сыра
Ферментация пайдаланғаннанбастап, өндірістің атасынан бері сыра қайнату пайда болды. Сыра күні бұрын өсетін арпа түйіршіктерінен қайнатылады, сыра қантқа айналу үшін, мысал,а мальтозаны алуға болады. Бұл процесс ашыту деп аталады. Оның жұмысын тездету үшін және де дұрыс қылу үшін гиббериллиндер қолданылады. Бұдан басқа пайда болатын секердің шамасын көбейту үшін көптеген ферменттерді қосады. Мысалы, амилаза. Бұл спирттің көбеюіне әкеліп соғады. Нәруыздардың жарылуынан сұйықтықта азот пайда болады.
Температураны жаймен 80°С ысытқанда дәндердің өсуін тоқтатады. Ашытқыны келесі кезеңдер бойынша дайындар кезінде одан төмен температурада ысытады. Сыраның иісі мен түсі оның ысыту температурасына байланысты. Содан кейін секерді бөліп алу үшін дәндерді ыссы суға малып, түбіне кіргізеді. Осыдан пайда болған сулы экстратқа немесе сыра ашытқысына ащы дәм беретін және бактерияның өсуін тоқтататын құлмақ қосады. Сыра ашытқысын қайнату және құлмақты алып тастау арқылы концентрацияланады, сол кезде ашытқы пайда болатын температураға дейін салқындатады. Периодттық өсу үшін сыра ашытқысын ферменттерге орналастырады және оларға алдыңғы ферментациямен қалған сыра ашытқыларын қосады. Көбінесе екітүрлі ашытқы қолданады: Saccharomycescerevisiaе және Saccharomycescarlsbergensis. Соңғысы одан жеңіл сыраларды өндіру үшін қолданылады. Ашыту анаэробты жағдайларда пайда болады. Нанды өндірудегі секілді бұл ашыту кезеңінде секер спиртке және көміртектің қостотығына айналады. 2 -5 тәуліктен кейін спирттің құрамы 3,5 - 8% (көбінесе3,5—5%)жетеді. Жеңіл сыраның түрлерін дәстүрлі өндіру кезінде қолданылатын ашытқылар әдетте ферментердің астыңғы бөлігінде өседі және төменгі ашытқылар деп аталады. Сол кезде жоғары алкогольді құрылымы бар берік сыраның түрін өндіру кезінде қолданатын жоғары ашытқылар жоғары көтеріледі. Бірақ қазіргі кезде берік сыра түрлерін өндіруде төменгі ашытқы болып табылатын жаңа сыралық ашытқылар қолданылады, олар сол үшін дәл сол ферментер екі түрлі сыраның түрін ала алмайды. Осы ашыту процесі өткеннен кейін ашытқылар сырадан бөлінеді.
Шарап
Ашытуға керек бірнеше күнге созылатын секер көзі жүзім болып табылады. Шараптың дәмі жүзім түрімен, сонымен қатар ашыту кезінде қолданылатын ашытқы штаммымен анықталады. Қызыл шарап өзінің түсін ашыту кезінде жоғалмайтын қабығымен алады. Ақ шарапты өндіру кезінде винофадты қабығын ереже бойынша лақтырады. Қазіргі таңда жабайыға қарағанда, сенімірек коммерциялық ашытқы штаммдарын пайдаланудыжөн көреді. Жиналған жүзімді езеді және осы «муст» пайда болған кезде ашыту пайда болады. Кейде шарап сүт қышқылы бактериялары қосылған екінші ашытуға тап келуі мүмкін. Бұл кезде алма қышқылы сүт қышқылына және көміртек қостытығына айналады. Бұл кезде шарап қышқылдығы азаяды.
Сыра
Ферментация пайдаланғаннанбастап, өндірістің атасынан бері сыра қайнату пайда болды. Сыра күні бұрын өсетін арпа түйіршіктерінен қайнатылады, сыра қантқа айналу үшін, мысал,а мальтозаны алуға болады. Бұл процесс ашыту деп аталады. Оның жұмысын тездету үшін және де дұрыс қылу үшін гиббериллиндер қолданылады. Бұдан басқа пайда болатын секердің шамасын көбейту үшін көптеген ферменттерді қосады. Мысалы, амилаза. Бұл спирттің көбеюіне әкеліп соғады. Нәруыздардың жарылуынан сұйықтықта азот пайда болады.
Температураны жаймен 80°С ысытқанда дәндердің өсуін тоқтатады. Ашытқыны келесі кезеңдер бойынша дайындар кезінде одан төмен температурада ысытады. Сыраның иісі мен түсі оның ысыту температурасына байланысты. Содан кейін секерді бөліп алу үшін дәндерді ыссы суға малып, түбіне кіргізеді. Осыдан пайда болған сулы экстратқа немесе сыра ашытқысына ащы дәм беретін және бактерияның өсуін тоқтататын құлмақ қосады. Сыра ашытқысын қайнату және құлмақты алып тастау арқылы концентрацияланады, сол кезде ашытқы пайда болатын температураға дейін салқындатады. Периодттық өсу үшін сыра ашытқысын ферменттерге орналастырады және оларға алдыңғы ферментациямен қалған сыра ашытқыларын қосады. Көбінесе екітүрлі ашытқы қолданады: Saccharomycescerevisiaе және Saccharomycescarlsbergensis. Соңғысы одан жеңіл сыраларды өндіру үшін қолданылады. Ашыту анаэробты жағдайларда пайда болады. Нанды өндірудегі секілді бұл ашыту кезеңінде секер спиртке және көміртектің қостотығына айналады. 2 -5 тәуліктен кейін спирттің құрамы 3,5 - 8% (көбінесе3,5—5%)жетеді. Жеңіл сыраның түрлерін дәстүрлі өндіру кезінде қолданылатын ашытқылар әдетте ферментердің астыңғы бөлігінде өседі және төменгі ашытқылар деп аталады. Сол кезде жоғары алкогольді құрылымы бар берік сыраның түрін өндіру кезінде қолданатын жоғары ашытқылар жоғары көтеріледі. Бірақ қазіргі кезде берік сыра түрлерін өндіруде төменгі ашытқы болып табылатын жаңа сыралық ашытқылар қолданылады, олар сол үшін дәл сол ферментер екі түрлі сыраның түрін ала алмайды. Осы ашыту процесі өткеннен кейін ашытқылар сырадан бөлінеді.
Шарап
Ашытуға керек бірнеше күнге созылатын секер көзі жүзім болып табылады. Шараптың дәмі жүзім түрімен, сонымен қатар ашыту кезінде қолданылатын ашытқы штаммымен анықталады. Қызыл шарап өзінің түсін ашыту кезінде жоғалмайтын қабығымен алады. Ақ шарапты өндіру кезінде винофадты қабығын ереже бойынша лақтырады. Қазіргі таңда жабайыға қарағанда, сенімірек коммерциялық ашытқы штаммдарын пайдаланудыжөн көреді. Жиналған жүзімді езеді және осы «муст» пайда болған кезде ашыту пайда болады. Кейде шарап сүт қышқылы бактериялары қосылған екінші ашытуға тап келуі мүмкін. Бұл кезде алма қышқылы сүт қышқылына және көміртек қостытығына айналады. Бұл кезде шарап қышқылдығы азаяды.

Ашытқыш экстраты
Ашытқыштар сыра шығарылыдары әр түрлі жолмен алынады. Белгілі бір, мысал ретінде вискиденашытқыштың алынуы. Мұнда да сырадағыдай секермен ашытып барып, арпа тұқымымен ашытып алынады. Бірақ вискиді уыттан тазартылынып алады. Бұл жерде қайнату керек болады, ал қайнатылған кезде ашытқылар өледі. Олардың ортаны татыру үшін сыра қалдықтарынан қалған ашытуды пайдалынады. Ашытқыш жас дәрумендердің В тобына бай. Соның ішінде ниацин (В6), рибофлофин (В1), тиамин (В), рибофламин (В1) фолио қыщқылы және В12 дәрумені.
Ашытқыларды құруға және одан дәрумендер таблеткаларын жасауға болады. сонымен бірге мармит жасауға болады. ол үшін ашытқышты 50 °С қыздырады. Содан соң үлкен көлемде тұз қосады. 5 .12.2. Сүтқышқылды ашыту – сүт өнімдері
Көптеген сүт текті өнімдер деп аталатын өнімдерді сүт ашыту жолымен алуға болады. Сүтте анаэробты тыныс алу процесі кезінде лактозаны(сүтсекер) сүт қышқылына дейін айналдыратын сүт қышқылдары болады. Пастерлеу процесінде бұл бактериялар егер ашыту тағы керек болып оларды тағы сүтке қосқанда өлуі мүмкін. Ашыту жағдайына тәуелді сүт құрамы және көзі болатын қоспаларды қолданғанда әртүрлі өнімдер пайда болады. өнімдердің консистенциясы мен дәмінің айырмашылықтары бар. Оларды өзіне ірімшік, иогурт, сүзбе, май, қаймақ түрлерін қосып алатын спекторлары арқылы көруге болады.
Иогурт
Ірімшікке қарағанда өндіруге қолданылатын белгілі сүт фракциясына қарағанда иогуртты дайындау кезніде қаймағы алынбаған сүтті ашытады. Басында иогут сұйық және қышқылды болып жасалатын бірақ, Батыс елдерінде оның дәмі мен консистенциясын ұлттық дәстүрге қарай өзгертті. Себебі иогуттың өзі қышқылды болып табылады(pH 3,7—4,3). Оның дәмін жақсарту үшін жеміс - жидектер қосады.
Иогуртты дайындау үшін сүттің әртүрлі түрлерін: бөлек – бөлек немесе сұйықтықта сонымен қатар қаймағы алынбаған, алынған, жартылай алынған, қайнатылған және құрғақ түрлерін қолдануға болады. әр комбинация иогурттың ерекше түрін береді. Ең көп таралғаны төменгі май құрамы бар сапа. Бұл процесс схемасы 12.20 суретінде келтірілген.
Ұлыбританияда барлық сүт пастерлеуіне ұшырайды. Әдетте ол 72°С – та 15с- да жүргізіледі. Бұл патогенді микрооргнаизмдерден құтылу үшін жасалады. Олар міндетті түрде сүтте болуы тиіс емес, бірақ сүт микроағзалардың өсуі үшін тамаша ортасы болып табылады, өйткені, әрқашан зиянды бактерия, мысалы Salmonella сияқты бактериялар, немесе саңырауқұлақтар ластанған болуы мүмкін. Осындай бактериялардан бөлек кейбір бактериялар өз – өзіне зиянды емес және олар сүтте көбейіп оның сапасына әсерін тигізуі мүмкін. Себептерінің қатары 12.20 суретінде көрсетілген, иогурт дайындауындағы қажетті термиялық өңдеу. Сүт қышқылдарын алмастыру үшін, қыздыру алдында жоғплғандар, «әуелгі дақылдарға» қосылады. Сүт қышқылы және спирт, йогуртқа тән дәм береді, және Streptococcus thermophilus өндіретін, ерекше крем дәм қосады. Әуелгі дақылдар әдетте Lactobacillus Bulgaricus қамтиды. Бұл бактериялар йогурт Грам-боялған үлгідегі (бөлім 12.9.2) көруге қатынасы шамамен тең болуы мүмкін. Бактериялар екі түрі де Грам бояуымен боялған, сондықтан қызылға айналады. Сіз бактериялардың санын санау үшін тырыссаңыз, осы түрлердің әрқайсысы үшін бір грамм шамамен 108 жасушаларын таба алады.Бактериялардан жинақталған дақыл бақыланбалы және
болжамдалатын қаймағы алынбаған сүтті ашытқылайды. Бастапқыда, йогурт көп сұйқылтым және қышқыл болады, бірақ содан кейін оның дәмі мен құрылымы батыс елдерінде айтарлықтай ұлттық дәстүрлерге сәйкес өзгерді. Иогурт қышқылтым (pH 3,7—4,3)болғандықтан, оған оның дәмін өзгерту үшін әртүрлі жеміс – жидектерді қосады.
Ірімшік
Әуелі Ірімшік сүт өндіруде сүзбе және сарысуға бөлінеді. Осы мақсатта сүтке мұқият бақыланбалы керекті микрооргнаизмдер тазартылған жағдайларда әуелгі дақылға енгізіленеді. Сүтте сақталатын казеин ұюы арқылы процессті құрағыш сүт қышқылы сүтті ашытуды бастайды. Бұл кезде сүттің «ұюы»болып жатады. Құрамында реннин бар іріткі ферментінің қосылуы, коагуляция жеделдету және одан әсіресе оны бақылауға мүмкіндік береді. Әдетте, іріткі бұзау асқазанынан алынады, бірақ протеиназдың өзіне лайықты қасиеттерін саңырауқұлақтардың түрлерінен, бактерия штаммдарынан, гендік инженериясының модификсириялық әдістерінен айыруға болады. Осы жаңа ферменттердің артықшылығы кошерлық және вегетариандық ірімшік дайындау үшін оларды пайдалану мүмкіндігі болады.
Сүзбелік массаны бөледі, сығады, қажетті пішінді беріп және кептіріледі. Сарысу өз кезегінде, өңделгеннен кейін оны малға азық ретінде бергеннен соң оны қорек көзі ретінде ашытқылардың өсуі ретінде қолданады. Өнімде ақуыз және дәрумендердің көп мөлшері бар.
Ірімшіктің дәм беруі және процесс барысында пісіп жетілуі оған әртүрлі микрооргнаизмдер қосады: бактериялар (сырдың бір түрі), өңезденген саңырауқұлақ (қой сүтінен жасалған ірімшіктің бір түрі және тағы басқа да көк ірімшіктер) немесе басқа тәсілдері. Сапасы әртүрлі ірімшіктердің әртүрлі дайындалуындағы барлық процесстердер кезінде бақыланбалы жағдайлар жасалынылады. Кейбір ірімшіктер, шикі күйінде ауылдық және кілегей түрінде қала береді.
Май
Май ең кілегейден жасалады, жаңа сүттің қосындысынан жасалады, және де заң бойынша ол 80%майдан төмен болмау керек. Ең бірінші, кілегейді пастерлейді, содан соң шайқайды.
12.12.3. Біржасушалы уыздар
Жаңа қоректік көзі болып табылатын бір жасушалы уыздар. Оның шығарылымы 1960 жылы басталады. Ірі масштабтағы микрооргнаизмдерөсіретін мысалы, балдырлар ашытқыштар, саңырауқұлақтар тобы уыз терминіне жатады. Уыз адам пайдасына және жануарлардың жемі болып саналады. Онда көптеген витаминдер көзі, дәрумендер, көміртегілер мен қажетті майлар. Теориялық жағынан бұл уыз тағамдарды ұн, бидай сияқтылардың орнын басады. Бірақ батыста ауыл шаруашылықтарының шикізаты барынша жеткілікті болуы мүмкін ел. Әлемдік масштаб бойынша, уыздардың жетіспеуі байқалады. Себебі, бұл елдерде биотехнологияның жұмысы дамымаған. Микрооргазимдердің қорек көзі ретінде пайдалануы төменде рет – ретімен көрсетілген:
Прутин
1980 жылы ІСІ компаниясы жұмысқа үлкен 1500м3, биіктігі 60 метр ферменттерді қосты.бұл Англияның солтүстік -батысындағы Билленгем қаласында болды. Оның құрамы фирмаға 40 млн фунтқа шықты. Ал жылдық 1 жасушалы уызын 70000 т шығарды. Бұл жерде дәндә дақылдар Methylophilusmethylotrophus бактерияны пайдаланды, ол оттегі мен қуат күшінің көзі болды және арзан әрі тез табылатын метанол табылды. Жұмыс аэробты жағдайларында жасалды. Бұл жерде шикізаттар қолданылды.
Органикалық оттегі
(метанол)++ Азот + —► + Н20 + минералды тұздар + жылулық+қуат + көміртегі
Қос тотықты көміртегіні балондарға толтырып, сатты. Мұндай бактерияны келісіп отырып, ең жоғарғы жылдамдығы өзі арзан көміртегімен қуат көзін , сонымен қатар термотұрақтысы болу керек . себебі, ферментация кезінде жылу бөлінеді және бір құрғақ массаға өте жоғары құнардағы уыз болды.
Methylotrophus бактериясы уақыт генерациясы болды. Ол 2 – 5 сағатқа дейін уһұстайтын болды. Азотты амони тұзы түрінде қосты. Сонымен бірге минералды қоспалар, фосфорлар және калий қолдануда.
Метанолды Биллингемнің табиғи газынан алды. Сонымен қатар, ол газ қосымша зиянды заттарды құрамады. Температураны қатаң қадағалап отырды. Ол 30—40 °С рН – ты құрады. Ферментер бірнеше ай бойы тоқтамай жұмыс жасап тұра алды.
Прутиннің шығарылымы дақылды өсірудің классикалық түрі болды. Оның негізі суреті 12.10.5 суретінде көрсетілген. Сондықтан оны, аэрациялау мен салқындатылуы жеңіл болды. Ол ауа берудің ең көрнекті жолы болып табылады. Ферментер түбіне қысымдалған ауа, көпіршіктер көтеріле кем жұмыс жасады. Жылына 6000т уыз алынды. 2000 тонна метанолдан 1 т кептірілген прутина алды. Оның құрамында 72% уыз және 8% ылғал болды. Жіне ең керекті аминқышқылмен, дәрумендер мен байылып, 2 есе қоректі болып шығарлды. Прутинді жануарлардың жемі ретінде пайдаланды. Дәнді дақылдардың жұмысы үздіңсіз юолғаннан кейін, бактерияларды ылғи жинап отырды. Жартысын ферменттерден шығарды, реагентпен тазалады. Шыққан жасушаларды желдетіп, кептірді. Ал қоректі сұйықтады ферменттерге қайтадан қайтарылды. Кептіріп болғаннан кейін бактериялық жасушаларды үгітеді. Себебі, қорауһптау алдында минералды құрамы мен рН-ы қадағалануы керек.
Микопротеин
Бір жасушалы ақуыздың Тағы бір әлеуетті көзі – саңырауқұлақтар.
Ашытқы сондай-ақ бактериялар, адам мен жануарлар пайдалануға болады (төменде қараңыз. Sec. «Ашытқы сіріндісі»). Қолданыс табады және өңезденген саңырауқұлақ. Мицелия дегеніміз - жіп – жіңішке гиф жиынтығынан тұрады, оларға типтік саңырауқұлақтық тән. Корам саңырауқұлақтарды қолдану үлгісі - микропротенді шығару. 1985 жылы Рэнк, Ховис және Макдоугал Marlowfoods компаниясымен біріге отырып, «Quorn» атты маркалы микропротеин шығарды. Осы мақсатта Fusariumgraminerium атты санырауқулақ пайдаланды.
Ол алғаш рет 1960 жылдары Марл оу топырақ үлгісі Бакингемширеге жақын шықты , осыдан компанияның атауы шықты. Ақуыз өнімін зерттеу 1964 жылдан басталды , ал адамдар үшін олардын қолданылыуын жане қауіпсіздігін 1986 алды . Басында кворнды хош иісті бәліштерді дайындау үшін қолданды, оны Дж. Сэйнсбери сатты.
Бірақ ол бәліш мен карри сияқты азық – түлік өнімдерін өндіру үшін қолданды, 1990 жылы сатылымға үйде пайдалануға кворн ұсақ тілімдермен келді, Ал 1992 жылы оны ұнтақ түрде сата бастады. Кворн сатып алушылар арасында жұмсак дәм жане хош иісімен табыс алды.
Саңырауқұлақты дәнді дақылдаумен өсіру уақыты 5,5 сағ құрайды. Демек, ол бактерияға қарағанда жай өседі. Көміртегі көзі және қуат көзі ретінде саңырауқұлағындағы глюкозаны қолданады. Оны кез келген арзан крахмал құрайтын өнімнен жүгеріден, бидайдан, күріштен, картоптан және млессадан. Саңырауқұлақ 1 кг қантқа 0,5 кг құрғақ биомассаны құрайды. Санырауқұлақтардың артықшылығы олар рН қышқыл жерлерде өсе алады. Бұл кезде бактериялардың өсуі тоқтатылады. Сөйтіп, дәнді дақылдың бұзылуына әкеліп соқтырады. Fusarium 30 градус жерлерде өседі. Бактерияларды өсіру секілді, азоттың көзі аммони тұздары болып табылады , және де олардың өсуіне минералды тұздарды олардың өсу ортасына қосады. Жоғарыдағы барлық үдерістердегідей, аэрация және суық орта қажет, ал қолданылатын барлық материалдар тазартылған болуы қажет. Аэрациялау механизмі арқылы ортаны араластыру жұмысы жүргізіледі. Бұл жұмыстарда механикалық қалақшаларлың жұмысы қажеті жоқ. Өйткені, қалақшалар тепе – тең ауданының жұмысын бұзады. Бұлардың жұмыс жасау механизмі ауа лифтісі деп аталады. Себебі, өнімдер үнемі қозғалыста болады. әрбір 2 мин сайын ауа көмегімен 1 айналым жасайлы. Ферменттердің аумағы 40000 дм 3 – ті құрайды. Осы ферменттердің үздіксіз жұмысына ылғи таңдалған өнімдермен жаңа орталар өажет.
Прокариотикалық жасушаларға қарағанда эукариоттық жасушалар өте жоғары нуклеин қышқылын құрайды. Сондықтан микропротеиндар аса маңызды көлемде нклеин қышқылының 5 - 15% құрғақ массасын құрайды. Бұл дегеніміз алдындағы мақсатта РНҚ оның құрамын азайту керек. Себебі, адам оны 2 күннен артық қолданбауы қажет.бқл дегеніміз бөтекеде тас олуы және подогра ауруына әкеліп соқтыруы мүмкін.. РНҚ дақылын алып тастау үшін 64 °Сқа дейін 20 – 30 мм арнайы дыс ішінде ұстап, қайнатылған бу жібереді. Қыздыру барысында протеиназдар белсенділігі төмендейді. Соөйтіп, ақуыз бұзылмайды. Бірақ , бұл РНҚ – ға қатысы жоқ. РНҚ құрамы бүкіләлемдік денсаулық орталығының жарлығы бойынша ең төмен 2 % көрсетілсе, бұл 1% - ға дейін төмендейді.
Мицелий саңырауқұлағын дәнді дақылдар орталығына бөлуге, бактерия жасушасын бөлуге қарағанда оңайырақ. Бұларды центрифугациялау қажет емес. Мицелийді жай сүзіге болады. сүзіп және кептіруден кейін, ол жұқа пленка тамыры болып табылады. Бұл сыртқы түріне қарағанда, шикі қамырды елестейді. Оған көкөніс қосқыштарын және аздаған жұмыс уызын қосады. Содан соң бөлшектетіп кеседі немесе ұсақтап сол түрінде сатылымға шығарады. Саңырауқұлақ табиғатынан етке ұқсайды. Сондықтан оған еттің дімңн оңай қосыға болады. адмның қолдануы жайында әңгімелегенде оның экономикалық факторларынан басқа көптеген жақтары қаралады. Мысалы, қолдану сақтылығын қатаң қадағалау. Заттың қорек санағы жоғары болуы. Осылардың бәрін 10 жылдай зерттеп, зиянды жолын таппады.
Тексерілу кем дегенде 4 ғасыр бері қарай 11 түрлі жануарларда, соның ішінде тышқан, шошқа мен сиырларды тәжірибе жүргізілді. Микропротеин шықпас бұрын өз еркімен келіскен адамдарды тексерілу жүргізілді. Алынған нәтижеде өнім адам денсаулығына еттен көрі пайдасы бар екенін көрсетті.
12.5. кесте қолданатын кейбір типті уыздармен майлардың құрамы көрсетілген. Бұлар холестеролды құрайды. Және майдың коллориясы төмен және бір жақсы қасиеті дәрімен көзі, В12 және цинк қоспасының
12.11. Кворнның проценті жарнамада табиғи өсімдік түйіршігі деп жазылады. Неге олай жазылған?
Ашытқыштар сыра шығарылыдары әр түрлі жолмен алынады. Белгілі бір, мысал ретінде вискиденашытқыштың алынуы. Мұнда да сырадағыдай секермен ашытып барып, арпа тұқымымен ашытып алынады. Бірақ вискиді уыттан тазартылынып алады. Бұл жерде қайнату керек болады, ал қайнатылған кезде ашытқылар өледі. Олардың ортаны татыру үшін сыра қалдықтарынан қалған ашытуды пайдалынады. Ашытқыш жас дәрумендердің В тобына бай. Соның ішінде ниацин (В6), рибофлофин (В1), тиамин (В), рибофламин (В1) фолио қыщқылы және В12 дәрумені.
Ашытқыларды құруға және одан дәрумендер таблеткаларын жасауға болады. сонымен бірге мармит жасауға болады. ол үшін ашытқышты 50 °С қыздырады. Содан соң үлкен көлемде тұз қосады. 5 .12.2. Сүтқышқылды ашыту – сүт өнімдері
Көптеген сүт текті өнімдер деп аталатын өнімдерді сүт ашыту жолымен алуға болады. Сүтте анаэробты тыныс алу процесі кезінде лактозаны(сүтсекер) сүт қышқылына дейін айналдыратын сүт қышқылдары болады. Пастерлеу процесінде бұл бактериялар егер ашыту тағы керек болып оларды тағы сүтке қосқанда өлуі мүмкін. Ашыту жағдайына тәуелді сүт құрамы және көзі болатын қоспаларды қолданғанда әртүрлі өнімдер пайда болады. өнімдердің консистенциясы мен дәмінің айырмашылықтары бар. Оларды өзіне ірімшік, иогурт, сүзбе, май, қаймақ түрлерін қосып алатын спекторлары арқылы көруге болады.
Иогурт
Ірімшікке қарағанда өндіруге қолданылатын белгілі сүт фракциясына қарағанда иогуртты дайындау кезніде қаймағы алынбаған сүтті ашытады. Басында иогут сұйық және қышқылды болып жасалатын бірақ, Батыс елдерінде оның дәмі мен консистенциясын ұлттық дәстүрге қарай өзгертті. Себебі иогуттың өзі қышқылды болып табылады(pH 3,7—4,3). Оның дәмін жақсарту үшін жеміс - жидектер қосады.
Иогуртты дайындау үшін сүттің әртүрлі түрлерін: бөлек – бөлек немесе сұйықтықта сонымен қатар қаймағы алынбаған, алынған, жартылай алынған, қайнатылған және құрғақ түрлерін қолдануға болады. әр комбинация иогурттың ерекше түрін береді. Ең көп таралғаны төменгі май құрамы бар сапа. Бұл процесс схемасы 12.20 суретінде келтірілген.
Ұлыбританияда барлық сүт пастерлеуіне ұшырайды. Әдетте ол 72°С – та 15с- да жүргізіледі. Бұл патогенді микрооргнаизмдерден құтылу үшін жасалады. Олар міндетті түрде сүтте болуы тиіс емес, бірақ сүт микроағзалардың өсуі үшін тамаша ортасы болып табылады, өйткені, әрқашан зиянды бактерия, мысалы Salmonella сияқты бактериялар, немесе саңырауқұлақтар ластанған болуы мүмкін. Осындай бактериялардан бөлек кейбір бактериялар өз – өзіне зиянды емес және олар сүтте көбейіп оның сапасына әсерін тигізуі мүмкін. Себептерінің қатары 12.20 суретінде көрсетілген, иогурт дайындауындағы қажетті термиялық өңдеу. Сүт қышқылдарын алмастыру үшін, қыздыру алдында жоғплғандар, «әуелгі дақылдарға» қосылады. Сүт қышқылы және спирт, йогуртқа тән дәм береді, және Streptococcus thermophilus өндіретін, ерекше крем дәм қосады. Әуелгі дақылдар әдетте Lactobacillus Bulgaricus қамтиды. Бұл бактериялар йогурт Грам-боялған үлгідегі (бөлім 12.9.2) көруге қатынасы шамамен тең болуы мүмкін. Бактериялар екі түрі де Грам бояуымен боялған, сондықтан қызылға айналады. Сіз бактериялардың санын санау үшін тырыссаңыз, осы түрлердің әрқайсысы үшін бір грамм шамамен 108 жасушаларын таба алады.Бактериялардан жинақталған дақыл бақыланбалы және
болжамдалатын қаймағы алынбаған сүтті ашытқылайды. Бастапқыда, йогурт көп сұйқылтым және қышқыл болады, бірақ содан кейін оның дәмі мен құрылымы батыс елдерінде айтарлықтай ұлттық дәстүрлерге сәйкес өзгерді. Иогурт қышқылтым (pH 3,7—4,3)болғандықтан, оған оның дәмін өзгерту үшін әртүрлі жеміс – жидектерді қосады.
Ірімшік
Әуелі Ірімшік сүт өндіруде сүзбе және сарысуға бөлінеді. Осы мақсатта сүтке мұқият бақыланбалы керекті микрооргнаизмдер тазартылған жағдайларда әуелгі дақылға енгізіленеді. Сүтте сақталатын казеин ұюы арқылы процессті құрағыш сүт қышқылы сүтті ашытуды бастайды. Бұл кезде сүттің «ұюы»болып жатады. Құрамында реннин бар іріткі ферментінің қосылуы, коагуляция жеделдету және одан әсіресе оны бақылауға мүмкіндік береді. Әдетте, іріткі бұзау асқазанынан алынады, бірақ протеиназдың өзіне лайықты қасиеттерін саңырауқұлақтардың түрлерінен, бактерия штаммдарынан, гендік инженериясының модификсириялық әдістерінен айыруға болады. Осы жаңа ферменттердің артықшылығы кошерлық және вегетариандық ірімшік дайындау үшін оларды пайдалану мүмкіндігі болады.
Сүзбелік массаны бөледі, сығады, қажетті пішінді беріп және кептіріледі. Сарысу өз кезегінде, өңделгеннен кейін оны малға азық ретінде бергеннен соң оны қорек көзі ретінде ашытқылардың өсуі ретінде қолданады. Өнімде ақуыз және дәрумендердің көп мөлшері бар.
Ірімшіктің дәм беруі және процесс барысында пісіп жетілуі оған әртүрлі микрооргнаизмдер қосады: бактериялар (сырдың бір түрі), өңезденген саңырауқұлақ (қой сүтінен жасалған ірімшіктің бір түрі және тағы басқа да көк ірімшіктер) немесе басқа тәсілдері. Сапасы әртүрлі ірімшіктердің әртүрлі дайындалуындағы барлық процесстердер кезінде бақыланбалы жағдайлар жасалынылады. Кейбір ірімшіктер, шикі күйінде ауылдық және кілегей түрінде қала береді.
Май
Май ең кілегейден жасалады, жаңа сүттің қосындысынан жасалады, және де заң бойынша ол 80%майдан төмен болмау керек. Ең бірінші, кілегейді пастерлейді, содан соң шайқайды.
12.12.3. Біржасушалы уыздар
Жаңа қоректік көзі болып табылатын бір жасушалы уыздар. Оның шығарылымы 1960 жылы басталады. Ірі масштабтағы микрооргнаизмдерөсіретін мысалы, балдырлар ашытқыштар, саңырауқұлақтар тобы уыз терминіне жатады. Уыз адам пайдасына және жануарлардың жемі болып саналады. Онда көптеген витаминдер көзі, дәрумендер, көміртегілер мен қажетті майлар. Теориялық жағынан бұл уыз тағамдарды ұн, бидай сияқтылардың орнын басады. Бірақ батыста ауыл шаруашылықтарының шикізаты барынша жеткілікті болуы мүмкін ел. Әлемдік масштаб бойынша, уыздардың жетіспеуі байқалады. Себебі, бұл елдерде биотехнологияның жұмысы дамымаған. Микрооргазимдердің қорек көзі ретінде пайдалануы төменде рет – ретімен көрсетілген:
- Микроорганизмдер өсімдіктер мен жануарларға қарағанда аз орын алады;
- Микроорганизмдер тез өседі;
- Микроорганизмдер арзан тамырлардың кеңінен қолдануға пайдалынылады. Сонымен бірге, ауыл шаруашылығында, мысалы, метан, метиль және этиль спирттерінде, сірнеде, ірімшікте, қан сарысуында және қағаз қалдықтарында қолданады.
- Микрооргнаизмдерді шығарғанда этикалық жағынан қиындықтар көп туындамайды және жану құқығын да бұзбайды.
- Мироорганизмдерге гендік инженерия әдісіг қолдану оңай;
- Микрооргназим көптеген уыздарды құрайды.
- Микроорганизмдер климат жағдайына бағынышты емес жәнежердің көп мөлшерін алмайды. Ең алғашқы басты өнім болып прутин алынды.
- 1970 жылы мұнайдың қымбаттауының себебінен метанол энергияны көп қажет етті.
- Дамыған елдерде, мысалы, АҚШ, Еуропа бірасушалы уыздардың шығарылуы керек еді. Бұл елдерде ауыл шаруашылығыныңөнімін қайта шаруаландыру әдетке айналдырылды. Бұл өнімдердің құрылымына біржасушалы уыздардың және байытылған протеиндер қосылды, мысалы, бидай, соя және сүт тағамдары.
- Жаңа дамып келе жатқан елдерде уыз керектігі байқалып, инвестицияның заттармен өздерінің уыздық заттарынан шығара алмады.
- Жануарларға беретін қоректік тағамдардың шығарылуы көбейіп, бағасы төмендеді. Соңғы осындай тағамның биотехнологиялық тағам отын шығару болды.
Прутин
1980 жылы ІСІ компаниясы жұмысқа үлкен 1500м3, биіктігі 60 метр ферменттерді қосты.бұл Англияның солтүстік -батысындағы Билленгем қаласында болды. Оның құрамы фирмаға 40 млн фунтқа шықты. Ал жылдық 1 жасушалы уызын 70000 т шығарды. Бұл жерде дәндә дақылдар Methylophilusmethylotrophus бактерияны пайдаланды, ол оттегі мен қуат күшінің көзі болды және арзан әрі тез табылатын метанол табылды. Жұмыс аэробты жағдайларында жасалды. Бұл жерде шикізаттар қолданылды.
Органикалық оттегі
(метанол)++ Азот + —► + Н20 + минералды тұздар + жылулық+қуат + көміртегі
Қос тотықты көміртегіні балондарға толтырып, сатты. Мұндай бактерияны келісіп отырып, ең жоғарғы жылдамдығы өзі арзан көміртегімен қуат көзін , сонымен қатар термотұрақтысы болу керек . себебі, ферментация кезінде жылу бөлінеді және бір құрғақ массаға өте жоғары құнардағы уыз болды.
Methylotrophus бактериясы уақыт генерациясы болды. Ол 2 – 5 сағатқа дейін уһұстайтын болды. Азотты амони тұзы түрінде қосты. Сонымен бірге минералды қоспалар, фосфорлар және калий қолдануда.
Метанолды Биллингемнің табиғи газынан алды. Сонымен қатар, ол газ қосымша зиянды заттарды құрамады. Температураны қатаң қадағалап отырды. Ол 30—40 °С рН – ты құрады. Ферментер бірнеше ай бойы тоқтамай жұмыс жасап тұра алды.
Прутиннің шығарылымы дақылды өсірудің классикалық түрі болды. Оның негізі суреті 12.10.5 суретінде көрсетілген. Сондықтан оны, аэрациялау мен салқындатылуы жеңіл болды. Ол ауа берудің ең көрнекті жолы болып табылады. Ферментер түбіне қысымдалған ауа, көпіршіктер көтеріле кем жұмыс жасады. Жылына 6000т уыз алынды. 2000 тонна метанолдан 1 т кептірілген прутина алды. Оның құрамында 72% уыз және 8% ылғал болды. Жіне ең керекті аминқышқылмен, дәрумендер мен байылып, 2 есе қоректі болып шығарлды. Прутинді жануарлардың жемі ретінде пайдаланды. Дәнді дақылдардың жұмысы үздіңсіз юолғаннан кейін, бактерияларды ылғи жинап отырды. Жартысын ферменттерден шығарды, реагентпен тазалады. Шыққан жасушаларды желдетіп, кептірді. Ал қоректі сұйықтады ферменттерге қайтадан қайтарылды. Кептіріп болғаннан кейін бактериялық жасушаларды үгітеді. Себебі, қорауһптау алдында минералды құрамы мен рН-ы қадағалануы керек.
Микопротеин
Бір жасушалы ақуыздың Тағы бір әлеуетті көзі – саңырауқұлақтар.
Ашытқы сондай-ақ бактериялар, адам мен жануарлар пайдалануға болады (төменде қараңыз. Sec. «Ашытқы сіріндісі»). Қолданыс табады және өңезденген саңырауқұлақ. Мицелия дегеніміз - жіп – жіңішке гиф жиынтығынан тұрады, оларға типтік саңырауқұлақтық тән. Корам саңырауқұлақтарды қолдану үлгісі - микропротенді шығару. 1985 жылы Рэнк, Ховис және Макдоугал Marlowfoods компаниясымен біріге отырып, «Quorn» атты маркалы микропротеин шығарды. Осы мақсатта Fusariumgraminerium атты санырауқулақ пайдаланды.
Ол алғаш рет 1960 жылдары Марл оу топырақ үлгісі Бакингемширеге жақын шықты , осыдан компанияның атауы шықты. Ақуыз өнімін зерттеу 1964 жылдан басталды , ал адамдар үшін олардын қолданылыуын жане қауіпсіздігін 1986 алды . Басында кворнды хош иісті бәліштерді дайындау үшін қолданды, оны Дж. Сэйнсбери сатты.
Бірақ ол бәліш мен карри сияқты азық – түлік өнімдерін өндіру үшін қолданды, 1990 жылы сатылымға үйде пайдалануға кворн ұсақ тілімдермен келді, Ал 1992 жылы оны ұнтақ түрде сата бастады. Кворн сатып алушылар арасында жұмсак дәм жане хош иісімен табыс алды.
Саңырауқұлақты дәнді дақылдаумен өсіру уақыты 5,5 сағ құрайды. Демек, ол бактерияға қарағанда жай өседі. Көміртегі көзі және қуат көзі ретінде саңырауқұлағындағы глюкозаны қолданады. Оны кез келген арзан крахмал құрайтын өнімнен жүгеріден, бидайдан, күріштен, картоптан және млессадан. Саңырауқұлақ 1 кг қантқа 0,5 кг құрғақ биомассаны құрайды. Санырауқұлақтардың артықшылығы олар рН қышқыл жерлерде өсе алады. Бұл кезде бактериялардың өсуі тоқтатылады. Сөйтіп, дәнді дақылдың бұзылуына әкеліп соқтырады. Fusarium 30 градус жерлерде өседі. Бактерияларды өсіру секілді, азоттың көзі аммони тұздары болып табылады , және де олардың өсуіне минералды тұздарды олардың өсу ортасына қосады. Жоғарыдағы барлық үдерістердегідей, аэрация және суық орта қажет, ал қолданылатын барлық материалдар тазартылған болуы қажет. Аэрациялау механизмі арқылы ортаны араластыру жұмысы жүргізіледі. Бұл жұмыстарда механикалық қалақшаларлың жұмысы қажеті жоқ. Өйткені, қалақшалар тепе – тең ауданының жұмысын бұзады. Бұлардың жұмыс жасау механизмі ауа лифтісі деп аталады. Себебі, өнімдер үнемі қозғалыста болады. әрбір 2 мин сайын ауа көмегімен 1 айналым жасайлы. Ферменттердің аумағы 40000 дм 3 – ті құрайды. Осы ферменттердің үздіксіз жұмысына ылғи таңдалған өнімдермен жаңа орталар өажет.
Прокариотикалық жасушаларға қарағанда эукариоттық жасушалар өте жоғары нуклеин қышқылын құрайды. Сондықтан микропротеиндар аса маңызды көлемде нклеин қышқылының 5 - 15% құрғақ массасын құрайды. Бұл дегеніміз алдындағы мақсатта РНҚ оның құрамын азайту керек. Себебі, адам оны 2 күннен артық қолданбауы қажет.бқл дегеніміз бөтекеде тас олуы және подогра ауруына әкеліп соқтыруы мүмкін.. РНҚ дақылын алып тастау үшін 64 °Сқа дейін 20 – 30 мм арнайы дыс ішінде ұстап, қайнатылған бу жібереді. Қыздыру барысында протеиназдар белсенділігі төмендейді. Соөйтіп, ақуыз бұзылмайды. Бірақ , бұл РНҚ – ға қатысы жоқ. РНҚ құрамы бүкіләлемдік денсаулық орталығының жарлығы бойынша ең төмен 2 % көрсетілсе, бұл 1% - ға дейін төмендейді.
Мицелий саңырауқұлағын дәнді дақылдар орталығына бөлуге, бактерия жасушасын бөлуге қарағанда оңайырақ. Бұларды центрифугациялау қажет емес. Мицелийді жай сүзіге болады. сүзіп және кептіруден кейін, ол жұқа пленка тамыры болып табылады. Бұл сыртқы түріне қарағанда, шикі қамырды елестейді. Оған көкөніс қосқыштарын және аздаған жұмыс уызын қосады. Содан соң бөлшектетіп кеседі немесе ұсақтап сол түрінде сатылымға шығарады. Саңырауқұлақ табиғатынан етке ұқсайды. Сондықтан оған еттің дімңн оңай қосыға болады. адмның қолдануы жайында әңгімелегенде оның экономикалық факторларынан басқа көптеген жақтары қаралады. Мысалы, қолдану сақтылығын қатаң қадағалау. Заттың қорек санағы жоғары болуы. Осылардың бәрін 10 жылдай зерттеп, зиянды жолын таппады.
Тексерілу кем дегенде 4 ғасыр бері қарай 11 түрлі жануарларда, соның ішінде тышқан, шошқа мен сиырларды тәжірибе жүргізілді. Микропротеин шықпас бұрын өз еркімен келіскен адамдарды тексерілу жүргізілді. Алынған нәтижеде өнім адам денсаулығына еттен көрі пайдасы бар екенін көрсетті.
12.5. кесте қолданатын кейбір типті уыздармен майлардың құрамы көрсетілген. Бұлар холестеролды құрайды. Және майдың коллориясы төмен және бір жақсы қасиеті дәрімен көзі, В12 және цинк қоспасының
12.11. Кворнның проценті жарнамада табиғи өсімдік түйіршігі деп жазылады. Неге олай жазылған?

Ашытқыш экстраты
Ашытқыштар сыра шығарылыдары әр түрлі жолмен алынады. Белгілі бір, мысал ретінде вискиденашытқыштың алынуы. Мұнда да сырадағыдай секермен ашытып барып, арпа тұқымымен ашытып алынады. Бірақ вискиді уыттан тазартылынып алады. Бұл жерде қайнату керек болады, ал қайнатылған кезде ашытқылар өледі. Олардың ортаны татыру үшін сыра қалдықтарынан қалған ашытуды пайдалынады. Ашытқыш жас дәрумендердің В тобына бай. Соның ішінде ниацин (В6), рибофлофин (В1), тиамин (В), рибофламин (В1) фолио қыщқылы және В12 дәрумені.
Ашытқыларды құруға және одан дәрумендер таблеткаларын жасауға болады. сонымен бірге мармит жасауға болады. ол үшін ашытқышты 50 °С қыздырады. Содан соң үлкен көлемде тұз қосады. 5 .10.6. бөлімдерінде қарайсыңдар.
Автолиз дегеніміз юұл өліп қалған жасушалардың әсерінен ферменттердің қайта қорытылуы. Өнімді сүзіп , сонан соң центррифигациялап, қабырға жасушасынан тазаланады. Сонан соң қоюлап, консестанциясын қосады. Мармитке көкөніс қоюуын қосады.
Альтернативтік автолиз ашытқыларф қимылының әсерінен болған гидролиз болып табылады. Кейін гидролизді гидрооксид гидроксид нартиймен тазартады. Өнімді мысалы, чипсыға, гамбургерге, сһботқаға, соусқа, тұздыққа, ащы қышқылға дәм беру үшін еттің дәміне ұқсас дім беру үшін қосады.
Фотосинтездік ағзадағы біржасушалы ақуыздар
Фотосинтездегін организмдердің біржасушалы жолдың алынуы үшін фотосинтездік бактериялар сияқты. Мысалы, ценабактериялар және балдырлар алынады. Цианобактерияларының мысалы ретінде спирулина алуға болады. оны табаннан жасалған батыс Африка үшін бұл басты тағам. Бұл спирулинді азғантай масштабта Мексикада және Гавай аралдарында диеталық тағам ретінде сатады. Ол өте көп жасалынады. Оны көлмен көлшіктердің бетімен жинайды. Бір жасушалы балдырлардың мысалы, хлорелла жатады. Жапонияда және тайванда көптеген хлорелланы денсаулыққа сапалы тағам ретінде сатады.
12.1.1. және 12.1.2. бөлімдерінде фотосинтезделген организмдердің өсуінің жоғарғыда көрсетілген. Бұларда өсімдіктің негізгі көзі – жарық пен оттегі болғандықтан. Ерекше айырмашылығы саңырауқұлақтардың фотосинтезделмеген бактерияларды айырмашылығы бар. Негізі оларды стирильденбеген жағдайда су ағымдарында өсіреді. Егер балдырларды тұрған су бетінде 2 есе пайда береді. Бұл Израилда қолға алынған.
12.12. Ауыл шаруашылығы
Бұл бөлімде биотехнологияның жетістіктерін ауыл шаруашылығындағы кейбір жолдарында қарастырамыз.
12.13.1. гендік инженерия
Гендік инженерияның ауыл шаруашылығына байланысты жоғарыда айтқандай гендік инженерияны биотехнологияның бір түрі ретінде қарауға болады. оның ауыл шаруашылықтарында қарастырылған өндіріс сомототрапин өсімдігі. Бұл пестицит және гербициттерге қарсы құрал. 25 бөлімде гендік инженерияның практикасында жазылған.
12.13.2. Сүрлемдік
Сүрлемді дайындау ферменттерінің анаэробтық процесі бұл фирмаларда қолданады. Сүрлемдік өнімнің қоректік құндылықтарын ауыл шаруашылығынң жануары үшін жақсы сақталады. Мысалы, қыс кезінде малдың қорын сақтау. Шөптің жоғары бетінде сүт қышқылы бақылауының лактобацилус құрайды. Шөпті шауып, ұсақтағаннан кейін, оны үлкен бункерге тиейді. Кейінірек, бұл үшін қарапластидтерге оралған тюктерді пайдаланды. Бактерий шөптің құрамындағы табиғи қантты, глюкозаны, фруктозаны, сахарозаны қуат көзі ретінде анаэробтық демалу үшін қоректі ферменттерді сүт қышқылына айналдырады. Негізгі өнім болып, сүт қышқылы қолданылады. Бірақ ферментация кезінде басқа заттар да қосылады. Олар дәммен кереме иіс 4 – ке дейін дөмендейді. рНтың бұндай төмен түсуі шірігіш бактериялардың белсенділішін тоқтатады. Ондай болмаған жағдайда шөптің қоректік құндылығы азайып, немесе шіріп кетуі мүмкін. Ферментация кезінде температура қатты көтерілу мүмкін. Сондықтан бактериялар термотұрақты болуы қажет. Шөптердің листерия бактерияларымен сақтану керек. Себебі, ол ауыл шаруашылығында жануарлардың өлі туылуына себеп болады. бұндай ластану, егер ферментация белсенділігін рН 5,5 төменде сақталса, ластану болмайды. Қазіргі таңда сүрлемдіктің шығарылуы едәуір көтерілді. Сонымен қатар оны тез өсіргіш бактериялар коммерциялық жағдайда қолданылады. Олар көпіршік және сұйық ретінде жаңа табиғаттағы шөп ретінде сатылады. Биллингемдегі ферменттерде мұндай ферменттің 3 түрі шығарылады:
12.13.3. Азотификация
Бұршақ дәнді дақыл ортасына горох беде, жоңышқа азот пен қамтамасыз ету үшін көптеген ғасырлар бойынша қолданылып келе жатыр. 19 ғасырда осы өсімдіктердің тамыр түптері ауадан азто юөліп, оны нитратқа айналдыратын қасиеті бар екенін біледі. Бұл жағдайда 10.4.1. бөлімінде жазылған бактерий, ризоблум штаммының түрін құрайды. Сурет 12.22. белгілі бір штаммдар бойынша бұршақтардың спецификалықтрлерін залалдайды. Көбіне бактерий тамырды шірітеді. Бұршақ тұқымдастары табиғатында ризибиум өсетін жерде болатындықтан, тамырын шірітеді екен. Бірақ, бұршақтұқымдарын бактерий бөлшектеп, жерге өсірсе болады. ризобиум штаммыныңкерек дақылын, ферментін өсіреді. Содан соң қолайлы ортаға асырайды. Көбінесе стирильді торфқа өсімдік өнімін қосуға болады. мысалы, борозда. Дақылдардың күнделікті өсіріліп отыратын жерінде залалданған тұқымдар қажет емес.
Резобиум көп жылдан сақталғандықтан қарапайым органикалық заттар пайдалана отырып, аммони немесе нитраттар азот көзі болып табылады. Бактерий азотын тек тамырлардан табады. Эксперттің мысалы болып табылатын беде Австралияда шығуы саналады. Бұршақ Солтүстік Америкадан, жоңышқа Еуропадан шыққан. Егер осы дақтардың өсіруде пайдалану керектігі азаятын болса, осы дәнді дақылдарды егуде, азоттық тыңайтқыштарының керектігі азаяды.
Ашытқыштар сыра шығарылыдары әр түрлі жолмен алынады. Белгілі бір, мысал ретінде вискиденашытқыштың алынуы. Мұнда да сырадағыдай секермен ашытып барып, арпа тұқымымен ашытып алынады. Бірақ вискиді уыттан тазартылынып алады. Бұл жерде қайнату керек болады, ал қайнатылған кезде ашытқылар өледі. Олардың ортаны татыру үшін сыра қалдықтарынан қалған ашытуды пайдалынады. Ашытқыш жас дәрумендердің В тобына бай. Соның ішінде ниацин (В6), рибофлофин (В1), тиамин (В), рибофламин (В1) фолио қыщқылы және В12 дәрумені.
Ашытқыларды құруға және одан дәрумендер таблеткаларын жасауға болады. сонымен бірге мармит жасауға болады. ол үшін ашытқышты 50 °С қыздырады. Содан соң үлкен көлемде тұз қосады. 5 .10.6. бөлімдерінде қарайсыңдар.
Автолиз дегеніміз юұл өліп қалған жасушалардың әсерінен ферменттердің қайта қорытылуы. Өнімді сүзіп , сонан соң центррифигациялап, қабырға жасушасынан тазаланады. Сонан соң қоюлап, консестанциясын қосады. Мармитке көкөніс қоюуын қосады.
Альтернативтік автолиз ашытқыларф қимылының әсерінен болған гидролиз болып табылады. Кейін гидролизді гидрооксид гидроксид нартиймен тазартады. Өнімді мысалы, чипсыға, гамбургерге, сһботқаға, соусқа, тұздыққа, ащы қышқылға дәм беру үшін еттің дәміне ұқсас дім беру үшін қосады.
Фотосинтездік ағзадағы біржасушалы ақуыздар
Фотосинтездегін организмдердің біржасушалы жолдың алынуы үшін фотосинтездік бактериялар сияқты. Мысалы, ценабактериялар және балдырлар алынады. Цианобактерияларының мысалы ретінде спирулина алуға болады. оны табаннан жасалған батыс Африка үшін бұл басты тағам. Бұл спирулинді азғантай масштабта Мексикада және Гавай аралдарында диеталық тағам ретінде сатады. Ол өте көп жасалынады. Оны көлмен көлшіктердің бетімен жинайды. Бір жасушалы балдырлардың мысалы, хлорелла жатады. Жапонияда және тайванда көптеген хлорелланы денсаулыққа сапалы тағам ретінде сатады.
12.1.1. және 12.1.2. бөлімдерінде фотосинтезделген организмдердің өсуінің жоғарғыда көрсетілген. Бұларда өсімдіктің негізгі көзі – жарық пен оттегі болғандықтан. Ерекше айырмашылығы саңырауқұлақтардың фотосинтезделмеген бактерияларды айырмашылығы бар. Негізі оларды стирильденбеген жағдайда су ағымдарында өсіреді. Егер балдырларды тұрған су бетінде 2 есе пайда береді. Бұл Израилда қолға алынған.
12.12. Ауыл шаруашылығы
Бұл бөлімде биотехнологияның жетістіктерін ауыл шаруашылығындағы кейбір жолдарында қарастырамыз.
12.13.1. гендік инженерия
Гендік инженерияның ауыл шаруашылығына байланысты жоғарыда айтқандай гендік инженерияны биотехнологияның бір түрі ретінде қарауға болады. оның ауыл шаруашылықтарында қарастырылған өндіріс сомототрапин өсімдігі. Бұл пестицит және гербициттерге қарсы құрал. 25 бөлімде гендік инженерияның практикасында жазылған.
12.13.2. Сүрлемдік
Сүрлемді дайындау ферменттерінің анаэробтық процесі бұл фирмаларда қолданады. Сүрлемдік өнімнің қоректік құндылықтарын ауыл шаруашылығынң жануары үшін жақсы сақталады. Мысалы, қыс кезінде малдың қорын сақтау. Шөптің жоғары бетінде сүт қышқылы бақылауының лактобацилус құрайды. Шөпті шауып, ұсақтағаннан кейін, оны үлкен бункерге тиейді. Кейінірек, бұл үшін қарапластидтерге оралған тюктерді пайдаланды. Бактерий шөптің құрамындағы табиғи қантты, глюкозаны, фруктозаны, сахарозаны қуат көзі ретінде анаэробтық демалу үшін қоректі ферменттерді сүт қышқылына айналдырады. Негізгі өнім болып, сүт қышқылы қолданылады. Бірақ ферментация кезінде басқа заттар да қосылады. Олар дәммен кереме иіс 4 – ке дейін дөмендейді. рНтың бұндай төмен түсуі шірігіш бактериялардың белсенділішін тоқтатады. Ондай болмаған жағдайда шөптің қоректік құндылығы азайып, немесе шіріп кетуі мүмкін. Ферментация кезінде температура қатты көтерілу мүмкін. Сондықтан бактериялар термотұрақты болуы қажет. Шөптердің листерия бактерияларымен сақтану керек. Себебі, ол ауыл шаруашылығында жануарлардың өлі туылуына себеп болады. бұндай ластану, егер ферментация белсенділігін рН 5,5 төменде сақталса, ластану болмайды. Қазіргі таңда сүрлемдіктің шығарылуы едәуір көтерілді. Сонымен қатар оны тез өсіргіш бактериялар коммерциялық жағдайда қолданылады. Олар көпіршік және сұйық ретінде жаңа табиғаттағы шөп ретінде сатылады. Биллингемдегі ферменттерде мұндай ферменттің 3 түрі шығарылады:
- Екосул
- Екорот
- Есебол
12.13.3. Азотификация
Бұршақ дәнді дақыл ортасына горох беде, жоңышқа азот пен қамтамасыз ету үшін көптеген ғасырлар бойынша қолданылып келе жатыр. 19 ғасырда осы өсімдіктердің тамыр түптері ауадан азто юөліп, оны нитратқа айналдыратын қасиеті бар екенін біледі. Бұл жағдайда 10.4.1. бөлімінде жазылған бактерий, ризоблум штаммының түрін құрайды. Сурет 12.22. белгілі бір штаммдар бойынша бұршақтардың спецификалықтрлерін залалдайды. Көбіне бактерий тамырды шірітеді. Бұршақ тұқымдастары табиғатында ризибиум өсетін жерде болатындықтан, тамырын шірітеді екен. Бірақ, бұршақтұқымдарын бактерий бөлшектеп, жерге өсірсе болады. ризобиум штаммыныңкерек дақылын, ферментін өсіреді. Содан соң қолайлы ортаға асырайды. Көбінесе стирильді торфқа өсімдік өнімін қосуға болады. мысалы, борозда. Дақылдардың күнделікті өсіріліп отыратын жерінде залалданған тұқымдар қажет емес.
Резобиум көп жылдан сақталғандықтан қарапайым органикалық заттар пайдалана отырып, аммони немесе нитраттар азот көзі болып табылады. Бактерий азотын тек тамырлардан табады. Эксперттің мысалы болып табылатын беде Австралияда шығуы саналады. Бұршақ Солтүстік Америкадан, жоңышқа Еуропадан шыққан. Егер осы дақтардың өсіруде пайдалану керектігі азаятын болса, осы дәнді дақылдарды егуде, азоттық тыңайтқыштарының керектігі азаяды.

12.14. Отынның биологиялы массадан алынуы – жаңа отын көзі
Көмір, мұна, ағаш, торф немесе құрғақ зәрді пайдалану табиғи жанармай болып саналады. Бұл жанармай көздері жыл өткен сайын азаяды, бағасы қымбаттайды. Сол себепті тірі ферменттермен биологиялық жұмыстар жасай отырып, жаңа отын көзін табу ойластырылуда. Алдағы уақытта қолдан жасалған фотосинтез судан сутегіні алу арқылы отын ойлап табуға болады. және бір басқа жолы энергияны юиомассада басқа формамен сақтап, жанармай май табу жолын іздеуде. Қазіргі уақытта тексеріліп жатқан материалдарға ағынды судың тоқтауы, үй шығындары, азық – түлік шығындары, маколатура, ауыл шаруашылық дәнді дақылдары плынады. Соныен бірге, ауыл шаруашылығының дәндерінде жүгері, қант қызылшасы, су балдырлары да тексеруге алынуда. Жылу көзін табуда: екі бағыттағы жұмыс істеген, соның ішінде биогаздың жиналуы мысалы, метан, биогаздың бактериямен жиналуы, этиль спиртінің ашытқымен қосылуы. Екі жағдай да анаэробты болып саналады.
12.14.1. Биогаз
Жалпы теңдеуі:
С6Н|206 ►ЗСН4+ ЗС02
Глюкоза Метан Көміртек диоксиді
Қуат мәні: 16 кДж г-156 кДж г1
Биогаз 54—74% метанды құрайды. Қалғандары көміртек газдың негізгі түрлерін, арнайы сапалы азотты, сутекті және тағы басқа газдарды құрайды. Ферментация кезінде түгел микроорганизм қатысады, соның ішінде метоноген, мысалы, Methanobacterium , яғни олар оттекті газдан және судан метан бөліп шығарады. Шынайы бактериялармен тығыз байланыс жазайтын, бұрынғы организм топтары архебактерияларына жатады. Субстрат ретінде ферментацияға ұзін шоғыр қалдықтары немесе өсімдік өнімдері жатады. Қуатты өмірлік әлеуеті бар зауыт және бітелтіретін каналдар мен су жолдары болуы үшін - Америка Құрама Штаттарында су гиацинтін пайдаланылады. Жанармайы немесе отын бар жерде, мысалы ИНдия мен Қытайда, мұндай әдіс кіші масштабты өнеркәсіптерге сәйкес келеді.
Көңнен, жылына сиырдан шамамен баламалы 227л бензин метан сапасын алуға болады. газ сапасына мысал ретінде 0,5 кг көңді алсақ бір отбасының барлық адамына 1 тәулік тамағына жетіп артылады. Қытайда осылай 18 млн. шамасында отбасыға осылай құрылған. Газ әдетте тамақ пісіру үшін, трактор немесе машишаларға құю үшін және электрлық генератор үшін қолданылады. Үлкен масштабтағы газ қалдықтан қосалқы қорек ретінде, тазалағыш зауыттардан ағын суларды алу ретінде, мысалы, қант өндіру айналысушы. Газ ағын сулардың электр генераторын жіберілуі қоқыстардың зауыттарда қайта істелінуі сияқты жұмыстарда қолданылады. Англиядағы қоқыс метан көзі болып табылады, 1 кг қоқыс 20 л дейінгі газды алуға мүмкіндік береді. Қазіргі уақытта газды қалдықтарды көму ретінде жинауда, трубаларды жүктеп, нығыздалған қоқыстар және одан газды жинап шығарады. Қалдықтарды көму жерлерінде тапшылықтың, жетіспеушіліктің өсуі және ыңғайсыздық, осылар шақыратын бұл процесстің ашылуына назар аудара отырып күш жігер салу керек, ферментациямен байланысы бар осындай материалдар , қағаз кардондар тіпті бұдан әдеттегі түрден жылудан отын алатын болсақ та қымбат емес . Меншіктік табиғи отынның қорынан айырылу және ормандардың артықтарын сақтайтын дамыған елдерге экономикалық саясат қолайлы болып келеді. Кептірілгеннен гөрі отын бағасы ферменттік көңнен 6ға көп.
Көмір, мұна, ағаш, торф немесе құрғақ зәрді пайдалану табиғи жанармай болып саналады. Бұл жанармай көздері жыл өткен сайын азаяды, бағасы қымбаттайды. Сол себепті тірі ферменттермен биологиялық жұмыстар жасай отырып, жаңа отын көзін табу ойластырылуда. Алдағы уақытта қолдан жасалған фотосинтез судан сутегіні алу арқылы отын ойлап табуға болады. және бір басқа жолы энергияны юиомассада басқа формамен сақтап, жанармай май табу жолын іздеуде. Қазіргі уақытта тексеріліп жатқан материалдарға ағынды судың тоқтауы, үй шығындары, азық – түлік шығындары, маколатура, ауыл шаруашылық дәнді дақылдары плынады. Соныен бірге, ауыл шаруашылығының дәндерінде жүгері, қант қызылшасы, су балдырлары да тексеруге алынуда. Жылу көзін табуда: екі бағыттағы жұмыс істеген, соның ішінде биогаздың жиналуы мысалы, метан, биогаздың бактериямен жиналуы, этиль спиртінің ашытқымен қосылуы. Екі жағдай да анаэробты болып саналады.
12.14.1. Биогаз
Жалпы теңдеуі:
С6Н|206 ►ЗСН4+ ЗС02
Глюкоза Метан Көміртек диоксиді
Қуат мәні: 16 кДж г-156 кДж г1
Биогаз 54—74% метанды құрайды. Қалғандары көміртек газдың негізгі түрлерін, арнайы сапалы азотты, сутекті және тағы басқа газдарды құрайды. Ферментация кезінде түгел микроорганизм қатысады, соның ішінде метоноген, мысалы, Methanobacterium , яғни олар оттекті газдан және судан метан бөліп шығарады. Шынайы бактериялармен тығыз байланыс жазайтын, бұрынғы организм топтары архебактерияларына жатады. Субстрат ретінде ферментацияға ұзін шоғыр қалдықтары немесе өсімдік өнімдері жатады. Қуатты өмірлік әлеуеті бар зауыт және бітелтіретін каналдар мен су жолдары болуы үшін - Америка Құрама Штаттарында су гиацинтін пайдаланылады. Жанармайы немесе отын бар жерде, мысалы ИНдия мен Қытайда, мұндай әдіс кіші масштабты өнеркәсіптерге сәйкес келеді.
Көңнен, жылына сиырдан шамамен баламалы 227л бензин метан сапасын алуға болады. газ сапасына мысал ретінде 0,5 кг көңді алсақ бір отбасының барлық адамына 1 тәулік тамағына жетіп артылады. Қытайда осылай 18 млн. шамасында отбасыға осылай құрылған. Газ әдетте тамақ пісіру үшін, трактор немесе машишаларға құю үшін және электрлық генератор үшін қолданылады. Үлкен масштабтағы газ қалдықтан қосалқы қорек ретінде, тазалағыш зауыттардан ағын суларды алу ретінде, мысалы, қант өндіру айналысушы. Газ ағын сулардың электр генераторын жіберілуі қоқыстардың зауыттарда қайта істелінуі сияқты жұмыстарда қолданылады. Англиядағы қоқыс метан көзі болып табылады, 1 кг қоқыс 20 л дейінгі газды алуға мүмкіндік береді. Қазіргі уақытта газды қалдықтарды көму ретінде жинауда, трубаларды жүктеп, нығыздалған қоқыстар және одан газды жинап шығарады. Қалдықтарды көму жерлерінде тапшылықтың, жетіспеушіліктің өсуі және ыңғайсыздық, осылар шақыратын бұл процесстің ашылуына назар аудара отырып күш жігер салу керек, ферментациямен байланысы бар осындай материалдар , қағаз кардондар тіпті бұдан әдеттегі түрден жылудан отын алатын болсақ та қымбат емес . Меншіктік табиғи отынның қорынан айырылу және ормандардың артықтарын сақтайтын дамыған елдерге экономикалық саясат қолайлы болып келеді. Кептірілгеннен гөрі отын бағасы ферменттік көңнен 6ға көп.

12.14.1. Этанол
Жалпы теңдеуі:
С6Н,20* 2С2Н,ОН + зсо2
Глюкоза ^ Этанол Көміртек диоксиді
Қуат бағасы: 16кДжг-' 30 кДж г-1
Жалпы теңдеуі:
С6Н,20* 2С2Н,ОН + зсо2
Глюкоза ^ Этанол Көміртек диоксиді
Қуат бағасы: 16кДжг-' 30 кДж г-1

Бразилияда, 1975 жылдан бастап табысты отын ретінде пайдаланылады –этанол өндіруге. Шикізат ретінде қант құрағы болды. Коммерциялық тағам ретінде сахарозаны шырыннан жұлып алады және бұдан мелласса сиробы қалады, яғни ол глюкозадан және фруктозадан тұрады. Мелассаны басқа ферменттеу өнімдер тазарту үшін тазартылған ашытқы Saccharomcescerevisiae ретінде пайдалануға болады..Etanol арқылы ашыту үшін шикізат ретінде пайдалануға болады. Ұсақтау кезінде қант құрағы қалатын құрғақ талшықты материалды отын ретінде ажыратуда пайдаланылады.
Кейбір Бразилиялық автомобильдер таза этанолдың жұмыс істей алады, бірақ этанол адамдар оны пайдалануға жол бермеу үшін бензинді аз мөлшерде ішіне әдетте араласады. Ол 1985 жылы жасалды
11 млрд. Дм3 этанол. Бразилиялық валютаны сақтау мақсатында өндірістік мұнай бағасы өсті, ол 1983 жылы іске қосылды. Бензин тұтынуы жылдан жылға 20% -ға төмендеді. Бразилия мен Америка Құрама Штаттарында кейбір автомобильдер gasohol деп аталады,ол ішімдік пен бензин қоспасы арқылы жұмыс істей алады (Cурет. 12,24). Америка Құрама Штаттарында жүгеріні крахмал этанол өндіру бойынша биомасса көзі ретінде пайдаланылады. 1980 жылдардың ортасында. 2,28 млрд. Дм3 астам әр жал сайын өндірді.
Бразилиялық схемасы болғанына қарамастан, бұл табысты болған жоқ, бәрі біркелкі жүрді. Мелассаның экономикалық пайдасына, және оның ауыл шаруашылық дақылдарынының басқа да оттектің көзі ретінде қолданыста бола алмайды деген пікір - таластар жүріп жатыр. Мысалы, мелассаны мұнайымен бірге қосып, дымқыл материал ретінде пластмасс өндіруде қолға келеді.
Кейбір Бразилиялық автомобильдер таза этанолдың жұмыс істей алады, бірақ этанол адамдар оны пайдалануға жол бермеу үшін бензинді аз мөлшерде ішіне әдетте араласады. Ол 1985 жылы жасалды
11 млрд. Дм3 этанол. Бразилиялық валютаны сақтау мақсатында өндірістік мұнай бағасы өсті, ол 1983 жылы іске қосылды. Бензин тұтынуы жылдан жылға 20% -ға төмендеді. Бразилия мен Америка Құрама Штаттарында кейбір автомобильдер gasohol деп аталады,ол ішімдік пен бензин қоспасы арқылы жұмыс істей алады (Cурет. 12,24). Америка Құрама Штаттарында жүгеріні крахмал этанол өндіру бойынша биомасса көзі ретінде пайдаланылады. 1980 жылдардың ортасында. 2,28 млрд. Дм3 астам әр жал сайын өндірді.
Бразилиялық схемасы болғанына қарамастан, бұл табысты болған жоқ, бәрі біркелкі жүрді. Мелассаның экономикалық пайдасына, және оның ауыл шаруашылық дақылдарынының басқа да оттектің көзі ретінде қолданыста бола алмайды деген пікір - таластар жүріп жатыр. Мысалы, мелассаны мұнайымен бірге қосып, дымқыл материал ретінде пластмасс өндіруде қолға келеді.

Бактериялық сiлтiсiздендiру қазір бірінші кезекте мыс және уран, кеннен металдарды оқшаулау үшін қосымша әдісі ретінде бүкіл әлемде (сур. 12,25) пайдаланылады. Өзінің бірегей үлесін қосуда, олардың әрқайсысы бактериялардың бірнеше түрін тарту шаймалау. 300-ден астам млн. Доллардың құны 1983 жылы АҚШ-та 10% артық мыс шығарды. Бұл әдістің көмегімен алынды. Төмендегідей бактериялық шаймалау артықшылықтары болып табылады:
1. Ол құрамдылығы төмен кенді пайдалануға болады. Бөлу дәстүрлі әдістері өте қымбат металдар, ол тек өте бай металл кенін пайдалануға ұсынылады. Сондықтан, дамыту саласында дәстүрлі әдістерін пайдалана отырды, кейін әлеуетті өнім көп болды.
2. бактериялық шаймалау әдісімен металды алсаңыз, терең дамытусыз істеуге болады. Бірінші жарылғыш айып пайдаланып тастарды қиыршық, және ол содан кейін шаймалау ерітіндісін айдалады. Еритін металл тұздары бар ерітіндісін шаймалау аяқталғаннан кейін, жартастан бұрғыланған ұңғымадан айдалады. Бұл әдіс аз қымбат; ол пайдаланған кезде айтарлықтай бетінің рок үлкен көлемін төзбейді кезде, терең дамыту сияқты, қоршаған ортаны құнсызданған және қалдықтардың тауға қалыптаса қой2ған жоқ.
3. кеннен мыс өндіру дәстүрлі әдістері жоғары температураға талап етеді. Бұл әдістер, қымбат қазба отын тұтынады, демек, мысалы, қышқыл жаңбыр тудырады ауаны ластайтын,. (бактериялық шаймалау күкірт қосылыстарының шаймалау әдісімен қазба отын түрлерін емдеу үшін болашақта пайдаланылуы мүмкін.)
4.Тау-кен қалдықтарын бақылаусыз шаймалау ауыр металдармен жақын сулардың ластануына әкелді. Бұл бақыланатын шаймалау және металл қалпына келтіру болдырмауға болады. Ол батыс Құрама Штаттарында ғана тау-кен үйінділерде мыс астам 33 млн. Тонна деп бағаланып отыр. Әдетте жазықтарға орналастырылған қадалар, және, осылайша, металдар алыс қашықтықтарға өзен су асырылуы мүмкiн. Сілтілі бар суда еритін металл даунстрим бөгеті кезде жинайды және металдарды өндірумен айналысады зауытта, айдалады болады. Қажет болса, тазартылған су үйіндісіне үшін қайта өңдеуге болады.
5.Гендік инженерия арқылы, атап айтқанда, Т. ferrooxidanss жылы, бактериялар жақсарту үшін әрекеттер қолданылады.
12.16. Ферменттік технология
Бұл бөлімде жасушадан бөлінген ферменттердің биотехнологияда қолданылуын қарастырамыз.
Ферменттер бұлар жасушалардағы химиялық белсенділікті реттеуге мүмкіндік беретін биология катализаторлар. Адамзат 1000 жылдан бері ферменттерді қолдануына қарамастан, тек 19 ғасыр соңында олардың қалай жұмыс жасайтынын түсініп бастады. Қазір біз білеміз, ферменттер дегеніміз спецификалық 3 өлшемді конфигурация бар күрделі белоктық молекулалар. Және құрылымы ДНК да кодталған. Мүмкін атын конфигурацияның саны есепсіз көп болуы мүмкін.
Өнеркәсіп үшін ферменнттер екі негізгі себептерге байланысты тартымды. Біріншіден, әртектілігінің арқасында ферменттер көптеген өндірістік химиялық реакцияларды катализдеуге потенциалды қабілетті. Екіншіден, олар әдете қолданылатын органикалық емес катализаторлардан әлдеқайда тиімді және ерекше. Қалыпты температура мен қалыпты қысым кезінде олардың қатысуымен жүзеге асатын дәл сондай реакциялар әдетте жоғары температураны, жоғары қысымды талап етеді. Мысалы, Хабер тәсіліне негізделген әлемдегі ең үлкен өнеркәсіп бірінде, аммиак (NH3) газ түрунде азот және сутекте 500 °С температура және жоғары қысымда алынады.
Азотфиксациялаушы бактериялар аммиакты атмосферадағы азот және сутектен бөлме температурасында және қалыпты атмосфералық қысымда АТФ-ті энергия көзі ретінде, ферменттердің көмегімен синтездеуге қабілетті. Егер ферменттердің көмегімен аммония алу технология өңдеу етудің сәті түссе, онда өте кқп энергияны үнемдеуге болар еді. Ферменттердің тағы бір артықшылығы, өте таза өнім беретінінде-олардың специфаукостісі.
Бұл болса, фариокология, тамақ және ауылшаруашылық өнеркәсіп үшін өте маңызды.
Ферменттерді қолданудың бірқатар кемшіліктері, негізінен жасушадан бөлінген белоктардың тұрақсыздығына байланысты. Мұндай белоктар темпаратура және рҺ өзгергенде және өндірістік процесте қолданылатын органикалық еріткіштердің әрекеті кезінде тез денатурацияланады. Олар реакция өнімдерімен де түтік арқылы қызмет етуі мүмкін. Ферменттер өздерінің бөліну процестерінің жоғары құны да кемшілік болып саналады. Және бұл үшін «қауіпсіз» (мысалы, залалсыздар) организмді пайдалану қажеттігі, әсіріесе бұл ферменттер жануарлар мен адамдарға қажет өнімдерді алу үшін арналған болса, күні бүгін 2500 жазылған, суреттелген (өз кезегінде олар табиғатта табылғандардың жалпы санының тек 10 %құрайды). Ферменттердің 200ден азы қолданылады. Ферменттердің көпшілігі бар болғаны саңырауқұлақтардың 11 түрінен, ашытқының 4 түрінен және бактерияның 8 түрінен шығып отыр.
Болашақта жаңа ферменттерді құрастыру үшін кең мүмкіндік ашылады. Олардың біреуі – ферменттер кодтайтын гендерді өзгерту жолымен кейбір аминқышқылдарды бағытты түрде өзгерту. Белоктардың спецификалық үш өлшемді конфигурация алу заңын біз терең біліп отырмыз. Сондықтан жақын болашақта толық жаңа құрастырылған ферменттердің жарыққа шығуы теріске шығарылмаған. Бұл бағыт белоктық инженерия деп аталады. Бұдан басқа да, біз ферменттердің қалай жұмыс жасайтынын көбірек білеміз. Оның үстіне, мүмкін, біз әдеттегі ферменттерге қарағанда әлдеқайда тұрақты белоктық емес немесе ішінара белоктық емес катализаторды құрастыра аламыз. Мүмкін, дәл о сы бағыт кәсіпкерлер үшін тартымды, ұнамды болады.
Құны тез қайтарылатын бағыттардың ішінде – қазір қолданыстағы ферменттерге толық альтернативті болатын табиғи ферменттерді іздеу. Қазір ферменттерді, өндірістік көптеген формадағы компаниялар; сыра,пиво, шарап жасаушылар детергент, текстил, жеміс шырынын өндірушілер және тағы басқалар қолдануда. Ферменттерді әлемдік сату деңгейі күні бүгін, жылына 1 млрд доллардан асып кетті. 12.6 кестеде ферментті қолданудың кейбір мәліметтері берілген.
12.16.1. Ферменттердің көзі (бұлағы)
Микроорганизмдердің өсімдік және жануарлармен салыстырғанда ферменттердің көзі ретінде, жоғарырақ тұруы келесі себептерге :
Ферменттерді әдетте микрооргнаизмдерден алады. Мынадай сұрақтуындауы мүмкін?
Алдын – ала ферменттерді шығарғаннан гөрі ферменттерлерді тұтас жасушаларды пайдалану мықты емес пе?
Егер өнімді алу үшін тек бір химиялық реакция керек болады және бұған көре бір фермент керек болады. Олай болса, бұл процесс жасушада болып жатқан процеске ұқсас. Тазаланған ферменттерді пайдалану тұтас микроорганизмдерді пайдаланудан артық болуының себептері:
Егер ферментті микроорганизммен алу керек болса, бұл микроорганизмді үлкен көлемде алдымен, фенһрментерде өсіріп алады. Қоректік көзі ретінде глюкощаны қолданбауға тырысады. Себеі ол көптеген пайдалы ферменттердің синтезін басып кетеді. Көбіне үщдіксіз ферментациялауды пайдаланады. Тек глюкозамераздарды шығаруына мүнің қатысы жоқ. Көптеген ферменттерден өздігінен жасушалардан бөлінеді. Сөйтіп олар, жасушадан тыс фермент болып саналады. Оларды ішкі жасушаға қарағанда тазалауөте оңай . оларғ алдымен, сұйық ортадан бөліп алып, әртүрлі жолдармен араластырып, құрамын өзгертеді.содан соң өнімді керегінше тазалайды. Кейбір ферменттерді толық тазалау міндетті емес. Ферменттерді алынған түрінде пайдалануға болады. Мысалы, шырындарды дайындағанда, кір жуатын түйіршіктерді пайдаланғанда соалй жасайды. 12.17.7 бөліменде қараңыз.
12.16.4. Жеміс шырындарын дайындау
Ири алдымен өнімді ұсаұқтайды. Басқа –өсмдіктердей жеміс жасушаларынын қабырғасы айналган жасушалардан тұрады. Бұл қабырғаларында целлюлоза, полисахаритер, гемицелюлозалар болады. Қабырға жасушалары өте мықты, оларды бұзу қиын. Өнімнен сөлін сапалы үрде алу үшін целлюлозамен гемицеллюлоза қосады. Ферменттерді таңдағанда,төменгі рН температурасында жұмыс жасай алатындай қылып жинақтайды, себебі жеміс шырындары қышқыл болады. рНтың оптикалық көрсеткіші 4 пен 5ті құрайды.өсім жасушалары күрделі полисахаридтердің әсерімен ұстап тұра алады. Оларды пектин дейді. Олардың құрамында жасбысқыш зат болады. Сол себепті, ол джемнің кебуіне әкеледі. Жемістерді сақтағанда немесе ұсақтағанда олар суды ерітетін пектинге айналады. Сондықтан олар жеміс шырындарында болады. Төменгі температурада пектиндер колоидті суспензия бөліп шығарып, шырыннан бөлінеді. Бұл шырынға лайлы түр береді. Кейбір Америка, Англия елдері үшін шырынның бұл түрі жақпайды.ал негізінде жемісті шырынның ең жоғарғы көрсеткіші.егер шырынға пектиназ ферменттерін қосса, олар пектинді полисахаридке езеді. Және шырында өте төменгі температурада сақталалды. Сөйтіп, пектиназдар шырынды сұйық қылады.
12.26. бөлімінде қараңыз. Пектиназ көзі болып бактериялар саналады. Осындай жолмен сыра мен шарап та дайындалады. Ал бұлардың лайлығы ақуыздағы танниннің болуыннан болады. Сырадағы лайдың төмен болуы үшін, салқындатқанда протеиназды қосады. Бұларға пепсин мен попоин жатады.
12.16.5. Етті жұмсарту
Ет бұл негізгі бұлшық ақуызы. Бұлшықтар түйілген – түйілген ақуызды волоконды??? құрайды. жалғастыратын терісі де структуралық ақуызды құрайды. Сонымен бұлар кологен мен илластин. Кологен қатты созылмайтын, ал иллостин созылмалы сары волокондар. Етті шайнауға ыңғайлы қылып, бір сөзбен айтқанда оны езе отырып, ақуызбен қосылған терісін ұсақтайды. Бұл кезде волокна қысқарады. Және бір – бірінен тез айырылады. Ақуызды бұзатфын ферменттерді протеиназдар деп атайды. Етті жұмсарту үшін көбіне попеин қолданылады. Оны попоя өсімдігінің шырынынан алады. Бұл фермент қосып езілген еттер үй жағдайында сатылымда кездесед. Қолданар бұрын етті белгілі бір уақытқа өзінің шырынына қояды. Бұл кезде ферменттер ө жұмысын жасайды. Кейбір елдерде попоинді малды сояр алдында қанына жібереді. Сөйтіп, сойылған малдың еті өте жұмсақ болады. Осы сияқты етті езі процестері өлімнен кейін жүреді. Өлген жасушалар лизосомдар деп атайды. Өлген жасушаларда лизосомалар бұзылады. Олардан ферменттер бөлініп шығып, бұзылу жұмысы басталады. Бұл процесс автолиз деп аталады.
5.10.6. бөлімдерін қараңыз
Сондықтан малдың ет туштарын қолданар алдында бірнеше күнге салқын қоймаларда бірнеше күнге қояды.
1. Ол құрамдылығы төмен кенді пайдалануға болады. Бөлу дәстүрлі әдістері өте қымбат металдар, ол тек өте бай металл кенін пайдалануға ұсынылады. Сондықтан, дамыту саласында дәстүрлі әдістерін пайдалана отырды, кейін әлеуетті өнім көп болды.
2. бактериялық шаймалау әдісімен металды алсаңыз, терең дамытусыз істеуге болады. Бірінші жарылғыш айып пайдаланып тастарды қиыршық, және ол содан кейін шаймалау ерітіндісін айдалады. Еритін металл тұздары бар ерітіндісін шаймалау аяқталғаннан кейін, жартастан бұрғыланған ұңғымадан айдалады. Бұл әдіс аз қымбат; ол пайдаланған кезде айтарлықтай бетінің рок үлкен көлемін төзбейді кезде, терең дамыту сияқты, қоршаған ортаны құнсызданған және қалдықтардың тауға қалыптаса қой2ған жоқ.
3. кеннен мыс өндіру дәстүрлі әдістері жоғары температураға талап етеді. Бұл әдістер, қымбат қазба отын тұтынады, демек, мысалы, қышқыл жаңбыр тудырады ауаны ластайтын,. (бактериялық шаймалау күкірт қосылыстарының шаймалау әдісімен қазба отын түрлерін емдеу үшін болашақта пайдаланылуы мүмкін.)
4.Тау-кен қалдықтарын бақылаусыз шаймалау ауыр металдармен жақын сулардың ластануына әкелді. Бұл бақыланатын шаймалау және металл қалпына келтіру болдырмауға болады. Ол батыс Құрама Штаттарында ғана тау-кен үйінділерде мыс астам 33 млн. Тонна деп бағаланып отыр. Әдетте жазықтарға орналастырылған қадалар, және, осылайша, металдар алыс қашықтықтарға өзен су асырылуы мүмкiн. Сілтілі бар суда еритін металл даунстрим бөгеті кезде жинайды және металдарды өндірумен айналысады зауытта, айдалады болады. Қажет болса, тазартылған су үйіндісіне үшін қайта өңдеуге болады.
5.Гендік инженерия арқылы, атап айтқанда, Т. ferrooxidanss жылы, бактериялар жақсарту үшін әрекеттер қолданылады.
12.16. Ферменттік технология
Бұл бөлімде жасушадан бөлінген ферменттердің биотехнологияда қолданылуын қарастырамыз.
Ферменттер бұлар жасушалардағы химиялық белсенділікті реттеуге мүмкіндік беретін биология катализаторлар. Адамзат 1000 жылдан бері ферменттерді қолдануына қарамастан, тек 19 ғасыр соңында олардың қалай жұмыс жасайтынын түсініп бастады. Қазір біз білеміз, ферменттер дегеніміз спецификалық 3 өлшемді конфигурация бар күрделі белоктық молекулалар. Және құрылымы ДНК да кодталған. Мүмкін атын конфигурацияның саны есепсіз көп болуы мүмкін.
Өнеркәсіп үшін ферменнттер екі негізгі себептерге байланысты тартымды. Біріншіден, әртектілігінің арқасында ферменттер көптеген өндірістік химиялық реакцияларды катализдеуге потенциалды қабілетті. Екіншіден, олар әдете қолданылатын органикалық емес катализаторлардан әлдеқайда тиімді және ерекше. Қалыпты температура мен қалыпты қысым кезінде олардың қатысуымен жүзеге асатын дәл сондай реакциялар әдетте жоғары температураны, жоғары қысымды талап етеді. Мысалы, Хабер тәсіліне негізделген әлемдегі ең үлкен өнеркәсіп бірінде, аммиак (NH3) газ түрунде азот және сутекте 500 °С температура және жоғары қысымда алынады.
Азотфиксациялаушы бактериялар аммиакты атмосферадағы азот және сутектен бөлме температурасында және қалыпты атмосфералық қысымда АТФ-ті энергия көзі ретінде, ферменттердің көмегімен синтездеуге қабілетті. Егер ферменттердің көмегімен аммония алу технология өңдеу етудің сәті түссе, онда өте кқп энергияны үнемдеуге болар еді. Ферменттердің тағы бір артықшылығы, өте таза өнім беретінінде-олардың специфаукостісі.
Бұл болса, фариокология, тамақ және ауылшаруашылық өнеркәсіп үшін өте маңызды.
Ферменттерді қолданудың бірқатар кемшіліктері, негізінен жасушадан бөлінген белоктардың тұрақсыздығына байланысты. Мұндай белоктар темпаратура және рҺ өзгергенде және өндірістік процесте қолданылатын органикалық еріткіштердің әрекеті кезінде тез денатурацияланады. Олар реакция өнімдерімен де түтік арқылы қызмет етуі мүмкін. Ферменттер өздерінің бөліну процестерінің жоғары құны да кемшілік болып саналады. Және бұл үшін «қауіпсіз» (мысалы, залалсыздар) организмді пайдалану қажеттігі, әсіріесе бұл ферменттер жануарлар мен адамдарға қажет өнімдерді алу үшін арналған болса, күні бүгін 2500 жазылған, суреттелген (өз кезегінде олар табиғатта табылғандардың жалпы санының тек 10 %құрайды). Ферменттердің 200ден азы қолданылады. Ферменттердің көпшілігі бар болғаны саңырауқұлақтардың 11 түрінен, ашытқының 4 түрінен және бактерияның 8 түрінен шығып отыр.
Болашақта жаңа ферменттерді құрастыру үшін кең мүмкіндік ашылады. Олардың біреуі – ферменттер кодтайтын гендерді өзгерту жолымен кейбір аминқышқылдарды бағытты түрде өзгерту. Белоктардың спецификалық үш өлшемді конфигурация алу заңын біз терең біліп отырмыз. Сондықтан жақын болашақта толық жаңа құрастырылған ферменттердің жарыққа шығуы теріске шығарылмаған. Бұл бағыт белоктық инженерия деп аталады. Бұдан басқа да, біз ферменттердің қалай жұмыс жасайтынын көбірек білеміз. Оның үстіне, мүмкін, біз әдеттегі ферменттерге қарағанда әлдеқайда тұрақты белоктық емес немесе ішінара белоктық емес катализаторды құрастыра аламыз. Мүмкін, дәл о сы бағыт кәсіпкерлер үшін тартымды, ұнамды болады.
Құны тез қайтарылатын бағыттардың ішінде – қазір қолданыстағы ферменттерге толық альтернативті болатын табиғи ферменттерді іздеу. Қазір ферменттерді, өндірістік көптеген формадағы компаниялар; сыра,пиво, шарап жасаушылар детергент, текстил, жеміс шырынын өндірушілер және тағы басқалар қолдануда. Ферменттерді әлемдік сату деңгейі күні бүгін, жылына 1 млрд доллардан асып кетті. 12.6 кестеде ферментті қолданудың кейбір мәліметтері берілген.
12.16.1. Ферменттердің көзі (бұлағы)
Микроорганизмдердің өсімдік және жануарлармен салыстырғанда ферменттердің көзі ретінде, жоғарырақ тұруы келесі себептерге :
- Олар өсудің өте жоғары жылдамдығын алуы.
- Оларды ферменттерде бақыланатын жағдайда, бұл экономикалық тиімде көп мөлшерде алуға болады.
- Олар химиялық реакциялардың кең спектрлерлік жүзеге асады.
- Олардың қасиетін гендік инженерияның тәсілдер көмегімен немесе әртірлі бейнедегі мутаттарды алу жолымен оңай жеңіл жаңартуға болады.
- Олардың азықтық, қоректік нәрселерден керектілеулері өте қарапайым.
- Олар өте арзан субстраттарда, қалдықтарда көбінесе өсуге бейімделген.
- Өнімнің жылдамдығын қолдануға көре өзгертуге болады. Олар жеңіл жиналатын, тазаланатын көптеген жасушалардан тыс ферменттер өнім береді.
Ферменттерді әдетте микрооргнаизмдерден алады. Мынадай сұрақтуындауы мүмкін?
Алдын – ала ферменттерді шығарғаннан гөрі ферменттерлерді тұтас жасушаларды пайдалану мықты емес пе?
Егер өнімді алу үшін тек бір химиялық реакция керек болады және бұған көре бір фермент керек болады. Олай болса, бұл процесс жасушада болып жатқан процеске ұқсас. Тазаланған ферменттерді пайдалану тұтас микроорганизмдерді пайдаланудан артық болуының себептері:
- Микроорганизмдерді өсіру үшін фермертерлерді өсіру және қолдану – қымбатқа түсетін іс.
- Ферментацияны стирильді жағдаййда жүргізу керек.
- Ферменттерде тұтас жасушалар аэроция және қозғалту кеззінде көп энергия қажет пайдаланады.
- Ферментерлерде өнімдер лас түрде болады, қалдықтарды шығару керек, бұл болса қоршаған ортаның ластануына әкеліп соғады. Тазаланған ферменттердің көмегімен жүзеге асырылып жатқан өндірістік процестерде «реакторлардың»ерекше кончтрукциясы қолданылады. Оны суретке жазу бұл кітаптың мәселесіне кірмейді. Алайда, олар ферментерден әлдеқайда қарапайым.
Егер ферментті микроорганизммен алу керек болса, бұл микроорганизмді үлкен көлемде алдымен, фенһрментерде өсіріп алады. Қоректік көзі ретінде глюкощаны қолданбауға тырысады. Себеі ол көптеген пайдалы ферменттердің синтезін басып кетеді. Көбіне үщдіксіз ферментациялауды пайдаланады. Тек глюкозамераздарды шығаруына мүнің қатысы жоқ. Көптеген ферменттерден өздігінен жасушалардан бөлінеді. Сөйтіп олар, жасушадан тыс фермент болып саналады. Оларды ішкі жасушаға қарағанда тазалауөте оңай . оларғ алдымен, сұйық ортадан бөліп алып, әртүрлі жолдармен араластырып, құрамын өзгертеді.содан соң өнімді керегінше тазалайды. Кейбір ферменттерді толық тазалау міндетті емес. Ферменттерді алынған түрінде пайдалануға болады. Мысалы, шырындарды дайындағанда, кір жуатын түйіршіктерді пайдаланғанда соалй жасайды. 12.17.7 бөліменде қараңыз.
12.16.4. Жеміс шырындарын дайындау
Ири алдымен өнімді ұсаұқтайды. Басқа –өсмдіктердей жеміс жасушаларынын қабырғасы айналган жасушалардан тұрады. Бұл қабырғаларында целлюлоза, полисахаритер, гемицелюлозалар болады. Қабырға жасушалары өте мықты, оларды бұзу қиын. Өнімнен сөлін сапалы үрде алу үшін целлюлозамен гемицеллюлоза қосады. Ферменттерді таңдағанда,төменгі рН температурасында жұмыс жасай алатындай қылып жинақтайды, себебі жеміс шырындары қышқыл болады. рНтың оптикалық көрсеткіші 4 пен 5ті құрайды.өсім жасушалары күрделі полисахаридтердің әсерімен ұстап тұра алады. Оларды пектин дейді. Олардың құрамында жасбысқыш зат болады. Сол себепті, ол джемнің кебуіне әкеледі. Жемістерді сақтағанда немесе ұсақтағанда олар суды ерітетін пектинге айналады. Сондықтан олар жеміс шырындарында болады. Төменгі температурада пектиндер колоидті суспензия бөліп шығарып, шырыннан бөлінеді. Бұл шырынға лайлы түр береді. Кейбір Америка, Англия елдері үшін шырынның бұл түрі жақпайды.ал негізінде жемісті шырынның ең жоғарғы көрсеткіші.егер шырынға пектиназ ферменттерін қосса, олар пектинді полисахаридке езеді. Және шырында өте төменгі температурада сақталалды. Сөйтіп, пектиназдар шырынды сұйық қылады.
12.26. бөлімінде қараңыз. Пектиназ көзі болып бактериялар саналады. Осындай жолмен сыра мен шарап та дайындалады. Ал бұлардың лайлығы ақуыздағы танниннің болуыннан болады. Сырадағы лайдың төмен болуы үшін, салқындатқанда протеиназды қосады. Бұларға пепсин мен попоин жатады.
12.16.5. Етті жұмсарту
Ет бұл негізгі бұлшық ақуызы. Бұлшықтар түйілген – түйілген ақуызды волоконды??? құрайды. жалғастыратын терісі де структуралық ақуызды құрайды. Сонымен бұлар кологен мен илластин. Кологен қатты созылмайтын, ал иллостин созылмалы сары волокондар. Етті шайнауға ыңғайлы қылып, бір сөзбен айтқанда оны езе отырып, ақуызбен қосылған терісін ұсақтайды. Бұл кезде волокна қысқарады. Және бір – бірінен тез айырылады. Ақуызды бұзатфын ферменттерді протеиназдар деп атайды. Етті жұмсарту үшін көбіне попеин қолданылады. Оны попоя өсімдігінің шырынынан алады. Бұл фермент қосып езілген еттер үй жағдайында сатылымда кездесед. Қолданар бұрын етті белгілі бір уақытқа өзінің шырынына қояды. Бұл кезде ферменттер ө жұмысын жасайды. Кейбір елдерде попоинді малды сояр алдында қанына жібереді. Сөйтіп, сойылған малдың еті өте жұмсақ болады. Осы сияқты етті езі процестері өлімнен кейін жүреді. Өлген жасушалар лизосомдар деп атайды. Өлген жасушаларда лизосомалар бұзылады. Олардан ферменттер бөлініп шығып, бұзылу жұмысы басталады. Бұл процесс автолиз деп аталады.
5.10.6. бөлімдерін қараңыз
Сондықтан малдың ет туштарын қолданар алдында бірнеше күнге салқын қоймаларда бірнеше күнге қояды.
12.16.5. Етті жұмсарту
Ет бұл негізгі бұлшық ақуызы. Бұлшықтар түйілген – түйілген ақуызды талшықты құрайды. жалғастыратын терісі де структуралық ақуызды құрайды. Сонымен бұлар кологен мен илластин. Кологен қатты созылмайтын, ал иллостин созылмалы сары волокондар. Етті шайнауға ыңғайлы қылып, бір сөзбен айтқанда оны езе отырып, ақуызбен қосылған терісін ұсақтайды. Бұл кезде волокна қысқарады. Және бір – бірінен тез айырылады. Ақуызды бұзатфын ферменттерді протеиназдар деп атайды. Етті жұмсарту үшін көбіне попеин қолданылады. Оны попоя өсімдігінің шырынынан алады. Бұл фермент қосып езілген еттер үй жағдайында сатылымда кездесед. Қолданар бұрын етті белгілі бір уақытқа өзінің шырынына қояды. Бұл кезде ферменттер ө жұмысын жасайды. Кейбір елдерде попоинді малды сояр алдында қанына жібереді. Сөйтіп, сойылған малдың еті өте жұмсақ болады. Осы сияқты етті езі процестері өлімнен кейін жүреді. Өлген жасушалар лизосомдар деп атайды. Өлген жасушаларда лизосомалар бұзылады. Олардан ферменттер бөлініп шығып, бұзылу жұмысы басталады. Бұл процесс автолиз деп аталады. 5.10.6. бөлімдерін қараңыз
Сондықтан малдың ет туштарын қолданар алдында бірнеше күнге салқын қоймаларда бірнеше күнге қояды.
12.16.6. Биологиялық қоспалар қосылған жуғыш түйіршіктер
Алғашқыбиологиялық қоспалар қосылған кір жуғыш түйіршіктер 1960 жылдарждың ортасында паййды болды. Олардың құрамында езгіщ ақуыздар протеиназдар болды. Олар өз орнында дақтарды кетіру үшін қолданылды. Мысалы, қанның дағы, шөптің дағы, жұмыртқа дағы. Уыз бұзылған кезе дақтар кетеді. Кір жуу жұмысы өте төменгі температуларда жасауға болатфн болды. Бұл дегеніміз электр ток күшін үнемдеуге пайдасын тигізді. Жуғыш түйіршіктегі протеиназдар рН 9,10 жағдайында жұмыс жасай алды. Және оларды ң құрамында көптеген мөлшерде фосфатттар болды. Олар қосымша жуу және шаю үшін керек. Болды. Батыс Еуропада кір жуғыш материалдардың қолданудың барысында 25 пайыз электр қуаты үнемделді. Кір жуғыш түйіршіктерді пайдаланғаннан бастап, кейбір фабрика жұмысшылараның арасында денсаулықтарына зиян келе бастады. Олардың кейбіреулеріне түйіршікке деген аллергия басталды. Ұсақ түйіршіктерді демалыс кезінде жұтып қоятын болатындықтан тамақтары қышып, демалыстары қиындады. Сөйтіп, 1970 жылдары АҚШта жуғыш түйіршіктер сатылымнан алды. Ал еуропада қолданыла берді. Жоғарыдағы проблемаларды болыдармау үшін, жуғыш түйіршіктердің құрамын өзгертті. Оларға инертті воск??? Қосты. Сонымен ұқатар кір жуғыштар сұйық түрінде шығарыла бастады. Қазіргі таңда кір жуғыш түйіршіктер АҚШта қайтадан пайда бола бастады. 1990 жылы олар сатылымда 15 пайызды құрады. Ал Батыс Еуропа 85 пайызын құрады.
Сурет 12.27. биологиялық қосымшалар қосылған жуғыш түйіршіктерді кішкентай суреті. Кейбір бөліктерінде ішінде орналасқан ферменттер көріндеді.
Ет бұл негізгі бұлшық ақуызы. Бұлшықтар түйілген – түйілген ақуызды талшықты құрайды. жалғастыратын терісі де структуралық ақуызды құрайды. Сонымен бұлар кологен мен илластин. Кологен қатты созылмайтын, ал иллостин созылмалы сары волокондар. Етті шайнауға ыңғайлы қылып, бір сөзбен айтқанда оны езе отырып, ақуызбен қосылған терісін ұсақтайды. Бұл кезде волокна қысқарады. Және бір – бірінен тез айырылады. Ақуызды бұзатфын ферменттерді протеиназдар деп атайды. Етті жұмсарту үшін көбіне попеин қолданылады. Оны попоя өсімдігінің шырынынан алады. Бұл фермент қосып езілген еттер үй жағдайында сатылымда кездесед. Қолданар бұрын етті белгілі бір уақытқа өзінің шырынына қояды. Бұл кезде ферменттер ө жұмысын жасайды. Кейбір елдерде попоинді малды сояр алдында қанына жібереді. Сөйтіп, сойылған малдың еті өте жұмсақ болады. Осы сияқты етті езі процестері өлімнен кейін жүреді. Өлген жасушалар лизосомдар деп атайды. Өлген жасушаларда лизосомалар бұзылады. Олардан ферменттер бөлініп шығып, бұзылу жұмысы басталады. Бұл процесс автолиз деп аталады. 5.10.6. бөлімдерін қараңыз
Сондықтан малдың ет туштарын қолданар алдында бірнеше күнге салқын қоймаларда бірнеше күнге қояды.
12.16.6. Биологиялық қоспалар қосылған жуғыш түйіршіктер
Алғашқыбиологиялық қоспалар қосылған кір жуғыш түйіршіктер 1960 жылдарждың ортасында паййды болды. Олардың құрамында езгіщ ақуыздар протеиназдар болды. Олар өз орнында дақтарды кетіру үшін қолданылды. Мысалы, қанның дағы, шөптің дағы, жұмыртқа дағы. Уыз бұзылған кезе дақтар кетеді. Кір жуу жұмысы өте төменгі температуларда жасауға болатфн болды. Бұл дегеніміз электр ток күшін үнемдеуге пайдасын тигізді. Жуғыш түйіршіктегі протеиназдар рН 9,10 жағдайында жұмыс жасай алды. Және оларды ң құрамында көптеген мөлшерде фосфатттар болды. Олар қосымша жуу және шаю үшін керек. Болды. Батыс Еуропада кір жуғыш материалдардың қолданудың барысында 25 пайыз электр қуаты үнемделді. Кір жуғыш түйіршіктерді пайдаланғаннан бастап, кейбір фабрика жұмысшылараның арасында денсаулықтарына зиян келе бастады. Олардың кейбіреулеріне түйіршікке деген аллергия басталды. Ұсақ түйіршіктерді демалыс кезінде жұтып қоятын болатындықтан тамақтары қышып, демалыстары қиындады. Сөйтіп, 1970 жылдары АҚШта жуғыш түйіршіктер сатылымнан алды. Ал еуропада қолданыла берді. Жоғарыдағы проблемаларды болыдармау үшін, жуғыш түйіршіктердің құрамын өзгертті. Оларға инертті воск??? Қосты. Сонымен ұқатар кір жуғыштар сұйық түрінде шығарыла бастады. Қазіргі таңда кір жуғыш түйіршіктер АҚШта қайтадан пайда бола бастады. 1990 жылы олар сатылымда 15 пайызды құрады. Ал Батыс Еуропа 85 пайызын құрады.
Сурет 12.27. биологиялық қосымшалар қосылған жуғыш түйіршіктерді кішкентай суреті. Кейбір бөліктерінде ішінде орналасқан ферменттер көріндеді.

Жуғыш түйіршіктердің сапасы жоғары болу үшін оларға жаңа ферменттер қосты. Целлюлоза микроволокноларды езіп, бүлінген мақта волокнолорынан айырады. Сөйтіп, киімді одан да жарқын және жұмсақ қылып шығаратын болды. Сонымен бірге, ферменттерге оттегін езетін амилазаны қосты. Бұлар крахмалды езеді. Кездескен кедергілердің бірі протеиназдар болды. Олар басқа ферменттерді бұзып жіберіп отырды. Жақында саңырауқұлақтардың бірінде соған ұқсас липаза табылды. Ол орташа температурада белсенді болды. Оның кодтың генін гендік инженерия әдістері арқылы басқа саіырауқұлаққа көшірілді. Липаздар липидтерді езеді. Бұл дегеніміз май дақтары, жақпа заттардың дақтарын кетіреді. Әдетте көбіне көпіршік түйіршіктері үшін бакцеллюз бактериялары үшін пайдаланады. Бұан шығарылған ферменттер жасушадан тыс фермен болып саналады. Сөйтіп , тазалау жұмысы орналасады.
12.16.7. Иммибилизденген ферменттер
Иммибилизденген ферменттердің пайдасы
Жоғары бөлімде айтылғандай коммерциялық ферменттерді қолдану кейбір факторларға әкеліпс оқтарыд. Олардың бастысы ферменттердің тұрақсыздығы және бағасының қымбаттығы. Бағасын ферменттердің иммобилизациялау арқылы түсіруге болады. Бұл дегеніміз ферментті қасық бетіне немесе ішіне бекітеді. Кейін ферментация біткен соң алынып тасталынады. Бұл фермент жұмыс барысының құнын түсіру үшін қолданылады. Иммубилизацияның және бір жұмысы ферменттер рН температурасының өзгеруінен тұрақты болуы. Мысалы, иммобилизленген глюкоозо иззамераз 65 градусқа бір жылдай шыдамды. Ал сұйықтықта 45 градусда бірнеше сағатқаа шыдамды. Иммобилизденген ферментті ашық шығарылым үшін жасалынған ферменттердің соңында қосады.
Ферменттерді Иммобилизациялау әдісі
Ферменттерді Иммобилизациялаудың түрлі әдісі бар
12.28 суретін қара.
Бұған механиклық қосылу немес матрица жатады. Механикалық түрінің пайдасы фермент негізгі түрінде сақталынады. Бірақ, ірі молекулаларға ферментке жету қиындық туғызады. Альгенат шарларымен лабараториялық шауруаларда мұны оңай көрсетіге болады. Бұл кеңінен қолданылатын әдіс. Альгенат пен натрий ферменттері қосылған сұйықтықты тамшылатып, хлолы кальцийге тамызады. орлы натрий мен әсерлескеннен кейін олар гельге айналады. Сөйтіп, шарик түрінде гельдер пайда болады. Ұзақ пайдаланау үшін гельді полиакремагламидтерменм тұрақтандырады.
Иммобиозденген ферменттердің қолданылуы
Иммобилизденген ферменттерді ең жақсы жұмысы болып, жоғарғы фруктозаның жоғары құрамының жүгері сорпасжол қосылған жұмысын айтамыз. Бұл Жапонияда, АҚШта тәттілеу жұмысында орын алады. Мысалы, жеміс шырындарында сорпаны арзан көміртеккөзінен крахмалдан, дайындайды. Бұл процесс 3 ферменттің қатысуымен болады. Алдымен, крахмал массасын жүгеріні қыру арқылы алады. Солан соң екі амилазды крахмал глюкозды сорпасына айналдырады. Қоюлатылған сұйық сорпаны неше түрлі тағамдармен ішімдіктерге қосады. Глюкозомераз ферментінің көмегімен бұл сорпаны глюкоза мен фруктзаны қоспасы бар сорпаға айналдыруға болады. Бұл үшін сорпаны колонкадан өткізеді. Мұнда иммобилизденген фермент адсорбция жолымен целлюлозаны қосады.жақұа 12.3
Ферменттердің белсенділігі уақыт өте төмендейді. Сондықтан бұл үшін бірқалыптажұмыс жасап тұрған бірнеше колонканы алады. Фруктоза колориясы мен салмағы бірдей болғанымен глюкозадан тәтті. Бұл дегеніміз жүгері сорпасы қосылған жоғары құрамдағы фруктозаны пайдалана отырып, колориясы аз көлемдегітәтті тағам алуға болады. Жыл сайын АҚШта 4 млн тонна сироп шығарылады. Алғашқы иммобилизденген фермент болып, аминоапилаз саналды. Ол 1969 жылы Жапонияда малға қосылатын жем ретінде қолданды. Әдемдік сатылымда бұл өнімүлкен сұранысқа ие.
Гельмен ұстау. Мысалы, селекагельмен, альгинат (теңіз балдырынан алынған полисахарид).
Микрокапсуламен ұстау
300нм диматерге дейін өткізгіш микрокапсулалар
Оң жақ
Осындай жолмен фермент адам денесіне немесе жануар денесіне енгізілген фермент иммундық системаның әсерінен қорғанады.
Адсорбция бұл өте арзан, ферменттің денаутурасысына әкеліп соқтырмайды. Фермент жағдай өзгерген кезде оңай айырылады. Мысалы, рН иондық күшінде
Астына
Әлсіз әсерлердің мысалы, гидрофобтық әсерлердің жұмысы. Бұлар глюкозаизморезаны иммобилациялауда қолданады. Негізі адцорб сапасы үшін ион алмасу смаласы қолданады.және гидрофобты смала қолданады. Қосылғаннан кейін ферментті глутаральдигидтік ігумен мықты байланыстырады. Адсорбтанған ферменттер мембран ферменттерімен байланысты.
4 ковалентті байлану
Оң жақ матрикс целлюлоза мен кологееннен құрылған полимер
Сол жақ. Матрикспен коваленттік байланыстыру .
Оң жақ түбігне. Бұл қымбат әдіс. Бірақ өнім ластанған кезде ферменттер шығарылмайды. Кейбір ферменттер байланысуын жоғалтады.
12.28. Иммобилизациялау әдісі
Әрбір аминқышқыл молекуласы екі формада өмір сүреді. Біреуі екіншісінің айнадағыдай мүсіні. Мысалы оң қол мен солұол сияқты. Бұл екі форма оптикалық иззамен деп аталады. Және оң және сол айналатын форма болып, саналады. Немесе Д және Л формасы бардық табиғатта кездемсетін аминқылқылдар, Л аминқышқылдар болып саналады. Аминқылқылдарды химиялық синтезбен оңай алуға болады. Бірақ аминқышқылдар синтезделген кезде екі бірдей көлемде Д және Л лизомерін құрайды. Л аминқышқылын алу проблемасы ферменттер арқылы шешіледі. Жұмыс барысының негізгі түрі 12.29. диаграммасында көрсетілген. Ферментті иондық байланыстцыру арқылы колонкіде иммобилиздейді.
Әдіс 3
Автоматты түрдегі тоқтаусыз жұмыстан кейін 50 градустық 20 кун 50 градустық температурадығы блсендіі ферменттер 40 пайызға төмендейді. Олапрдың орнын толтыру үшін жаңа ферменттер қосады. Иммобилизация барысында 40 пайыз ферментті үнемдейді. Иммобилизденген ферменттерді қолданудың басқа мысалы жартылай синтетикалық шығарылымлар. Пенцилиндердің табиғи пеницилиннен шығарылуы иммобилизденген фермент химиялық түрден пеницилиннің бір жарты тобын модификсириялайды . бұл пеницилннің антибиологиялық белсенділігін күшейтеді.
12.17 . биосенсорлар
Биосенсорлар электрондық тіркелген құралдардан жасалынады. Бұлар химиялық заттардың қоблығын тексеру үшін қолданылады. Мысалы, жасушаны ферментті және антиденені оның көмегімен әсіресе ферменттпен антидененің өлшеу ыңғайлы. Өйткені , олар қиын қосындыдағы молекулаларды ажырата алады. Биологиялық материалдар мен субстраттар арасындағы биологиялық реакциялар арқылы электрлік сигналын алынады. Бұл жануарлар органдарының сезімталдылығын аулау үшін қажет. Электр сигналының санау үшін мысалы, цифр таблосында дауыс көбейткіш керке. Биосенсор неше түррдегі типтегі мысалы, жылу бөлуді, жарықты, немесе массасын электр жүйесін жаңа химиялық қосындыларды ұстауға қабілетті.
Сол жақ Л аминқышқыл оң жақ Д аминқышықлы
Қоспа
Ацел топтарының қосындысы
Аминациклаз ферменттерінің қосындысы
Ортаға амина аксилаз ферменті ацил тобын жояды.
Оң жақ Л және Д аминқышқылдары химиялық араласып, қайтадан пайдалынады.
М аминқыщқылдары Дациламинқылықылы
Л аминқышылы Даминқылына қарағарда өзгеше ерігіш
Бөлунген аминқышщылы
12.29. малдарға жем дайындау үшін аминқышқылдың қосылуы. Сурет
12.30
Өнім қосылғанда өнмі өзгертеді.
12.17.1. биосерсорларды пайдаланудың пайдасы мен зияны
Негізгі пайдасы
Олар мықты емес, сондықтан мұқият емес. *
12.31 суретін қараңыз
Биосенсор иммобилизденген түрдегі глюкозоксидорті ферментті құрайды. Фермент қандағы глюкозаны ашытады.со кезд электо тоғын тудыратын элекрондар шығарылады.
Бұлар қандағы глюкозаның санына пропорционалды болады. Биосенсор өтесезімтал ол 1 тамшы қандағы глюкоза құрамының 20 с – да анықтай алады. Болашақта сусақ ауруының қан тамырына инсулинді қадағалап отыратын құралды қондырмақ . егер биосенсорды минанососпен жалғастырса ол автоматты түрде инсулин бөліп, ауру дер кезінде көмек алады. Мұндай бақылаулар аурулардың көзінің бөтекесінің бөлінуінен қорғайды.
12.17.2. медицинада қолдану
Қазіргі уақытта медицинада кеңінен қолданылады: адамның биологиялық сұйықтығындағы болған гормондар,және метоболит құрамдарын, дәрілерді тез айыру үшін керекті құрал. Бұл әсіресе клиникалық диагностикада қадет. Биосенсорларды пайдаланубарысында қатте диагноз қою азаюда. Биосенсор көмегімен дәрігер терапеф кабинетінен шықпай, анализдерін біледі. Сонымен қатар ауруларғочиптер а диагнозын білу үшін қайта келу қажеті тумайды. Емдеуу тез басталады. Адам қанындағы анаша қолданған адамды қанынан тез айырады. Осыған қарағанда алдағы уақытта үй жағдайында тексеруге болатфын аппараттар шығады. Қазіргі кезде ота жасау барысында қауіпті метаболиті табу үшін биосенсорлар пайдалынылады. Биочиптерді пайдалану барысында сезімтал биосенсорлардың кішкене мөлшерлерін ойлап табуда. Дәл осылай селекондық микрочиптерді пайдалана отырып, компьюетр өлшемлерінің кішіреюін аламыз . биосипетер адам денесіне енгізідетінндей кішкентай болуы қажет. Сонда мысалы, жасанды түрде жүрек қағысын қадағалау сияқты жұмыстары жемісті болады.
12.17.4. Басқа салада қолданылуы
Биосенсорларда басқа салада қолдану, мысалы, Шик ізат процесстерінде. Тірі жасушалар (ашытқы мен бактерий) электроддтармен қосылып, аминқышқыдың , спирттің, финолдың ментанның, уксустың антибиотиктерін, қоблықтарын өлшеуге қолданады. Оның көмегімен ферменттер ішіндегі жағдайды бақылайды. Бұл өз жолында дәндә дақылдарды тексеру үшін қажет. Оттектің сапасының анықтайтын сезімтал системалар ойлап табылды. Бұл системада бактериялар оттекпен қосылғанда жарқырай бастайды. Жарқыраған жарық фотоэлементтермен тіркеледі. Үлкен жетістіктерге оптикаық биосенсорлар жетті. Көптеген ферменттер жылу бөлумен жалғасады. Термо биосенсорлар температураны 0-0,01ге дейін өзгерісін тексереді. Оларды сүт қышқылын табу үшін қолдануға болады. Сурет 12.30.
Болашақта биосенсорлар ауыл шаруашылығында ветеранияда адам ортасын сақтау үшін (нервпоролистік газдарды анықтауға, )және қоршаған ортаны(алдымен ластанудан қорғану үшін) пайдалаады. Бұл сферадағы биосенсордларды пайдалану жыл сайын 30 пайызға өсуде.
12.16.7. Иммибилизденген ферменттер
Иммибилизденген ферменттердің пайдасы
Жоғары бөлімде айтылғандай коммерциялық ферменттерді қолдану кейбір факторларға әкеліпс оқтарыд. Олардың бастысы ферменттердің тұрақсыздығы және бағасының қымбаттығы. Бағасын ферменттердің иммобилизациялау арқылы түсіруге болады. Бұл дегеніміз ферментті қасық бетіне немесе ішіне бекітеді. Кейін ферментация біткен соң алынып тасталынады. Бұл фермент жұмыс барысының құнын түсіру үшін қолданылады. Иммубилизацияның және бір жұмысы ферменттер рН температурасының өзгеруінен тұрақты болуы. Мысалы, иммобилизленген глюкоозо иззамераз 65 градусқа бір жылдай шыдамды. Ал сұйықтықта 45 градусда бірнеше сағатқаа шыдамды. Иммобилизденген ферментті ашық шығарылым үшін жасалынған ферменттердің соңында қосады.
Ферменттерді Иммобилизациялау әдісі
Ферменттерді Иммобилизациялаудың түрлі әдісі бар
12.28 суретін қара.
Бұған механиклық қосылу немес матрица жатады. Механикалық түрінің пайдасы фермент негізгі түрінде сақталынады. Бірақ, ірі молекулаларға ферментке жету қиындық туғызады. Альгенат шарларымен лабараториялық шауруаларда мұны оңай көрсетіге болады. Бұл кеңінен қолданылатын әдіс. Альгенат пен натрий ферменттері қосылған сұйықтықты тамшылатып, хлолы кальцийге тамызады. орлы натрий мен әсерлескеннен кейін олар гельге айналады. Сөйтіп, шарик түрінде гельдер пайда болады. Ұзақ пайдаланау үшін гельді полиакремагламидтерменм тұрақтандырады.
Иммобиозденген ферменттердің қолданылуы
Иммобилизденген ферменттерді ең жақсы жұмысы болып, жоғарғы фруктозаның жоғары құрамының жүгері сорпасжол қосылған жұмысын айтамыз. Бұл Жапонияда, АҚШта тәттілеу жұмысында орын алады. Мысалы, жеміс шырындарында сорпаны арзан көміртеккөзінен крахмалдан, дайындайды. Бұл процесс 3 ферменттің қатысуымен болады. Алдымен, крахмал массасын жүгеріні қыру арқылы алады. Солан соң екі амилазды крахмал глюкозды сорпасына айналдырады. Қоюлатылған сұйық сорпаны неше түрлі тағамдармен ішімдіктерге қосады. Глюкозомераз ферментінің көмегімен бұл сорпаны глюкоза мен фруктзаны қоспасы бар сорпаға айналдыруға болады. Бұл үшін сорпаны колонкадан өткізеді. Мұнда иммобилизденген фермент адсорбция жолымен целлюлозаны қосады.жақұа 12.3
Ферменттердің белсенділігі уақыт өте төмендейді. Сондықтан бұл үшін бірқалыптажұмыс жасап тұрған бірнеше колонканы алады. Фруктоза колориясы мен салмағы бірдей болғанымен глюкозадан тәтті. Бұл дегеніміз жүгері сорпасы қосылған жоғары құрамдағы фруктозаны пайдалана отырып, колориясы аз көлемдегітәтті тағам алуға болады. Жыл сайын АҚШта 4 млн тонна сироп шығарылады. Алғашқы иммобилизденген фермент болып, аминоапилаз саналды. Ол 1969 жылы Жапонияда малға қосылатын жем ретінде қолданды. Әдемдік сатылымда бұл өнімүлкен сұранысқа ие.
Гельмен ұстау. Мысалы, селекагельмен, альгинат (теңіз балдырынан алынған полисахарид).
Микрокапсуламен ұстау
300нм диматерге дейін өткізгіш микрокапсулалар
Оң жақ
Осындай жолмен фермент адам денесіне немесе жануар денесіне енгізілген фермент иммундық системаның әсерінен қорғанады.
Адсорбция бұл өте арзан, ферменттің денаутурасысына әкеліп соқтырмайды. Фермент жағдай өзгерген кезде оңай айырылады. Мысалы, рН иондық күшінде
Астына
Әлсіз әсерлердің мысалы, гидрофобтық әсерлердің жұмысы. Бұлар глюкозаизморезаны иммобилациялауда қолданады. Негізі адцорб сапасы үшін ион алмасу смаласы қолданады.және гидрофобты смала қолданады. Қосылғаннан кейін ферментті глутаральдигидтік ігумен мықты байланыстырады. Адсорбтанған ферменттер мембран ферменттерімен байланысты.
4 ковалентті байлану
Оң жақ матрикс целлюлоза мен кологееннен құрылған полимер
Сол жақ. Матрикспен коваленттік байланыстыру .
Оң жақ түбігне. Бұл қымбат әдіс. Бірақ өнім ластанған кезде ферменттер шығарылмайды. Кейбір ферменттер байланысуын жоғалтады.
12.28. Иммобилизациялау әдісі
Әрбір аминқышқыл молекуласы екі формада өмір сүреді. Біреуі екіншісінің айнадағыдай мүсіні. Мысалы оң қол мен солұол сияқты. Бұл екі форма оптикалық иззамен деп аталады. Және оң және сол айналатын форма болып, саналады. Немесе Д және Л формасы бардық табиғатта кездемсетін аминқылқылдар, Л аминқышқылдар болып саналады. Аминқылқылдарды химиялық синтезбен оңай алуға болады. Бірақ аминқышқылдар синтезделген кезде екі бірдей көлемде Д және Л лизомерін құрайды. Л аминқышқылын алу проблемасы ферменттер арқылы шешіледі. Жұмыс барысының негізгі түрі 12.29. диаграммасында көрсетілген. Ферментті иондық байланыстцыру арқылы колонкіде иммобилиздейді.
Әдіс 3
Автоматты түрдегі тоқтаусыз жұмыстан кейін 50 градустық 20 кун 50 градустық температурадығы блсендіі ферменттер 40 пайызға төмендейді. Олапрдың орнын толтыру үшін жаңа ферменттер қосады. Иммобилизация барысында 40 пайыз ферментті үнемдейді. Иммобилизденген ферменттерді қолданудың басқа мысалы жартылай синтетикалық шығарылымлар. Пенцилиндердің табиғи пеницилиннен шығарылуы иммобилизденген фермент химиялық түрден пеницилиннің бір жарты тобын модификсириялайды . бұл пеницилннің антибиологиялық белсенділігін күшейтеді.
12.17 . биосенсорлар
Биосенсорлар электрондық тіркелген құралдардан жасалынады. Бұлар химиялық заттардың қоблығын тексеру үшін қолданылады. Мысалы, жасушаны ферментті және антиденені оның көмегімен әсіресе ферменттпен антидененің өлшеу ыңғайлы. Өйткені , олар қиын қосындыдағы молекулаларды ажырата алады. Биологиялық материалдар мен субстраттар арасындағы биологиялық реакциялар арқылы электрлік сигналын алынады. Бұл жануарлар органдарының сезімталдылығын аулау үшін қажет. Электр сигналының санау үшін мысалы, цифр таблосында дауыс көбейткіш керке. Биосенсор неше түррдегі типтегі мысалы, жылу бөлуді, жарықты, немесе массасын электр жүйесін жаңа химиялық қосындыларды ұстауға қабілетті.
Сол жақ Л аминқышқыл оң жақ Д аминқышықлы
Қоспа
Ацел топтарының қосындысы
Аминациклаз ферменттерінің қосындысы
Ортаға амина аксилаз ферменті ацил тобын жояды.
Оң жақ Л және Д аминқышқылдары химиялық араласып, қайтадан пайдалынады.
М аминқыщқылдары Дациламинқылықылы
Л аминқышылы Даминқылына қарағарда өзгеше ерігіш
Бөлунген аминқышщылы
12.29. малдарға жем дайындау үшін аминқышқылдың қосылуы. Сурет
12.30
Өнім қосылғанда өнмі өзгертеді.
12.17.1. биосерсорларды пайдаланудың пайдасы мен зияны
Негізгі пайдасы
- Биосенсор специфисті – химиялық заттардың қиын қосындыларын қосымша тазартусыз айыра алады.
- *олар өте сезімтал, сондықтан өте төменгіқоблықта көріге болады.
- Тез жауап береді
- Қолдануы қауіпсіз
- Олар өте дәл
- Олар өте кішкентай
- Көптеген шығарымға пайдалануға болады.
Олар мықты емес, сондықтан мұқият емес. *
- Олар тұрақты емес , оларды тазалауға болмайды.
- 12.171. қандағы глюкозаның құрамын бақылау
12.31 суретін қараңыз
Биосенсор иммобилизденген түрдегі глюкозоксидорті ферментті құрайды. Фермент қандағы глюкозаны ашытады.со кезд электо тоғын тудыратын элекрондар шығарылады.
Бұлар қандағы глюкозаның санына пропорционалды болады. Биосенсор өтесезімтал ол 1 тамшы қандағы глюкоза құрамының 20 с – да анықтай алады. Болашақта сусақ ауруының қан тамырына инсулинді қадағалап отыратын құралды қондырмақ . егер биосенсорды минанососпен жалғастырса ол автоматты түрде инсулин бөліп, ауру дер кезінде көмек алады. Мұндай бақылаулар аурулардың көзінің бөтекесінің бөлінуінен қорғайды.
12.17.2. медицинада қолдану
Қазіргі уақытта медицинада кеңінен қолданылады: адамның биологиялық сұйықтығындағы болған гормондар,және метоболит құрамдарын, дәрілерді тез айыру үшін керекті құрал. Бұл әсіресе клиникалық диагностикада қадет. Биосенсорларды пайдаланубарысында қатте диагноз қою азаюда. Биосенсор көмегімен дәрігер терапеф кабинетінен шықпай, анализдерін біледі. Сонымен қатар ауруларғочиптер а диагнозын білу үшін қайта келу қажеті тумайды. Емдеуу тез басталады. Адам қанындағы анаша қолданған адамды қанынан тез айырады. Осыған қарағанда алдағы уақытта үй жағдайында тексеруге болатфын аппараттар шығады. Қазіргі кезде ота жасау барысында қауіпті метаболиті табу үшін биосенсорлар пайдалынылады. Биочиптерді пайдалану барысында сезімтал биосенсорлардың кішкене мөлшерлерін ойлап табуда. Дәл осылай селекондық микрочиптерді пайдалана отырып, компьюетр өлшемлерінің кішіреюін аламыз . биосипетер адам денесіне енгізідетінндей кішкентай болуы қажет. Сонда мысалы, жасанды түрде жүрек қағысын қадағалау сияқты жұмыстары жемісті болады.
12.17.4. Басқа салада қолданылуы
Биосенсорларда басқа салада қолдану, мысалы, Шик ізат процесстерінде. Тірі жасушалар (ашытқы мен бактерий) электроддтармен қосылып, аминқышқыдың , спирттің, финолдың ментанның, уксустың антибиотиктерін, қоблықтарын өлшеуге қолданады. Оның көмегімен ферменттер ішіндегі жағдайды бақылайды. Бұл өз жолында дәндә дақылдарды тексеру үшін қажет. Оттектің сапасының анықтайтын сезімтал системалар ойлап табылды. Бұл системада бактериялар оттекпен қосылғанда жарқырай бастайды. Жарқыраған жарық фотоэлементтермен тіркеледі. Үлкен жетістіктерге оптикаық биосенсорлар жетті. Көптеген ферменттер жылу бөлумен жалғасады. Термо биосенсорлар температураны 0-0,01ге дейін өзгерісін тексереді. Оларды сүт қышқылын табу үшін қолдануға болады. Сурет 12.30.
Болашақта биосенсорлар ауыл шаруашылығында ветеранияда адам ортасын сақтау үшін (нервпоролистік газдарды анықтауға, )және қоршаған ортаны(алдымен ластанудан қорғану үшін) пайдалаады. Бұл сферадағы биосенсордларды пайдалану жыл сайын 30 пайызға өсуде.
