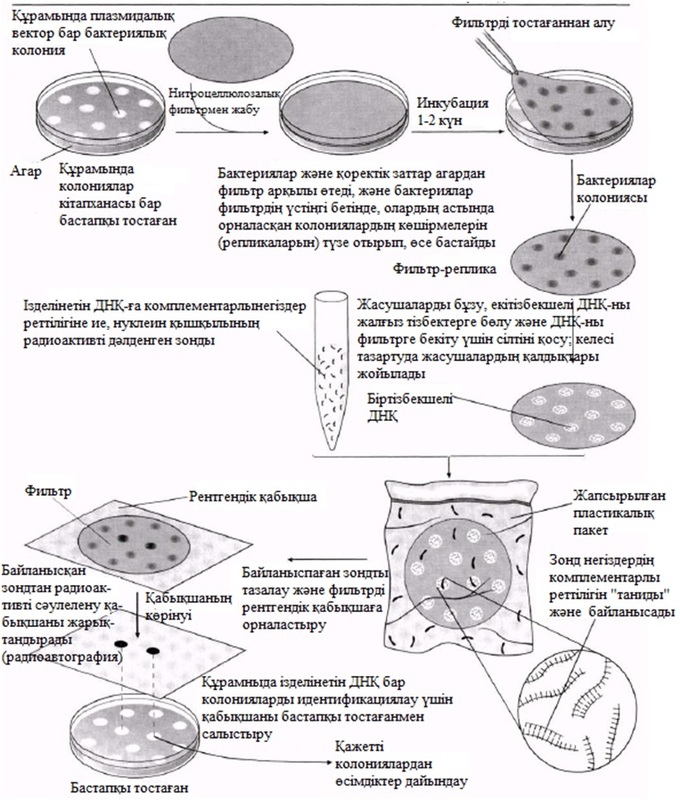
23
ӨМІРДІҢ ҮЗДІКСІЗДІГІ
| 23_тарау_Камшат.pptx |
5- бөлімде атап өтілгендей, биологияның ең басты ережелерінің бірі- барлық тірі организмдердің құрылымы мен функциясының негізгі бірлігі ретінде жасуша концепциясы.
Ең алғаш жасуша теориясы 1838 жылы Шлейденмен және 1839 жылы Шваннмен тұжырымдалған. Рудольф Вирхов жаңа жасушалардың, бұрынғы жасушалардың бөлінуі нәтижесінде пайда болатынын 1855 жылы жариялау арқылы жасуша теориясын кеңейтті. Өмірдің үздіксіздігінің мойымдалуы ХІХ ғасырдың екінші жартысында басқа ғалымдардың жасушаның құрылымы мен жасушаның бөліну механизміне зерттеу жүргізуіне түрткі болды. Гистологиялық тәсілдерді жетілдіру және жоғары үлкейту мүмкіндігі бар микроскоптардың жасалуы, ядроның маңызды рөлін және ондағы жасалған хромосомалардың, жасушалардың келесі ұрпақтар арасындағы сабақтастығын қамтамасыз ететін құрылымын анықтауға мүмкіндік берді. 1879 жылы Бовери және Флемминг, нәтижесінде екі бірдей жасуша пайда болатын ядродағы болған құбылысты сипаттады, ал 1887 жылы Вейсман гаметалардың, белгілі бір ерекше түрдің бөлінуі нәтижесінде пайда болатындығы туралы ой білдірді. Бұл бөлінудің екі түрі тиісінше митоз және мейоз атауларына ие. Олар туралы оқымастан бұрын, хромосомалармен жақынырақ танысқан пайдалы.
23.1. Хромосомалар
23.1.1. Хромосомалар және кариотүрлер
Жасушалардағы бөлінудің ең маңызды рөлін тұқымқуалаушылық ақпараттарды ұрпақтан-ұрпаққа таратылуын қамтамасыз ететін хромосомалар атқарады. Бұл тасымалдау хромосомадағы ДНҚ молекуласының болуына байланысты жүзеге асады. Ядроның бөліну кезеңінде әр хромосома құрамында бір ДНҚ молекуласы болады. Бөлінуден бұрын бұл ДНҚ молекуласының қатарында оның дәл көшірмесі пайда болады, сондықтан хромосома екі бірдей ДНҚ молекулаларынан тұратын жұптық құрылымға айналады. Бұл екі хромосома бөлігі хроматидтер деп аталады. Әр хроматидте екі бірдей ДНҚ молекуласының біреуі болады.
Интерфаза деп аталатын бөліну арасындағы кезеңде хромосомаларды құрайтын материал негізгі кейбір бояулармен боялатындықтан хроматин деп аталса да, хромосомалар жарық микроскоптарымен қарағанда жекеленген құрылымдар тәрізді айыруға тұрмайды. Осы даму барысында хромосомалар ұзын, жіңішке жіптер түйінін құрайды. Ядроны бөлу алдында олар қысқа, жуан және бір-біріне дағдыланған, ерекше ықшам және тез түсін өзгертетін құрылымға айналады. 23.1. суретінде метафаза дамуы барысындағы адамның жасушадағы хромосомаларының фотосуреті көрсетілген.
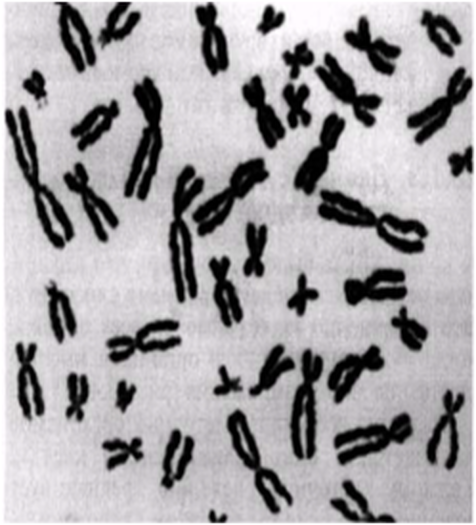
23.1. сур. Адамның хромосома жиытығының фотосуреті (ер адамда). Әр хромосома центромера деп аталатын, нүктеде байланысқан екі хроматидтен тұрады. Жиынға 46 хромосома кіреді. Олардың әр-түрлі пішініне және центромерлердің әр-түрлі орналасуына назар аударыңыз
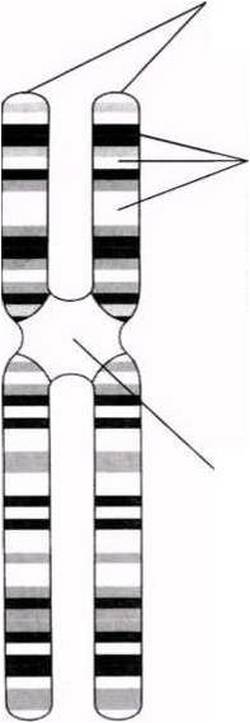
Екі бірдей хроматидтер- біреуі екіншісінің көшірмесі. Әрқайсысында бір ДНҚ молекуласы болады.
Әр-түрлі белгілірді анықтайтын гендер.Әр ген белгілі бір хромосомада, белгілі бір орын немесе локус алады. Мысалы, фиброздық-кистозды дегенерация гені 7 хромосомада локализденген.
Центромера-хроматидтерді байланыстыратын аймақ. Бұл аймақ боялмайды.
Әр-түрлі белгілірді анықтайтын гендер.Әр ген белгілі бір хромосомада, белгілі бір орын немесе локус алады. Мысалы, фиброздық-кистозды дегенерация гені 7 хромосомада локализденген.
Центромера-хроматидтерді байланыстыратын аймақ. Бұл аймақ боялмайды.
23.2. сур. Хромосома құрылымының оңайлатылған сызбасы. Негізінде, хромосомадағы гендер саны бірнеше жүзден бірнеше мыңға дейін жетеді. Гендердің өлшемдері де осы сияқты өзгеріп отырады.
Бұл жерден әр хромосоманың екі хроматидтен тұратынын көре аламыз. Хроматидтер хромосоманың ұзындығының кез-келген жерінде, бір-бірімен центромера деп аталатын нүктеде бірігеді(23.2. сур.).
Организмнің әр жасушасы берілген түрге сәйкес белгілі хромосома санын құрайды. Адамда, 23.1. суретінде көретініміздей, 46 хромосома бар. Әр түрлерде хромосома саны қатты өзгеріп тұрады. Мысалы, шыбында 8 хромосома болса, ал Lysandra атты кішкентай, Испанияда кездесетін көбелекте, ол 380-ге жетеді. Мысықтарда 38 хромосома, ал иттерде 78 хромосома болады. Көптеген түрлерде әр жасушасында 12-ден 50-ге дейін хромосомалар болады. Тұқымқуалаушылық бірлігі-гендер-хромосомаларға ұзына бойы орналасқан (23.2. сур.). Адамда әр-түрлі гендердің саны, шамамен, 100 000-ға жетеді (соңғы көрсеткіштерге байланысты, олардың саны азырақ).
Егер 23.1. суретіндегідей, хромосомаларды фотосуреттен қиып алсақ және олардың өлшемімен және пішінімен сәйкестендіріп қойсақ, жасуша, шындығына келгенде хромосома жұптарынан тұратыны анықталады; оларды гомологиялық хромосомалар деп атайды, себебі олар бірдей құрылымды. 23.3. суретінде хромосомалар гомологиялық жұптарымен орналасқан. Мұндай фотосуретті кариограмма деп атайды, ал бүкіл хромосома жиынтығын-кариотүр деп атайды. 23.3. суретінде 23 жұп хромосома суреттелген. Жұп хромосомалардың бар болуы, адамның бір хромосома жиынтығын анасының жұмыртқа жасушасынан, ал басқасын әкесінен спермий арқылы алатынымен түсіндіріледі. Спермий мен жұмыртқа жасушасы қосылуы барысындағы ұрықтану процесінде екі хромосома жиынтығы бар зигота пайда болады.
Организмнің әр жасушасы берілген түрге сәйкес белгілі хромосома санын құрайды. Адамда, 23.1. суретінде көретініміздей, 46 хромосома бар. Әр түрлерде хромосома саны қатты өзгеріп тұрады. Мысалы, шыбында 8 хромосома болса, ал Lysandra атты кішкентай, Испанияда кездесетін көбелекте, ол 380-ге жетеді. Мысықтарда 38 хромосома, ал иттерде 78 хромосома болады. Көптеген түрлерде әр жасушасында 12-ден 50-ге дейін хромосомалар болады. Тұқымқуалаушылық бірлігі-гендер-хромосомаларға ұзына бойы орналасқан (23.2. сур.). Адамда әр-түрлі гендердің саны, шамамен, 100 000-ға жетеді (соңғы көрсеткіштерге байланысты, олардың саны азырақ).
Егер 23.1. суретіндегідей, хромосомаларды фотосуреттен қиып алсақ және олардың өлшемімен және пішінімен сәйкестендіріп қойсақ, жасуша, шындығына келгенде хромосома жұптарынан тұратыны анықталады; оларды гомологиялық хромосомалар деп атайды, себебі олар бірдей құрылымды. 23.3. суретінде хромосомалар гомологиялық жұптарымен орналасқан. Мұндай фотосуретті кариограмма деп атайды, ал бүкіл хромосома жиынтығын-кариотүр деп атайды. 23.3. суретінде 23 жұп хромосома суреттелген. Жұп хромосомалардың бар болуы, адамның бір хромосома жиынтығын анасының жұмыртқа жасушасынан, ал басқасын әкесінен спермий арқылы алатынымен түсіндіріледі. Спермий мен жұмыртқа жасушасы қосылуы барысындағы ұрықтану процесінде екі хромосома жиынтығы бар зигота пайда болады.

23.3. сур. 23.1. суреті негізінде жасалған, адам кариограммасы(ер адамдікі). Өлшемдері бойынша топтарға бөлінген аутосомалар(жыныссыз хромосомалар). Жыныстық хромосомалар бөлек топқа белгілінген Х-аналық, Y-аталық.Адам жасушасы 22 жұп аутосома және бір жұп жыныстық хромосомалардан тұрады. Аутосомалар құрамындағы гендерді, аутосомды деп, ал жыныстық хромосомалар құрамындағы гендерді, жыныстық байланысты деп атайды.
23.3. суретіндегі көрсетілген хромосомалар арасынан, бірдей емес хромосомалардан тұратын, Х және Y әріптерімен белгіленген бір жұпты көруге болады. Бұл жыныс хромосомалары. Аталық немесе Y-хромосома, аналық хромосомаларынан немесе Х хромосомаларынан қысқалау, онда аналық хромосомада болатын кейбір гендер жетіспейді (24.6. бөлім). Гомологиялық хромосомалар қалыпты жағдайда, бірдей белгілерді анықтайтын гендерден тұрады. 23.3 суретінде көрсетілген кариотүрі бар индивидуум—аталық (XY). Аналық хромосомасында екі Х- хромосомалары болады (ХХ). Кариограммаларда кейде хромосомалардың мутациясын көруге болады.
23.1.2. Гаплоидтық және диплоидтық жасушалар
Екі хромосома жиынтығынан тұратын жасуша түрлерін диплоидты деп атайды және 2n белгісімен көрсетеді. Көптеген үлкен жануарлар және өсімдіктердің жартысы диплоидты, яғни екі хромосома жиынтығынан тұрады. Кейбір сол тәріздес ағзалар тек бір хромосома жиынтығынан тұрады, және оларды гаплоидты деп атайды (белгісі n\ мысалға, 2 тараудағы ұрпақтардың алмасуы бөлімін қараңыз). Одан басқа, барлық гаметалар гаплоидты. Кейбір ағзалар, соның ішінде көптеген өсімдіктер, үш немесе одан да көп хромосома жиынтықтарынан тұрады, мұндай ағзаларды полиплоидты деп атайды, алайда бұл бөлімде біз олар туралы қозғамаймыз.
Екі жұп хромосомалардың бар болуы екі мүмкіндік береді:
1) қанша дегенмен, әр индивидуум екі ата-ананың да белгілерін алатын болғандықтан, генетикалық өзгергіштік өседі;
2) егер қандай да бір ген толық емес болса, екі хромосоманың біреуіндегі гомологиялық хромосомада болатын ген бұл кемшіліктің орнын толтыра алады.
23.1.3. Ядроның бөлінуінің екі тәсілі не үшін керек?
ХІХ ғасырдың соңында, ядроны бөлу екі түрлі тәсілмен жүзеге асатындығы анықталды. Бұл, ағзаның өмірлік циклінде жыныстық көбею кезеңі болған кезде, кез-келген жағдайда өте қажет (23.4. сур.).
Барлық көпжасушалы ағзалардың дамуы, бір ғана жасушадан басталады. Әр жасушаның бөлінуінің алдында, ядро бөлінеді. Егер де бұдан ядродағы хромосома саны әр кезде екі есе азаятын болса, онда жақын арада әр жасушада өте аз хромосома қалар еді. Еншілес жасушалардағы хромосомалар саны, ата-анасының хромосома санымен бірдей, сондықтан берілген ағзадағы барлық жасушаларда бірдей мөлшерде хромосомалар сақталады. Бұл митоз атымен белгілі бөліну әсерінен жеткізіледі.
Бірақ, 23.4. суретінде көрсетілгендей, жыныстық көбею қатысатын өмірлік жүйеде, зигота аталық және аналық гаметаларының қосылуы нәтижесінде пайда болады. Егер де бұл жасушалар екі хромосомадан құралатын болса, онда зиготада және барлық келесі жасушаларда, төрт хромосома жиынтығы болар еді, және болашақта олардың саны әр ұрпақ сайын екіге көбейетін еді.
23.1.2. Гаплоидтық және диплоидтық жасушалар
Екі хромосома жиынтығынан тұратын жасуша түрлерін диплоидты деп атайды және 2n белгісімен көрсетеді. Көптеген үлкен жануарлар және өсімдіктердің жартысы диплоидты, яғни екі хромосома жиынтығынан тұрады. Кейбір сол тәріздес ағзалар тек бір хромосома жиынтығынан тұрады, және оларды гаплоидты деп атайды (белгісі n\ мысалға, 2 тараудағы ұрпақтардың алмасуы бөлімін қараңыз). Одан басқа, барлық гаметалар гаплоидты. Кейбір ағзалар, соның ішінде көптеген өсімдіктер, үш немесе одан да көп хромосома жиынтықтарынан тұрады, мұндай ағзаларды полиплоидты деп атайды, алайда бұл бөлімде біз олар туралы қозғамаймыз.
Екі жұп хромосомалардың бар болуы екі мүмкіндік береді:
1) қанша дегенмен, әр индивидуум екі ата-ананың да белгілерін алатын болғандықтан, генетикалық өзгергіштік өседі;
2) егер қандай да бір ген толық емес болса, екі хромосоманың біреуіндегі гомологиялық хромосомада болатын ген бұл кемшіліктің орнын толтыра алады.
23.1.3. Ядроның бөлінуінің екі тәсілі не үшін керек?
ХІХ ғасырдың соңында, ядроны бөлу екі түрлі тәсілмен жүзеге асатындығы анықталды. Бұл, ағзаның өмірлік циклінде жыныстық көбею кезеңі болған кезде, кез-келген жағдайда өте қажет (23.4. сур.).
Барлық көпжасушалы ағзалардың дамуы, бір ғана жасушадан басталады. Әр жасушаның бөлінуінің алдында, ядро бөлінеді. Егер де бұдан ядродағы хромосома саны әр кезде екі есе азаятын болса, онда жақын арада әр жасушада өте аз хромосома қалар еді. Еншілес жасушалардағы хромосомалар саны, ата-анасының хромосома санымен бірдей, сондықтан берілген ағзадағы барлық жасушаларда бірдей мөлшерде хромосомалар сақталады. Бұл митоз атымен белгілі бөліну әсерінен жеткізіледі.
Бірақ, 23.4. суретінде көрсетілгендей, жыныстық көбею қатысатын өмірлік жүйеде, зигота аталық және аналық гаметаларының қосылуы нәтижесінде пайда болады. Егер де бұл жасушалар екі хромосомадан құралатын болса, онда зиготада және барлық келесі жасушаларда, төрт хромосома жиынтығы болар еді, және болашақта олардың саны әр ұрпақ сайын екіге көбейетін еді.
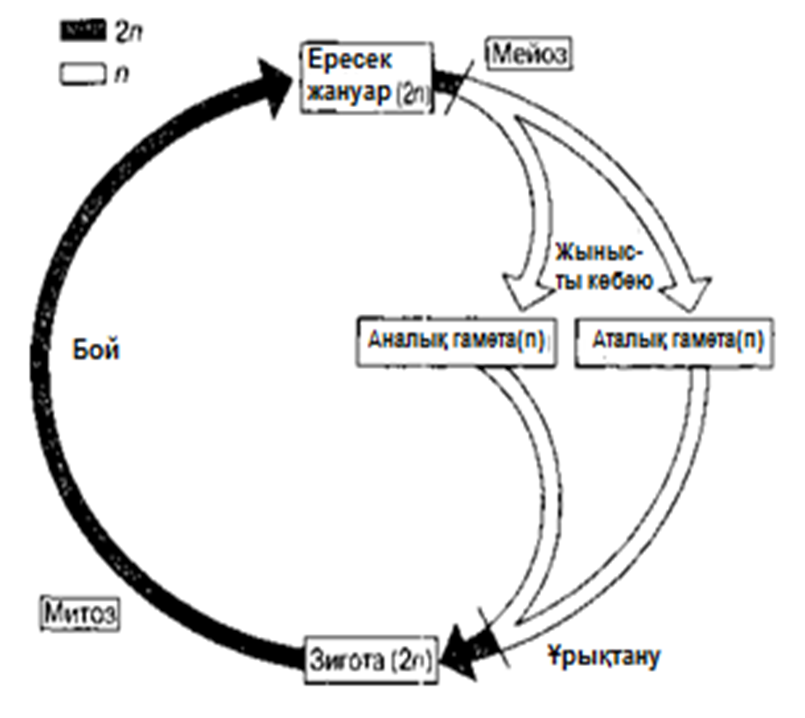
23.4. сур.Жануардың өмірлік циклінің схемасы
Сондықтан да қандай да бір өмірлік жүйе кезеңінде ядро, хромосома саны азаюымен бірге, басқа бір түрдің бөлінуіне шыдауы қажет. Алда, зигота түзілуі кезінде, диплоидтық жағдай қайта қалпына келеді. Еншілес жасушалардағы хромосома жиынтығының саны екіден бірге дейін азайғандағы ядроның бөлінуі, мейоз немесе редукциялық бөліну деп аталады.
23.1.4. Қысқаша қорытындылар
Митоз — бұл ата-аналық жасушалардың жиынтығымен бірдей хромосома жиынтығы бар екі еншілес ядро пайда болатын, жасуша ядросының бөлінуі. Әдетте, ядро бөлінген бойда, екі еншілес жасушалар түзілуімен барлық жасушаның бөлінуі жүреді. Митоз жасушаның бөлінуімен бірге, бой, регенерация және эукариоттардағы жасушалар алмастыру процесстерін қамтамасыз ете отырып жасуша санының көбеюіне әкеледі. Біржасушалы эукариоттарда, митоз, популяциялардың санының артуына әкелетін жыныссыз көбею механизмі ретінде қызмет атқарады.
Мейоз бастапқы ядроға қарағанда әрқайсысы екі есе аз хромосомалардан тұратын еншілес ядро түзілуінен жасуша ядросының бөліну процесін қамтамасыз етеді. Мейозды жасушадағы хромосомалар саны диплоидтыдан (2n) гаплоидтыға (n) дейін азаятын болғандықтан, редукциондық бөліну деп те атайды. Мейоздың мағынасы мынада, жынысты көбейетін түрлерде ол ұрпақтар қатарындағы тұрақты хромосома санын сақтауды қамтамасыз етеді. Мейоз жануарларда гаметаның және өсімдіктерде спораның түзілуінен пайда болады. Ұрықтану кезінде гаплоидтық гаметалардың қосылуы нәтижесінде диплоидтық хромосома саны қалыптасады.
23.2. Жасушалық цикл
Берілген жасушаның және оның еншілестерге бөлінуінің арасындағы оқиғалардың кезектесуі жасушалық цикл деп аталады. Бұл цикл үш басты кезеңнен тұрады.
1. Интерфаза. Синтез бен бой периоды. Жасушада, оның бойына және өзіне қатысты функцияларды орындауға қажетті көптеген заттар синтезделеді.
Интерфаза кезінде ДНҚ репликациясы жүреді.
2. Митоз. Бұл алда сипатталатын, ядроның бөліну процесі.
3. Жасушаның бөлінуі — цитоплазманың екі еншілес жасушалар арасынан бөліну процесі.
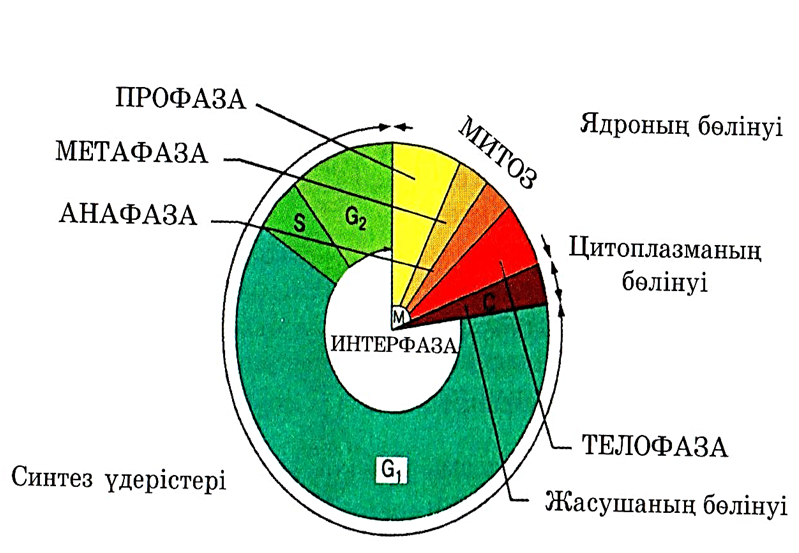

23.5. сур. Жасушалық цикл.
Барлық цикл 23.5. суретінде көрсетілген. Жасушалық цикл ұзақтығы түріне және температура, қоректік заттар және оттекпен қамтамасыз етілу сияқты сыртқы факторларға байланысты. Бактериялар әр 20 мин сайын бөліне алады, ішек эпителий жасушалары — әр 8-10 сағ, пияз тамырының ұшындағы жасушалары — әр 20 сағ, ал кейбір жүйке жүйесі жасушалары мүлдем бөлінбейді.
23.1. Тәжірибесі. Мацерирленген тамыр ұшындағы митоз фазаларын зерттеу
Әдетте хромосомаларды тек қана ядроның бөлінуі кезінде көруге болады. Бұл үшін сәйкес материал ретінде сарымсақ (2n=16) , пияздың (2n=16) және атбұршақтың (2n=12) тамыры ұшының апикальдық меристемасы қызмет атқарады. Бұл материалды, тамыршаның дамуын күшейтетін жағдайы бар жерге орналастырады; тамыршаның ұшын кеседі, тұрақтандырады, бояйды және ылғалдандырады (мацерация), осылардан кейін хромосомаларды микроскопта зерттеуге болады.
Материалдар және құрал-жабдықтар
Түйреуіштер
Суы бар шыны түтік
Скальпель
Тығыны бар кішкентай шыны түтік
Пинцет
Екі Петри табақшасы
Су моншасы және шыны түтік
Заттық шыны
Жабынды шыны
Жіңішке инелер жұбы
Фильтрлік қағаз
Сарымсақ бөлігі
Дистилденген су
Сірке қышқылы
Бір молярлық тұз қышқылы ерітіндісі
Фёлген реактиві
Тәсіл
1. Сарымсақ бөлігін түйреуішпен түйреңіз және бөлік негізі
суда болатындай шыны түтіктің үстіңгі бөлігіне іліңіз.
Кез келген бөтен әсерлер жасушаның бөлінуін
уақытша басып тастайтын болғандықтан, 3-4 күнге жалғыз
қалдырыңыз.
2.1-2 см ұзындықта бірнеше тамыршалар пайда
болғаннан кейін олардың бітетін бөлігін 1 см
ұзындықта кесіп алыңыз.
3.Кесілген тамырша бөліктерін сірке қышқылы бар кішілеу
шыны ыдысқа орналастырыңыз, оны тығынмен бітеңіз және
тұрақтану (фиксациялану) үшін бөлме температурасында
қалдырыңыз.
4.Тамыршаларды үстіңгі бітетін бөлігінен пинцетпен қысып,
дистильденген суы бар Петри табақшасына ауыстырыңыз
және фиксаторды жою үшін бірнеше минут ішінде жуыңыз.
5.Тамырша ұштарын бір молярлық тұз қышқылы ерітіндісі бар
шыны ыдысқа ауыстырыңыз және 60 ˚С-та 3 мин ұстаңыз
(пияз, асбұршақ немесе бұршақ тамыршаларына 6-10 мин).
Осы кезде жасушаларды өзара байланыстыратын орта
пластинкалар жойылады; ал ДНҚ хромосомалар бояуышпен
әрекеттесе алатын, дезоксирибозаның альдегидтік
формасының қалыптасуымен бірге гидролизденеді.
6.Тамырша ұштарымен бірге қышқылды дистильденген суы
бар Петри табақшасына құйыңыз және қышқылды жуыңыз. 5
мин-қа қалдырыңыз.
7.Тамыршаларды Фёльген реактиві бар кішкентай шыны
ыдысқа ауыстырыңыз және шыны ыдысты тығынмен
жабыңыз. Қараңғы, салқын жерге кем дегенде 2 сағ-қа
қойыңыз (мұздатқышқа салған дұрыс).
8.Бір ұшын тартып шығарыңыз және оны таза заттық шыны
үстіне сірке қышқылы тамшысына орналастырыңыз.
9.Бітетін бөлігін 1-2 мм ұзындықта кесіп алыңыз және қалған
бөлігін лақтырыңыз.
10.Тамыршаның ұшын екі жіңішке иненің көмегімен
шашақтаңыз және оны заттық шынымен жабыңыз.
Препаратты жазық бетке орналастырыңыз, бірнеше бет
фильтрлік қағазбен жабыңыз және оның үстінен затттық
шыныға үлкен саусақпен қатты басыңыз. Заттық шынының
бір жаққа қарай ығысуына жол бермеңіз. Бұл процедура
мацерация деп аталады.
11.Препаратты кішірек және үлкенірек ұлғайтатын
микроскоппен зерттеңіз және митоздың әр түрлі кезеңінде
орналасқан жасушаларды табыңыз.
12.Митоздың әр-түрлі кезеңінде орналасқан ядроны салыңыз
және суреттерді атап жазыңыз.
23.3. Митоз
Митоз кезіндегі ядрода болатын құбылыстарды, әдетте фиксирленген және боялған препараттардан бақылайды. Мұндай препараттар жасуша бөлінуі кезінде хромосомалардың өтетін фазаларын көруге мүмкіндік береді, бірақ олардың кезектесуін анықтамайды.
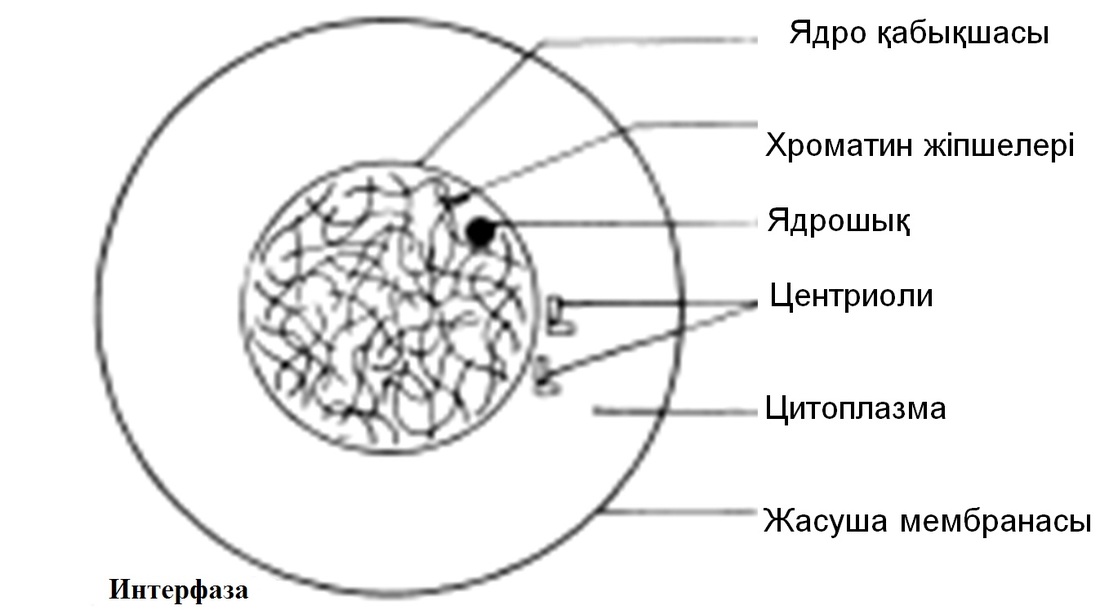
Интерфаза
Интерфаза ұзақтығы, берілген жасушаның функциясына байланысты әр-түрлі болып келеді. Ядро бөлінбестен бұрын, дереу әр ДНҚ хромосомалары көшірмеленеді. Нәтижесінде әр хромосома енді центромерада байланысқан, хроматид түрінде көрсетіледі. Бұл кезеңде жасуша әр ДНҚ молекуласының 4 көшірмесін (4n), гомологиялық жұптардың әр хромосомасында екі-екіден құрайды. Интерфаза кезінде, хромосомдық материал, оралған жіпшелерден тұратын борпылдақ масса күйінде болады, ол хроматин деп аталады. Центриольдар көшірмеленді.
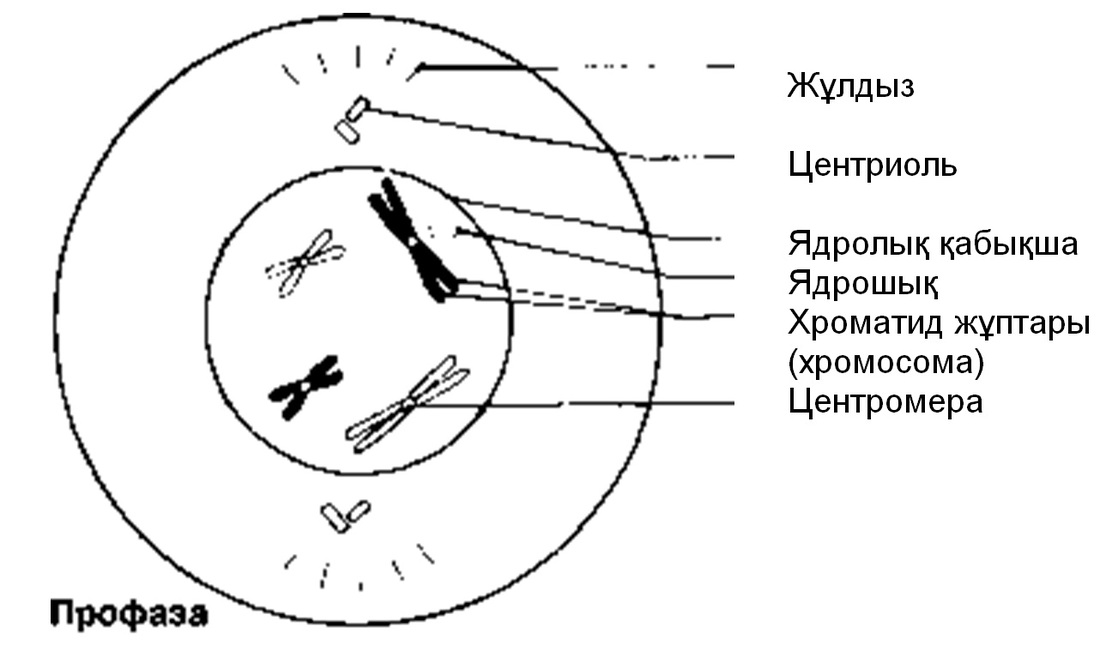
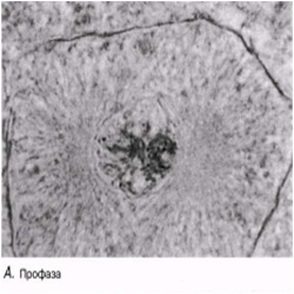
Профаза
Әдетте ең созылмалы бөліну фазасы. Хромосомалар қысқарады және спирализация және оның тығызырақ компоненттерінің нәтижесінде қалыңданады.Олар боялғанда анық көрінеді. Әр хромосома боялмайтын, центромерамен байланысатын екі хроматидтен тұрады. Жануарлар жасушасында центриоли жасушаның қарама-қарсы ұштарына таралады. Центриолиден радиусқа дейін созылып жатқан және жұлдызша құрайтын қысқа микротүтікшені көруге болады. Ядрошықтар, олардың ДНҚ-сы кейбір хромосомаларға ауысатындықтан, жойылады. Профазаның соңында ядро қабықшасы көптеген ұсақ көпіршіктер түзілуі арқылы дезинтегрирленеді (ыдырайды). Ұршық пайда болады.
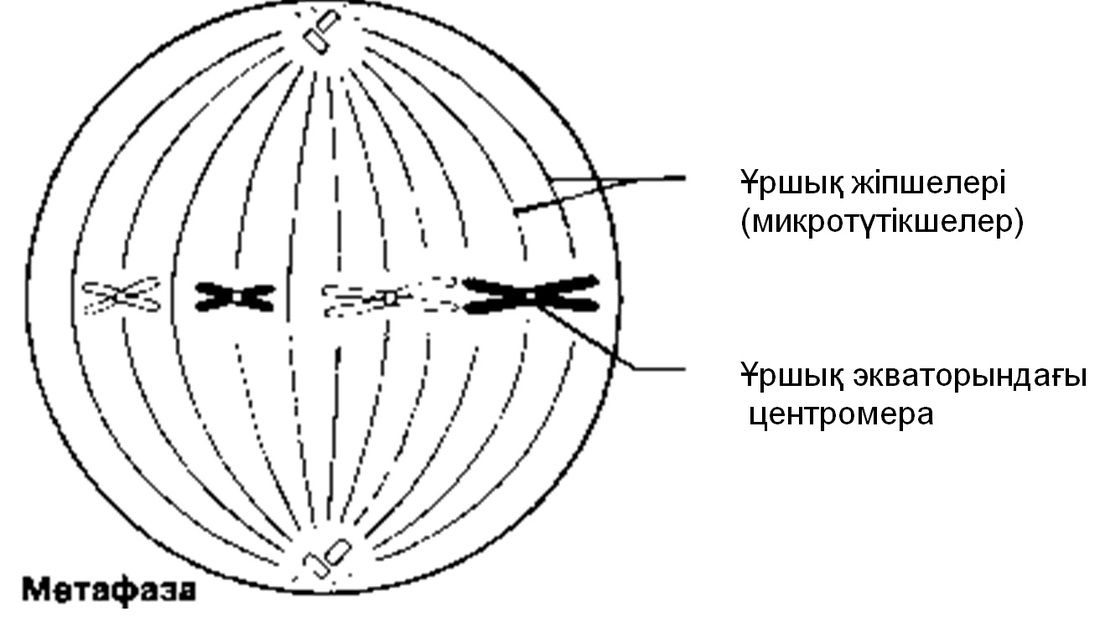
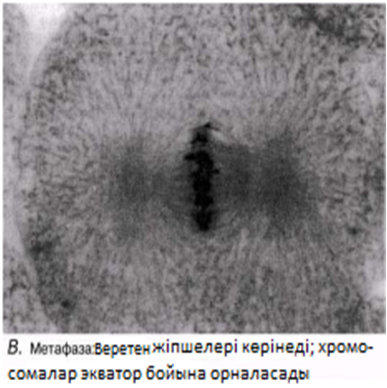
Метафаза
Хромосомалар жасушаның экватор жазығына орналасады. Олар ұршық жіпшелеріне (микротүтікше) өздерінің центромерасы арқылы біріккен.
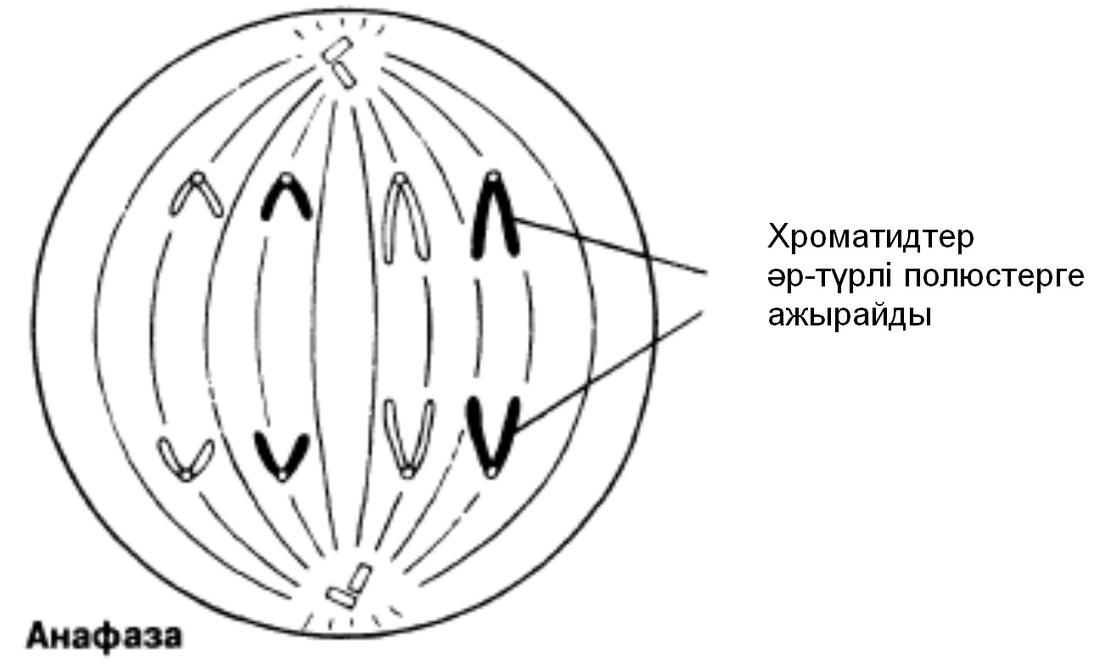
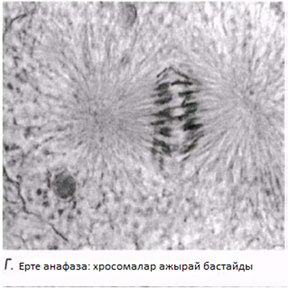
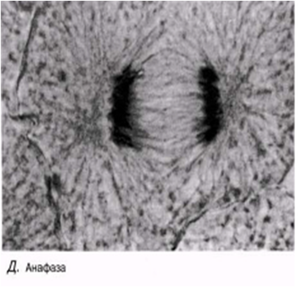
Анафаза
Бұл кезең өте тез жүреді. Центромералар екіге бөлшектенеді және ұршық жіпшелері еншілес центромерлерді қарама-қарсы полюстерге тартады. Центромералар өздерімен бірге бір-бірінен бөлінген хроматидтерді тартады.
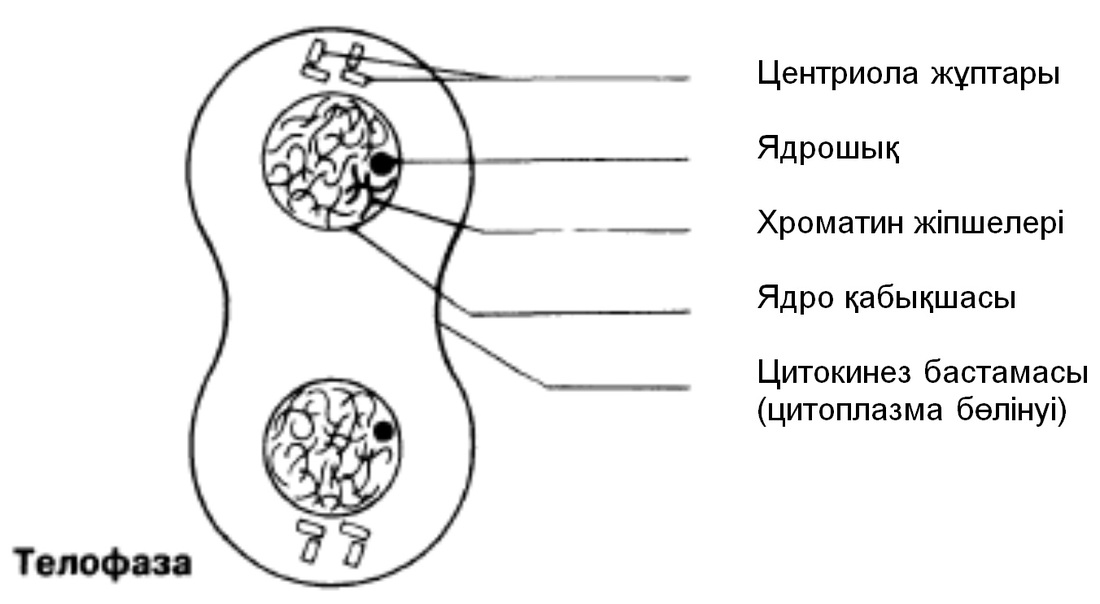
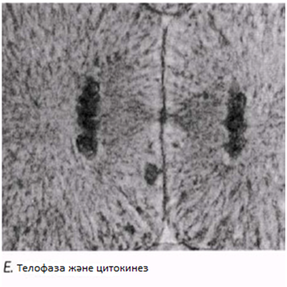
Телофаза
Хроматидтержасуша полюстеріне жетеді; айналады және созылады; олар қайтадан хроматинге айналады және нашар ажыратылған болады. Ұршық жіпшелері жойылады, ал центриоли көшірмеленеді. Әр полюстегі хромосома айналасында қайтадан ядро қабықшасы қалыптасады және ядрошық пайда болады. Телофазадан соң бірден цитокинез жүреді (жасушаның бөлінуі)
23.6. сур. Жануар жасушасындағы митоз
|

|
|
|
|
|
23.7. сур. Жануар жасушасындағы митоз және жасушалық бөліну кезеңдері.
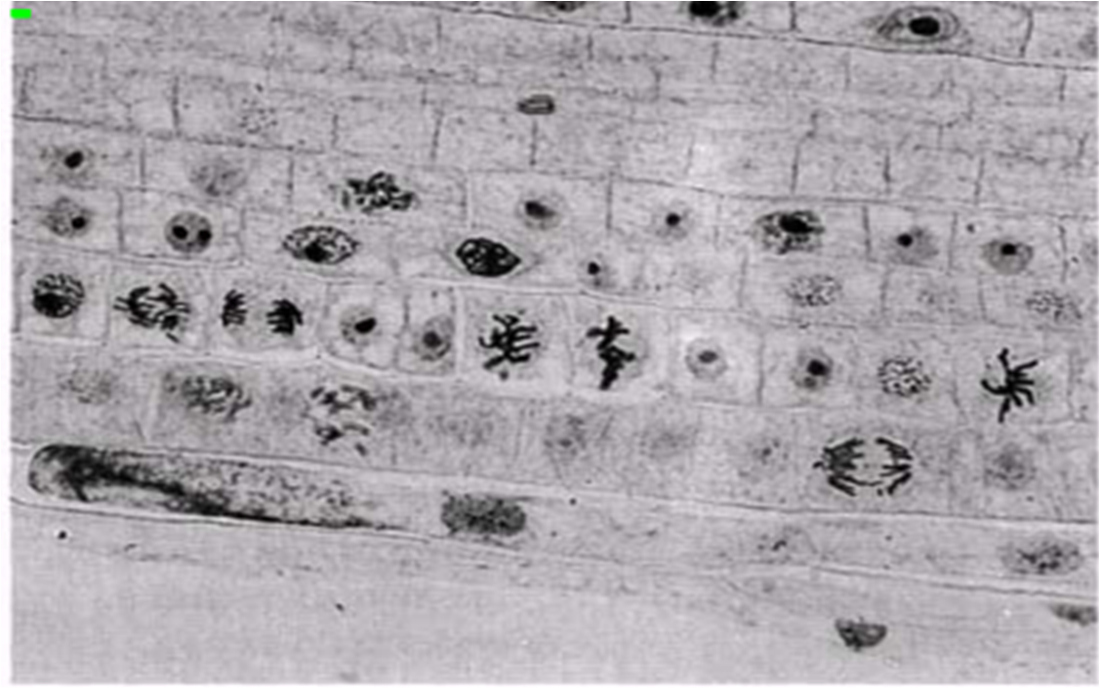
23.8. сур. Тамыр ұшының бойлық қимасы. Өсімдік жасушасына тән, митоз кезеңдері көрінеді. Бұл кезеңдерді 23.7. суретінде ұсынылған ақпараттар негізінде анықтауға тырысыңыз.
Бірақ, митоздың фазалар арасындағы кенет шектеулерсіз, үздіксіз процесс екендігін есте сақтау керек. Фазалық-контрасттық микроскопия және цейтраферлік түсірілім әдістері, майлы жасушадағы ядроның қалай бөлінетіндіктерін бақылауға мүмкіндік береді. Қабықтың тез айналуы кезінде митоз үздіксіз процесс ретінде қабылданады. Ыңғайлылық үшін оны 4 фазаға бөледі. Осы фазалар кезеңіндегі жануар жасушасындағы өзгерістер 23.6. суретінде көрсетілген. Жануар және өсімдік жасушаларындағы митоз фотосуреттері 23.7. және 23.8. суреттерінде көрсетілген.
23.3.1. Центриольдер және ұршықтың түзілуі
Центриольдер — бұл цитоплазмада ядро қабықшасының жанында орналасқан органеллалар; олар жануар жасушасында және төмен өсімдіктер жасушасында болады. Бұл бір-біріне перпендикуляр орналасқан жұп органеллалар.
Әр центриольдің ұзындығы шамамен 500 нм-ді құрайды, ал оның диаметрі 200 нм; олар әр топта үшеуден, тоғыз микротүтікше топтарынан тұрады. Көршілес үштік микротүтікше өзара фибриллалармен байанысқан деп саналады (23.9. сур.). Микротүтікше тубулин нәруызының суббірлігінен тұратын, диаметрі 25 нм болатын, ұзын қуыс түтіктерді құрайды (5.10.7. бөлімі).

23.9.А.сур.Центриольдің көлденең кесіндісінің тауық ұрығының ұйқы безі жасушасынан алынған электронды микрофотосуреті. Б. Центриольдің көлденең кесіндісінің сызба түріндегі суреті.
Центриоль әрқашан микротүтікшелердің дамуына жағдай туғызатын, анық белгіленген құрылымы жоқ материалда орналасады. Жасушаның осы аймағын центросома деп атайды. Центросома ғана ұршықты құрайды, себебі «ұршық жіпшелері» негізінде микротүтікшелерді құрайды. Бұл өсімдік ретінде центриоласы жоқ саңырауқұлақтардың да ұршық түзе алатындығын түсіндіреді. Ядро бөлінгендегі центриола функциясы анық емес болып қалады. Мүмкін олар ұршық бағытын таңдауға қатысып, осылайша жасуша бөлінуінің қай жазықтықта өтетінін анықтауға көмектеседі. Кейбір ұршық жіпшелері бір полюстен басқа полюске созылған кезде, басқалары полюстен центромераға созылады. Тубулин суббірліктерінің жойылуы нәтижесінде бұл ұршық жіпшелерінің қысқаруы, жасуша бөлінуі кезіндегі хромосомалар мен хроматидтердің орын ауыстыруын түсіндіреді. Олар центросомамен айқын «оралады».
Белсенді бөлінетін жасушаларға колхицин қосу, ұршық түзілуін басады; сонымен қатар хроматид жұптары метафазада орналасқан орнында қалады. Бұл тәсіл хромосома санын анықтауға және олардың құрылымын микроскоп астында зерттеуге мүмкіндік береді. Модифицирленген центриоли сонымен қатар "базальдық денешіктер" деп аталатын кірпікшелер мен талшықтарда да болады.
23.3.2. Жасушаның бөлінуі
Цитоплазманың бөлінуі цитокинез деп аталады. Ол әдетте телофазамен қатар жүреді және интерфазаның G1 периодына апарады. Бөлінуге дайындық кезінде, жасуша органеллалары хромосомалармен бірге, телофаза жасушасының екі полюсіне бірдей бөлініп орналастырылады. Жануар жасушасында плазмалық мембрана телофаза кезінде ішке қарай, бұған дейін ұршық экваторы орналасқан деңгейде, сығылып кіре бастайды. Бұл, осы жерде орналасқан микрофиламенттердің әс-ірекетінен болады деп саналады. Сығылып кіру нәтижесінде, жасушаны экватор бойымен орайтын үздіксіз жүлге пайда болады. Ақырында, жүлге айналасындағы жасуша мембраналары, екі жасушаны толықтай бөлу арқылы, бірігеді.
Өсімдік жасушаларында ұршық жіпшелері телофаза кезінде жойыла бастайды; олар тек қана экваториалдық пластинка аймағында сақталады. Мұнда олар жасуша перифериясына қарай орын ауыстырады, олардың саны көбейеді және олар бөшке тәрізді денешік— фрагмопласт түзеді. Бұл аймаққа сонымен қатар микротүтікшелер, рибосомалар, митохондриялар, эндоплазмалық тор және Гольджи аппараты орын ауыстырады; соңғысы сұйықтықпен толтыралған көптеген ұсақ көпіршіктер түзеді. Көпіршіктер алдымен жасуша орталығында, содан соң микротүтікшелер арқылы бағытталып, экваторлық торда орналасатын, жасуша пластинкасын түзіп, орнын ауыстырады және бір-бірімен қосылады (5.30. сур. қара). Көпіршіктердің құрамы жаңа орта пластинканың және еншілес жасушаның іргесін құруға қатысады, ал олардың мембраналарынан жаңа сыртқы жасуша мембранасы пайда болады. Жасуша пластинкасы өсу арқылы, ақырында ата-ана жасушасы іргесімен қосылады және екі еншілес жасушаларды түгелдей бөледі. Жаңадан пайда болған жасуша іргелері біріншілік деп аталады; әрі қарай олар целлюлозаның қалдықты қабаты және лигнин және суберин тәрізді басқа да заттар есебінен, екіншілік жасуша іргесін құру арқылы, қосымша жуандай алады. Жасушаның белгілі аймақтарында жасуша пластинкасының көпіршіктері қосылмайды, сондықтан көршілес еншілес жасушалар арасында байланыс сақталады. Бұл цитоплазмалық каналдар, жасуша мембранасы арқылы жіберілген және олар плазмодесма деп аталатын құрылымдарды құрайды.
23.3.3. Жануар және өсімдік жасушаларындағы митозды салыстыру
Митоз кезіндегі болған ең маңызды әрекет бұл — екіеселенген хромосомаларды екі еншілес жасушалар арасына теңдей бөлу.
Митоз жануар және өсімдік жасушаларында, бірдей дерлік жүреді, бірақ айырмашылықтары да бар (23.1. кестесі).
23.1. кестесі. Жануар және өсімдік жасушаларындағы митоз арасындағы айырмашылықтар

23.3.4. Қысқаша қорытындылар
Митоздың нәтижесінде ата-аналық жасуша ядросы екі еншілес ядроға бөлінеді, олардың әрқайсысы ата-ана ядросында қанша болса, сонша хромосомадан тұрады. Бұлардың ізінен барлық жасушаның бөлінуі жүреді. Бұл мүмкін болуы үшін, хромосомалар алдымен интерфаза периодында көшірмеленеді. Көшірме (репликация) нәтижесінде пайда болған жұп құрылымдарды хроматидтер деп атайды және митоз кезінде олар әр-түрлі жасушаларға ажырайды.
23.3.5. Митоздың мәні
1. Генетикалық тұрақтылық. Митоздың нәтижесінде әрқайсысында ата-ана ядросындағыдай хромосома саны бар екі ядро пайда болады. Қанша дегенмен, хромосомалар ата-ана хромосомаларынан дәл ДНҚ репликациясы жолымен жүзеге асатын болғандықтан, олардың гендері бірдей генетикалық ақпаратты құрайды. Еншілес жасушалар ата-ана жасушаларымен бірдей болады, сондықтан митоз генетикалық ақпаратқа ешқандай өзгеріс енгізе алмайды. Сондықтан да дәл сол ата-ана жасушасынан пайда болған популяциясы, генетикалық тұрақты.
2. Бой. Митоздың нәтижесінде берілген ағзадағы жасуша саны өседі және бұл, көпжасушалы ағзалардың бойы негізінде жатыр(22 тарау).
3. Жасушалардың орын ауыстыруы. Жасушалардың және ұлпалардың орын ауыстыруы митозбен байланысты. Жасушалар әрдайым өледі және жаңадан ауысады- айқын үлгі ретінде тері жасушалары қызмет атқарады.
4. Регенерация. Кейбір жануарлар жоғалған дене бөліктерін регенерациялауға қабілетті, мысалы аяқ (шаянтәріздестер) немесе сәулелер (теңіз жұлдызы). Бұл үшін қажетті жасушалар митоз нәтижесінде пайда болады.
5. Жыныссыз көбею. Митоз жыныссыз көбеюдің негізінде жатыр— берілген түрдің ата-аналық жұбының біреуінен ғана өнген жаңа ағзалардың туындауы. Жыныссыз көбею көптеген түрлерге тән. Мұндай көбеюдің әр-түрлі тәсілдері 21 тарауда сипатталған.
23.4. Мейоз
Мейоз (грек тілінен аударғанда meiosis— азаю) —бұл диплоидтыдан(2n) гаплоидтыға(n) дейін хромосома санының азаюымен бірге жүретін ядроның бөліну формасы. Митоздағы сияқты, сонымен қатар интерфаза кезінде ДНҚ репликациясы ата-аналық жасушада жүреді, бірақ бұның артында мейоздың бірінші бөлінуі (мейоз І) және мейоздың екінші бөлінуі (мейоз ІІ) деп белгілі ядро және жасуша бөлінуінің екі циклі жүреді. Осылайша, бір диплоидтық жасуша төрт гаплоидтық жасушаға бастама береді, 23.10 суретінде сызба түрінде көрсетілгендей.
Мейоз жануарларда(21 тарау) спермий мен жұмыртқа жасушасы түзілуінен(гаметогенез) және өсімдіктерде споралардың түзілуінен жүреді.
Митоз тәрізді, мейоз да үздіксіз процесс, бірақ оны да ыңғайлылық үшін профаза, метафаза, анафаза және телофазаға бөлуге болады. Бұл кезеңдер мейоздың бірінші бөлінуінде жүреді және екінші бөлінуінде тағы бір рет қайталанады. Хромосомалардың осы кезеңдер барысындағы іс-әрекеті, төрт хромосомадан(2n=4), яғни екі жұп гомологиялық хромосомалардан құралатын ядроның бөлінуі көрсетілген 23.11. суретінде ұсынылған.
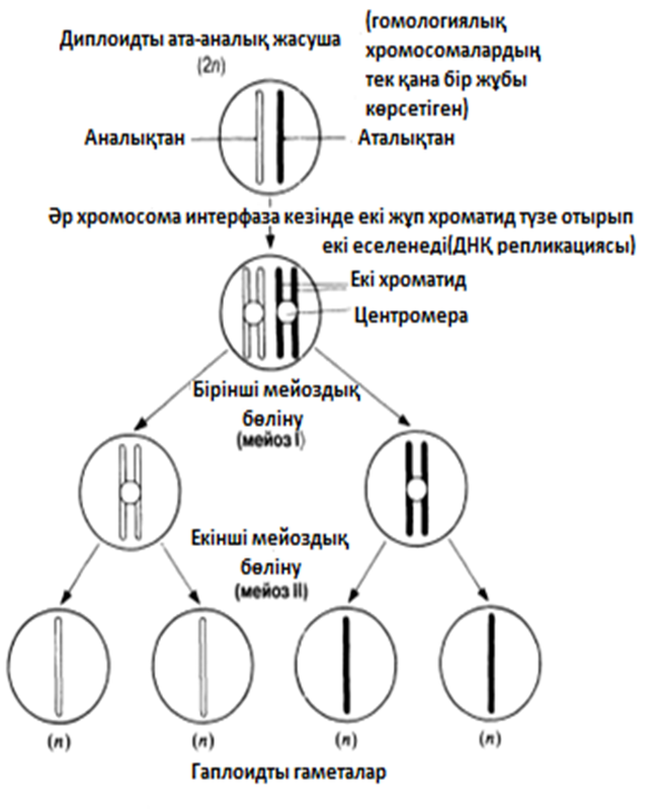
23.10. сур. Бір хромосоманың көбеюі және ядро мен жасушаның екі кейінгі бөлінуі үлгісіндегі, мейоздың басты кезеңдерінің сызба түріндегі суреті . Митоздағы тәрізді, хромосомалар жалғыз және қос құрылымды бола алатындығына назар аударыңыз. Екіеселенген хромосоманың екі бөлігін хроматидтер деп атайды.
23.4.1. Қысқаша қорытындылар
Мейоздың нәтижесінде бір ата-аналық жасушадан төрт еншілес жасуша пайда болады. Әр еншілес жасуша ата-ана жасушасындығы хромосома санының жартысынан тұрады. Әдетте ата-аналық жасуша диплоидтық болып келеді, сондықтан да еншілес жасушалар гаплоидтық болады.
23.4.2. Мейоздың мәні
1.Жыныстық көбею. Мейоз жыныстық жолмен көбейетін барлық ағзаларда жүреді. Ұрықтану кезінде екі гамета ядросы қосылады.
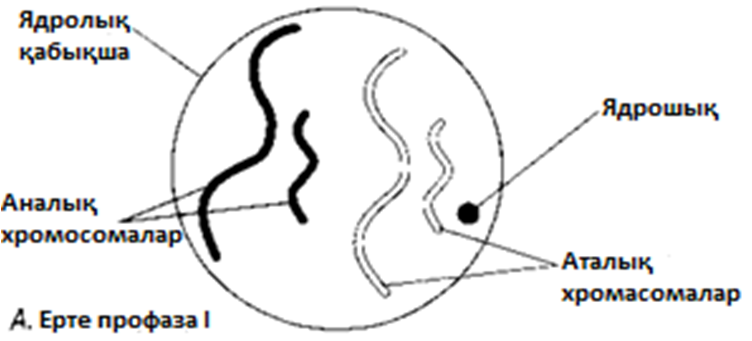
МЕЙОЗ I
Мейоз фотосуреттері 23.12 — 23.14.сур. ұсынылған.
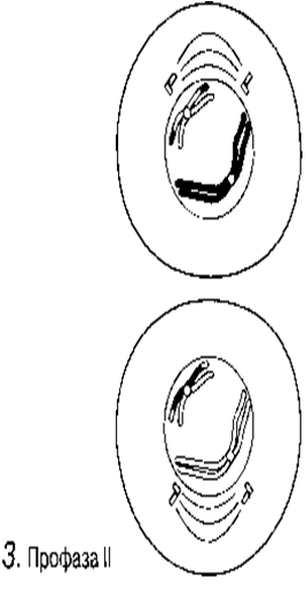
Профаза I
Ең созылмалы фаза.
А. Хромосомалар қысқарады және дискреттік құрылымдар тәрізді көрінетін болады.
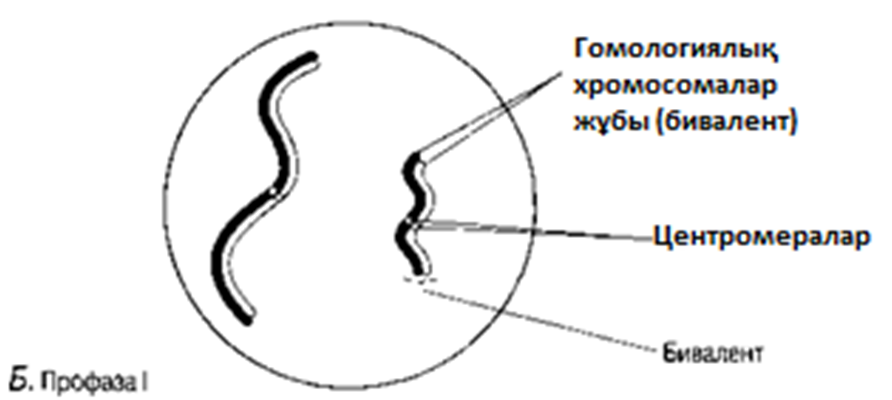
Б. Гомологиялық хромосомаларжұптасады. Бұл процесс синапсис деп, ал әр хромосома жұптары-бивалент деп аталады; бір хромосома аталық жынысты ата-анадан, ал екіншісі— аналық жынысты ата-анадан пайда болады. Биваленттің екі хромосомасы да бірдей ұзындықта болады, олардың центромералары бірдей орында болады және олар әдетте бәрдей ретте орналасқан бірдей ген сандарынан тұрады. Биваленттер ішінара бұралу есебінен қысқарады және жуандайды. Бұл кезеңдегі хромосомалар және центромералар анық ажыратылады.
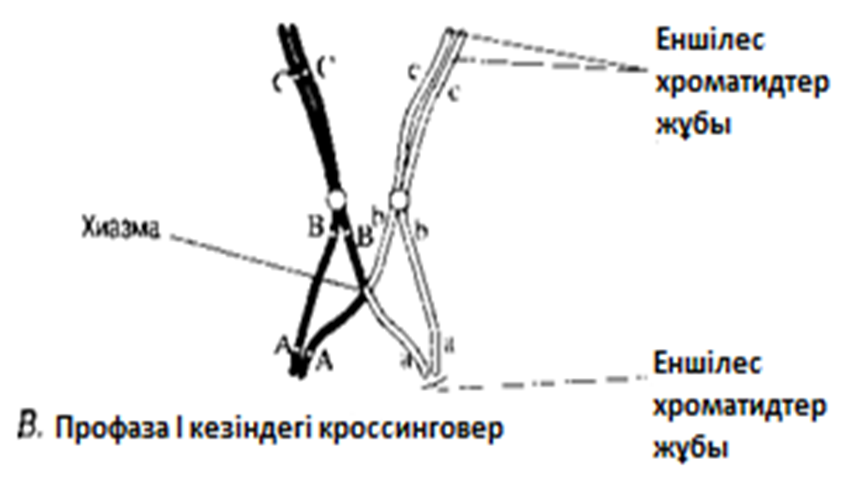
В. Гомологиялық хромосомалар бірін-бірі итергендей болып, ішінара бөлінеді. Олардың әрқайсысы екі хроматидтен тұратыны көрәне бастайды. Хромосомалар әлі де бір-бірімен бірнеше нүктелерде байланысқан. Бұл нүктелер хиазмалар деп аталады(грек тілінен chiasma —қиылыс). Әр хиазмада, хиазманың әрқайсысында болатын төрт жіптің әр екеуі арасындағы үзілу мен қайтадан қалпына келу нәтижесінде жүзеге асырылатын хроматидтер арасындағы алмаасу болады. Нәтижесінде, бір хромосома гендері (мысалы, аталық А, В, С) басқа хромосома гендерімен (аналық a, b, c) байланысты болып шығады, бұл қалыптасып жатқан хроматидтерде жаңа гендік комбинацияға әкеледі. Бұл процесс кроссинговер деп аталады.
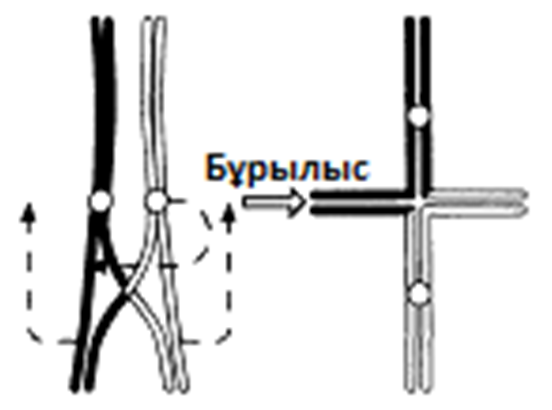
Г. Жалғаз хиазмасы бар бивалент: хроматидтердің бұрылысы нәтижесінде крест тәрізді құрылым түзілді
23.11. А — Г сур. Жануар жасушасындағы мейоз.
Г. Гомологиялық хромосомалардың хроматидтері бір-бірінен итерілуін жалғастырады және биваленттер хиазма сандарына байланысты, айқын конфигурация қабылдайды. (Бір хиазмалы биваленттер крест құрайды, ал екі хиазмалалар сақина тәрізді форма қабылдайды, ал үш немесе одан да көп хиазмалылар бір-біріне тік бұрышпен орналасқан ілмекті құрайды).Профаза І-дің соңында:
1) барлық хромосомалар түгелдей қысқарады және қарқынды боялады;
2) центриоли (егер бар болса) полюстерге қарай орын ауыстырады;
3)ядрошықтар және ядро қабықшасы ыдырайды;
4)ұршық жіпшелері пайда болады.
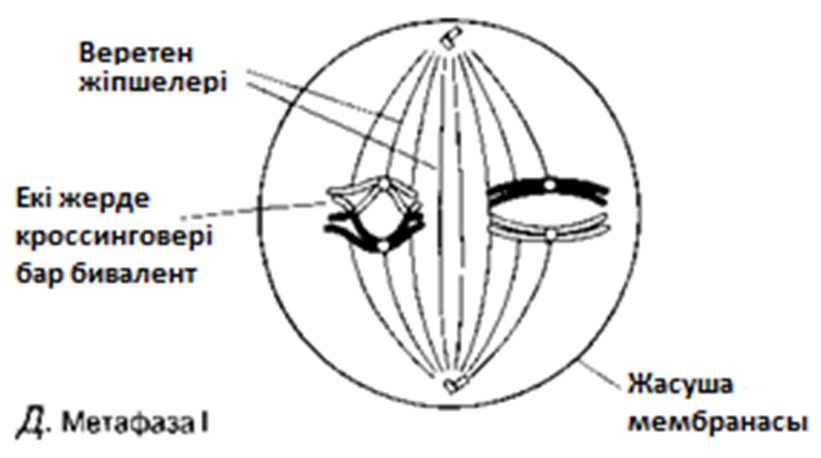
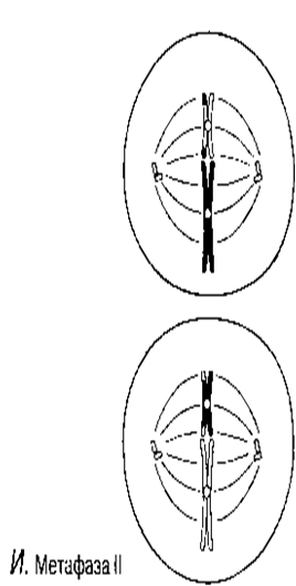
Метафаза I
Биваленттер центромераға жіпшелермен бекітілу арқылы, ұршық экваторына орналастырылады.
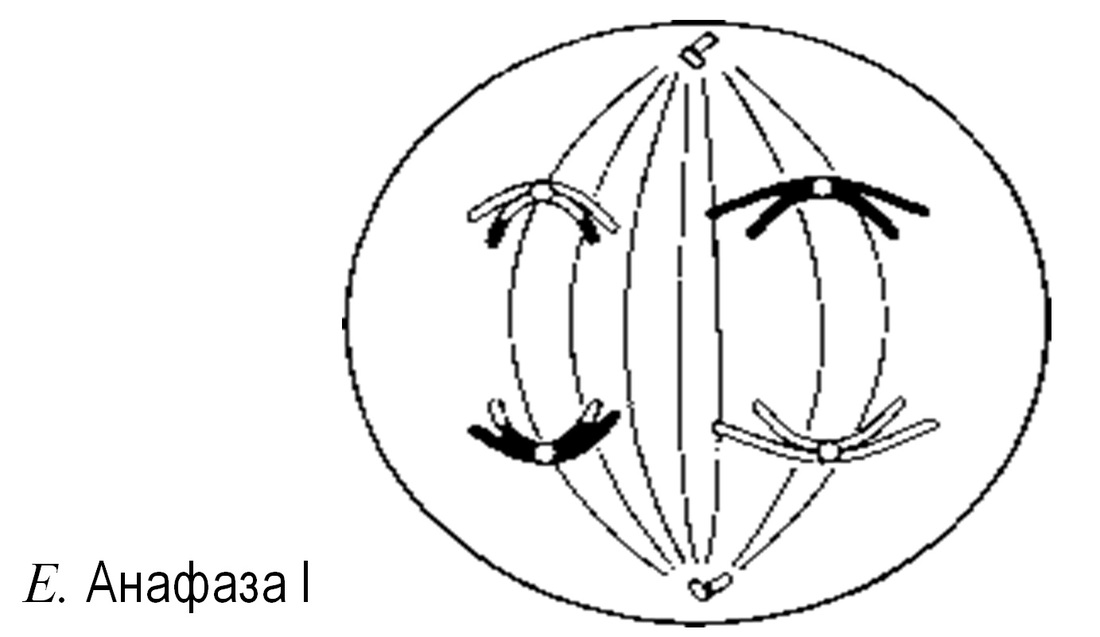
Анафаза I
Ұршық жіпшелері гомологиялық хромосомаларды центромерлерден бастап, қарама-қарсы полюстерге қарай тартады. Нәтижесінде әр ұршық полюсінде біреуден болатындай хромосомалар екі гаплоидтық жиынтыққа бөлінеді.
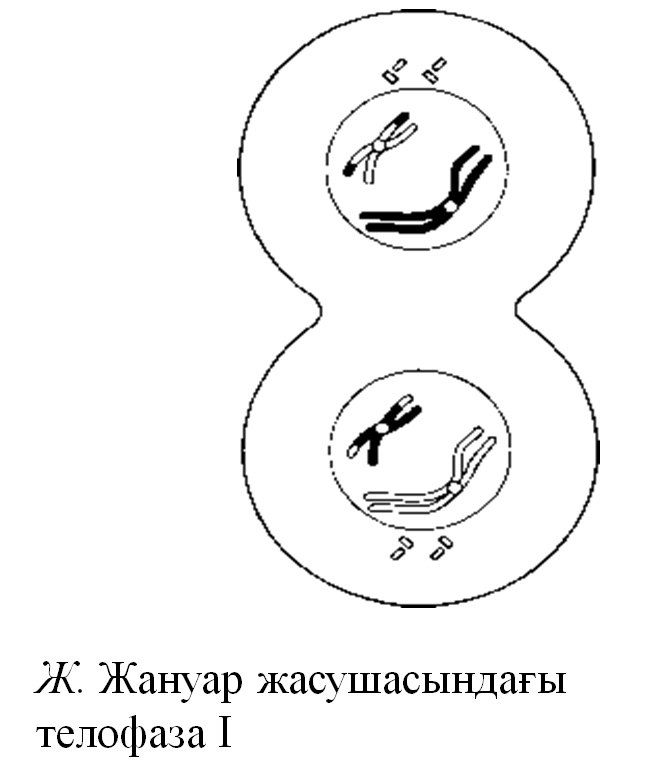
23.11. Д — Ж.сур. Жануар жасушасындағы мейоз
Телофаза I
Гомологиялық хромосомалардың қарама-қарсы полюстерге қарай алшақтауы мейоз І-дің аяқталуына сәйкес келеді. Хромосома саны екі есеге азайды, бірақ олардың әрқайсысы әлі де екі хроматидтен тұрады. Кроссинговер жүрген болса, онда бұл хроматидтер генетикалық бірдей емес және екінші мейоздық бөлінуде ажырауы тиіс. Ұршық және оның жіптері әдетте жойылады. Жануарларда және кейбір өсімдіктерде хроматидтер, әдетте, тарқатылады, әр полюсте қайтадан ядро қабықшасы пайда болады және ядро интерфазаға кіреді. Содан соң, митоздағыдай, ұсақталу (жануарларда) немесе жасуша іргесінің қалыптасуы (өсімдіктерде) болады. Көптеген өсімдіктерде не телофаза, не жасуша іргесінің қалыптасуы, не интерфаза бақыланбайды және анафаза І- ден шыққан жасуша бірден екінші мейоздық бөлінудегі профазаға өтеді.
Интерфаза ІІ
Бұл кезең, әдетте, тек қана жануар жасушасында болады. Оның ұзақтығы өзгеріп отырады. Одан әрі ДНҚ репликациясы болмайды.
|
|
МЕЙОЗ II
Мейоз II митозға ұқсас
Профаза II
Егер интерфаза ІІ болмаса, бұл кезең де болмайды. Ядрошықтар және ядро мембраналары бұзылады, ал хроматидтер қысқарады және жуандайды. Центриолалар, егер олар бар болса, жасушаның қарама-қарсы полюстеріне орын ауыстырады, және профаза ІІ-нің соңында жаңа ұршық жіпшелері пайда болады. Олар мейоз І-дің ұршығына тік бұрыш жасап орналасқан.
Метафаза II
Хромосомалар ұршық экваторы айналасына бөлек орналастырылады.
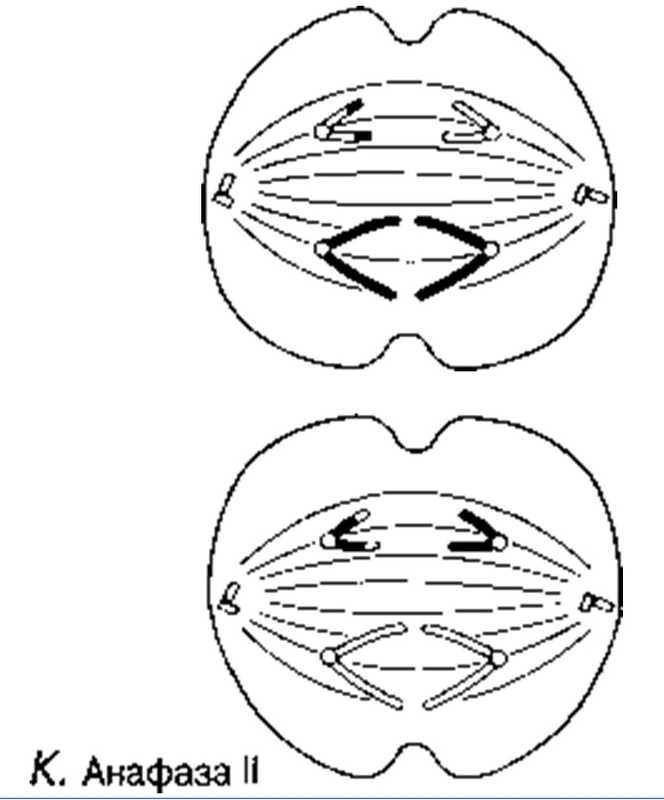
Анафаза II
Центромерлар бөлінеді және ұршық жіпшелері оларды тасып әкетеді, ал олардың артынан хроматидтер де қарама-қарсы полюстерге барады.
Л. Телофаза II және жануар жасушасындағы цитоплазманың бөлінуі
23.11. 3 — Л сур. Жануар жасушасындағы мейоз .
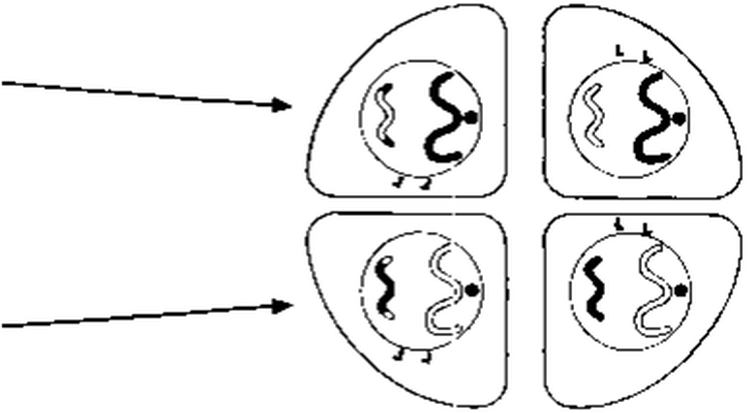
Телофаза II
Митоз телофазасы сияқты жүреді, бір ғана айырмашылық, мұнда төрт гаплоидтық еншілес жасуша түзіледі. Хромосомалар тарқатылады, ұзарады және нашар ажыратылатын болады. Ұршық жіпшелері жойылады, ал центриоли көшірмеленеді. Әр ядроның айналасында қайтадан ядро қабықшасы пайда болады, бірақ ядро енді бастапқы ата-ана жасушасының(ол гаплоидты) хромосома санының жартысын құрайды. Кейінгі ұсақтауда (жануарларда) немесе жасуша іргесі қалыптасуда (өсімдіктерде), бір ғана ата-аналық жасушадан төрт еншілес жасуша пайда болады.
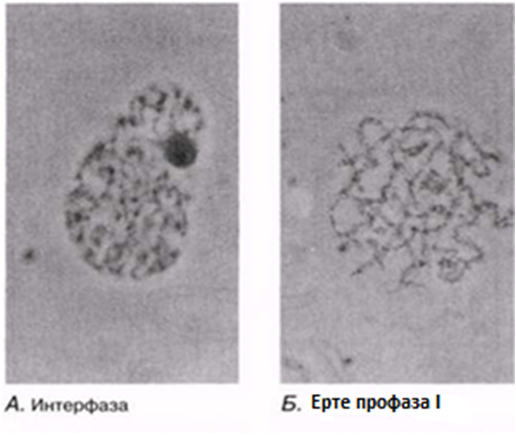
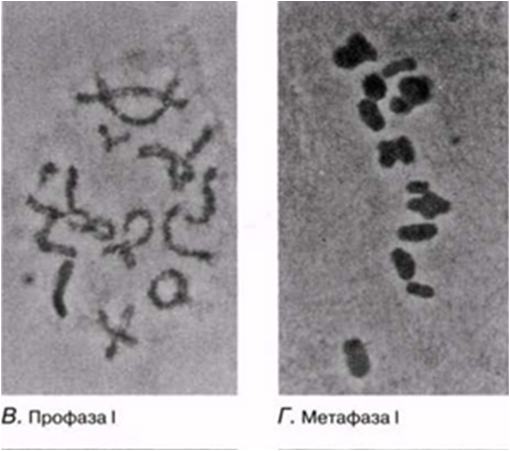
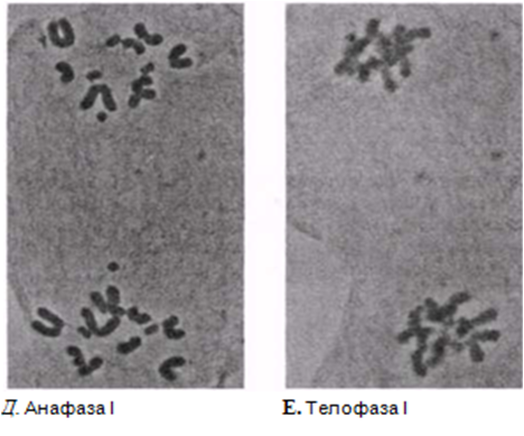
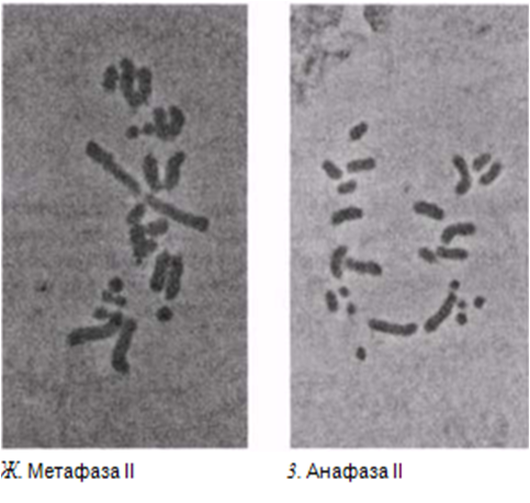
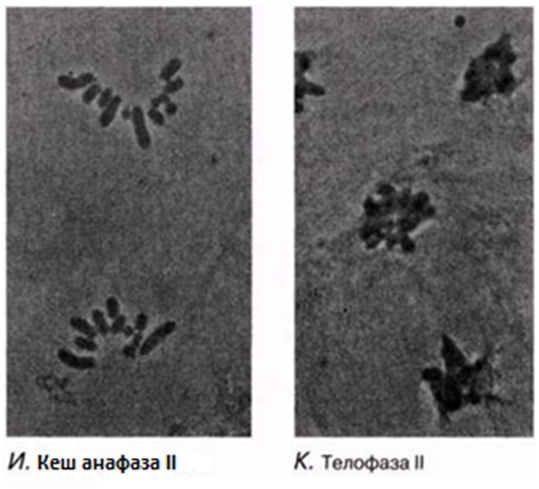
23.12. сур. Жануар жасушасындағы мейоз кезеңдері. Сонымен қатар, интерфаза да көрсетіген.
|
|
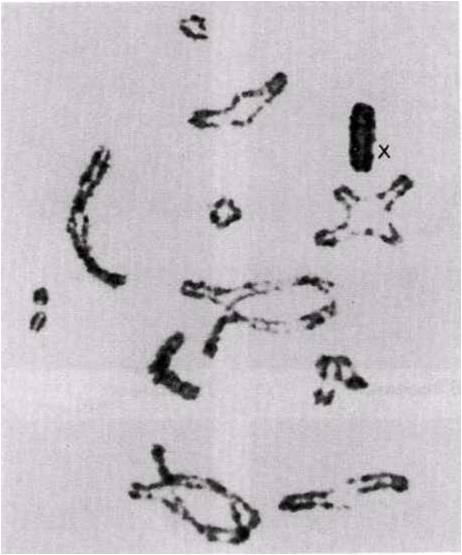
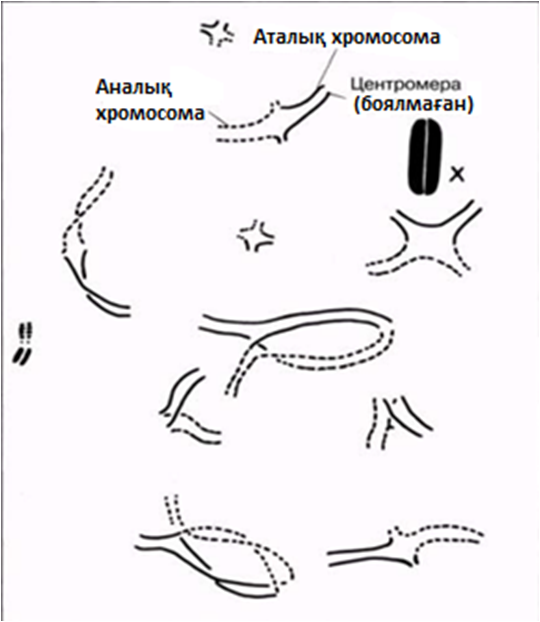
23.13.сур. Кезбе шегіртке (Locusta migratoria) жасушасындағы профаза І-дегі кроссинговер. Әрқайсысында бір немесе екі хиазмамен 11 бивалент көрінеді. Сызбада (оң жағында) аталық және аналық хромосомалар сәйкес толық және үзік сызықтармен салынған. Әр хиазмада генетикалық ақпаратпен алмасу жүрді. Биваленттердің пішіндері таяқша тәріздестен крест тәріздес немесе сақина тәріздеске дейін, хиазмалардың санына және орналасуына байланысты, өзгеріп тұрады. Бұл кезеңде Х-хромосома қарқынды боялады(Д-р S. A. Henderson).
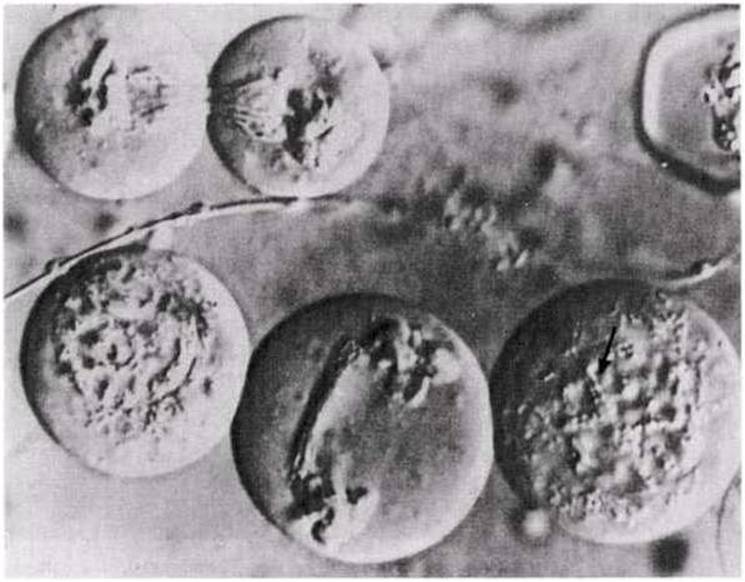
23.14.сур. Жануар жасушасындағы мейоз. Кезбе шегірткенің (Locusta migratoria) тірі сперматоциттеріндегі конъюгация және жасушалық бөліну. Препараттар Номарскийдің интерференциондық жырық тәсілімен түсірілген;бұл полярланған жарық қолданылатын тәсіл,тірі боялмаған жасушалардың көлемді картиналарын алуға мүмкіндік береді. Екі жасушада ерте профазаІ-дегі(бағытпен көрсетілген) хромосомалардың конъюгациясын көруге болады.Екі жасуша (жоғарыда сол жақта)мейоздың бірінші бөлінуін аяқтайды . Екі полюстік топ түзілгеннен кейін, барлық жасушаның бөлінуі басталады. Шамамен бірдей екі еншілес жасушалар түзіледі. Екі хромосома топтарының арасындағы жасушадан-жасушаға тартылып жатқан жіп тәріздес құрылымдар-бұл ұршық микротүтікшелері.
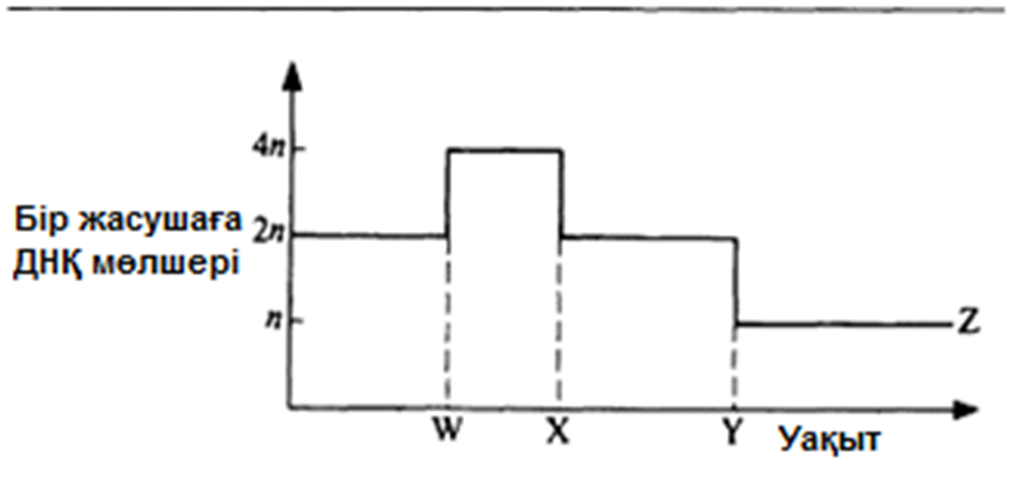
23.15. сур. 23.1. тапсырмасын орындау кезінде қолданылатын график.
Әр гамета бір хромосома жиынтығынан тұрады(яғни, ол гаплоидты, n).Гаметалар қосылуы нәтижесінде екі хромосома жиынтығы бар (яғни, диплоидты,2n) зигота түзіледі. Мейоз болмаған жағдайда, гаметалардың қосылуы, жыныстық көбею нәтижесінде пайда болатын, әр кейінгі ұрпақ хромосомаларының екі еселенуіне әкелетін еді. Осы тәртіптен тыс тек қана полиплоидияда бақылатантын еді (24.9.бөлім). Диплоидты хромосома санының (2n) гаплоидтыға (n) дейін қысқаратын ерекше жасушалық бөлінуінің арқасында бұл жынысты көбейетін ағзаларда жүзеге аспайды.

2. Генетикалық өзгергіштік. Мейоз сонымен қатар гаметалардың қосылуы нәтижесінде алынатын, ұрпақта генетикалық өзгергіштікке әкелетін, гаметаларда жаңа ген комбинацияларының пайда болуына мүмкіндік береді.
Метафаза І-дегі биваленттің мүмкін бөлінулері
|
Түзіле алатын ядро жасушалары |
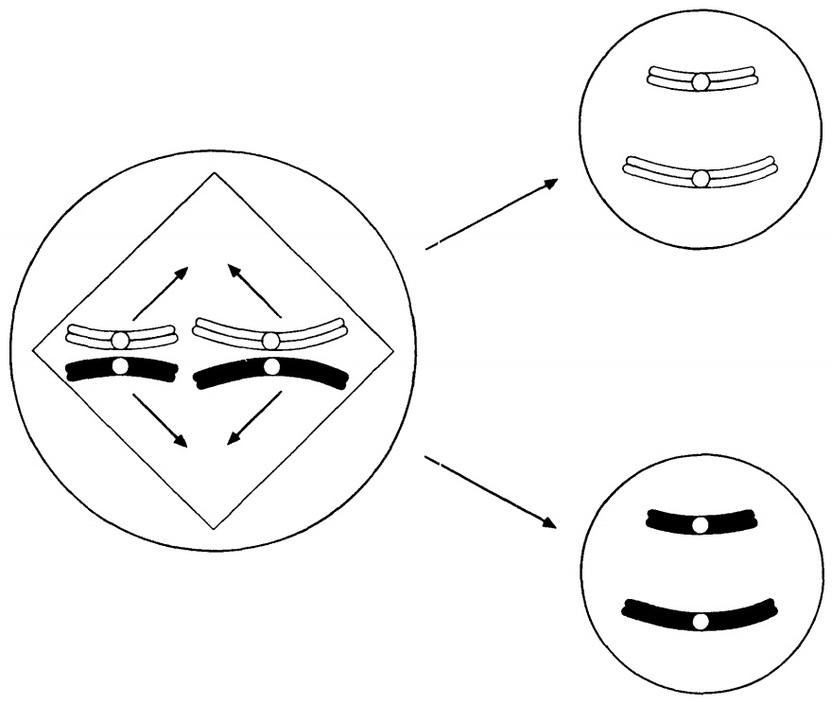
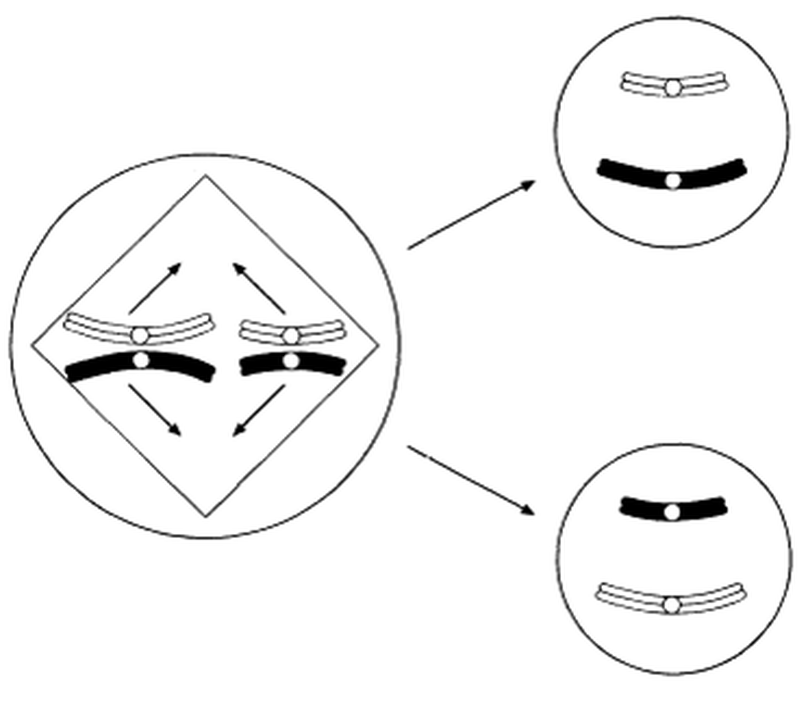
23.16. сур. Мейоздағы хромосомалардың тәуелсіз бөлінуі. Басқа биваленттерден тәуелсіз экватор айналасына орнығатын екі бивалент ұсынылған, сондықтан сызбада көрсетілген екі мысал да бірдей мүмкіндікке ие. Бұл, сонымен қатар суретте ұсынылған, гаметалардың ықтимал әртүрлілігін арттырады.
Мейоз процесінде бұл екі тәсілмен жүзеге асырылады, олар - хромосомалардың тәуелсіз бөлінуі және бірінші мейоздың бөлінуіндегі кроссинговер.
- Хромосомалардың тәуелсіз бөлінуі. Бұл процесті бәрінен гөрі сызба түрінде түсіндірген жақсырақ (23.16.сурет). Метафаза І-де биваленттер ұршық экваторында кездейсоқ түрде орналасады. 23.16.суретінде жай жағдай көрсетілген, онда тек қана екі бивалент қатысады, сондықтан да орналасу тек қана екі тәсілмен ғана мүмкін (олардың біреуінде ақ хромосомалар бір-біріне жақын орналасқан, ал басқасында-ақ хромосома қара хромосомаға жақын орналасқан). Бивалент саны көп болған сайын, мүмкін комбинациялар саны көп болады, ал сондықтан өзгергіштік жоғары болады. Тәуелсіз бөліну, анафаза І-дегі берілген бивалентті құрайтын хромосомалар, басқа биваленттер хромосомаларынан тәуелсіз бөлінетіндігін білдіреді.
- Кроссинговер. Профаза І- дегі гомологиялық хромосомалар арасында хиазма түзілуі нәтижесінде, гамета хромосомаларында жаңа ген комбинациясының түзілуіне әкелетін, кроссинговер жүреді. Мейоз нәтижесінде пайда болған барлық төрт гаметаның бір-бірінен ажыратылатындығы 23.11.Л. суретінде көрсетілген. Өзгергіштік 24.8.4. бөлімінде нақтырақ қаралады.
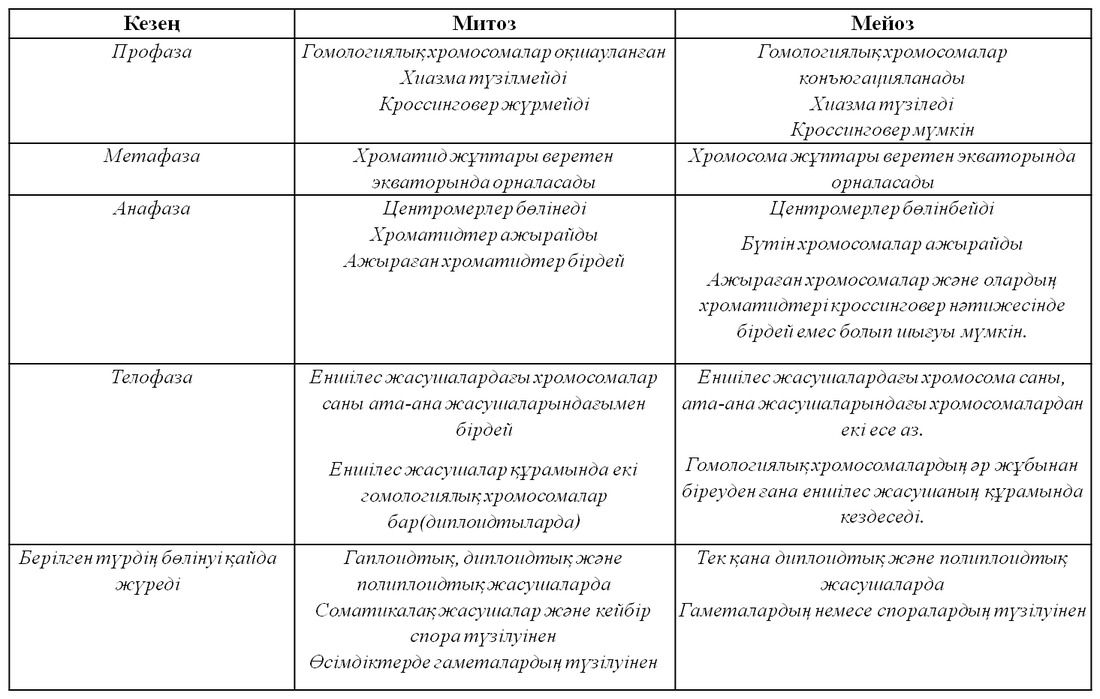
23.4.3. Митоз бен мейозды салыстыру
Митоз бен мейоз арасындағы биологиялық маңызды айырмашылықтар, митоз бен мейоз І арасындағы айырмашылықтарға келіп тіреледі. Мейоз ІІ мейозға бірдей дерлік ұқсас. Сондықтан да 23.2. кестесінде тек митоз бен мейоз І салыстырылады.
23.5. Хромосомалардың құрылымы
Эукариоттық жасуша хромосомасының анализі көрсетуі бойынша, олар дезоксиробонуклеин қышқылынан(ДНҚ) және нәруыздан, сонымен қатар РНҚ хромосомасының аз мөлшерінен тұрады. (прокариоттық жасуша «хромосомалары» — бактериялар— бір ДНҚ-дан тұрады). ДНҚ молекуласы, оның бүкіл ұзындығына таратылған теріс зарядтан тұрады, ал оған байланысқан нәруыздық молекулалар - гистондар оң зарядталған. Бұл ДНҚ—нәруыз кешені хроматин деп аталады.
Жасушада болатын ДНҚ-ның көп мөлшері, қаптама мәселесімен түйіседі. Мысалы, адамның бір жасушасы, 46 хромосомалардың өзара үлестірілген, шамамен 2,2 м ДНҚ-дан құралады. Осылайша, әр хромосома құрамында, шамамен 4,8м(48000 мкм) ДНҚ болады. Адамның хромосома ұзындығы орташа 6 мкм-ді құрайды, қаптама коэффициенті 8000:1. Құрастырылған ДНҚ-да ұйымдасудың жоғары деңгейін қамтамасыз ету үшін, гистондық нәруыздар ДНҚ үшін өте дәл жоспарланған шикізат көзін құрайды.
ДНҚ спиралі 8 гистондық молекула топтарымен нуклеосома —жіпке тізілген моншақ тәрізді бірлік түзе отырып байланысатыны көрсетілген болатын. Нуклеосомалар және оларды байланыстыратын ДНҚ аймақтары тығыз қапталған; олар әр орамға шамамен 6 нуклеосомадан келетін, қалыңдығы «30 нм талшықтар» немесе соленоидты талшықтар аттарымен белгілі. Оның қаптама коэффициенті 40-қа тең, яғни ұзындығы 1 мкм соленоид аймағында 40 мкм ДНҚ қапталған. Соленоидтық талшықтардың және «қапталмаған» соленоидтың («моншақ жіпшесі») сыртқы түрі 23.17. суретінде көрсетілген.
Қанша дегенмен, ДНҚ тығызырақ қапталуы тиіс болғандықтан, соленоидтардың өзі қалай болса да бүктелген және оралған болуы тиіс. Бұған қалай қол жеткізуге болатындығы әлі белгісіз болып қалады. Қазіргі уақытта бізде бар жалғыз ақпарат, бұл ерекше хромосомалары бар бірнеше жасушалар туралы мәліметтер. Құрамында «шамдық щетка» тәрізді хромосомалары бар деп, өзінің бұрынғы кездері керосин шамының шынысын тазалаған щеткаға ұқсас болғандықтан аталып кеткен, сондай бір жасуша түрі—амфибия ооциті(жұмыртқажасуша). Бұл хромосомалардың метафаза кезіндегі электрондық микрофотосуреттерінде әр хроматидтің екі ДНҚ спиралінің біреуінен тұратын, бірнеше ілмектер шығатын, тығыз оралған осьтік жіпшелерден тұратындығы көрінеді(23.18. сур.). Бұл ілмектер, мүмкін, өздерімен бірге белсенді ДНҚ-ны, яғни транскрипцияны жүзеге асыру үшін нәруыздан босаған ДНҚ-ны көрсетеді (23.8.6. бөлім).
Болжамды хромосома құрылымы 23.17 суретінде көрсетілген.
23.2.Кестесі . Митоз және мейоз І-ді салыстыру
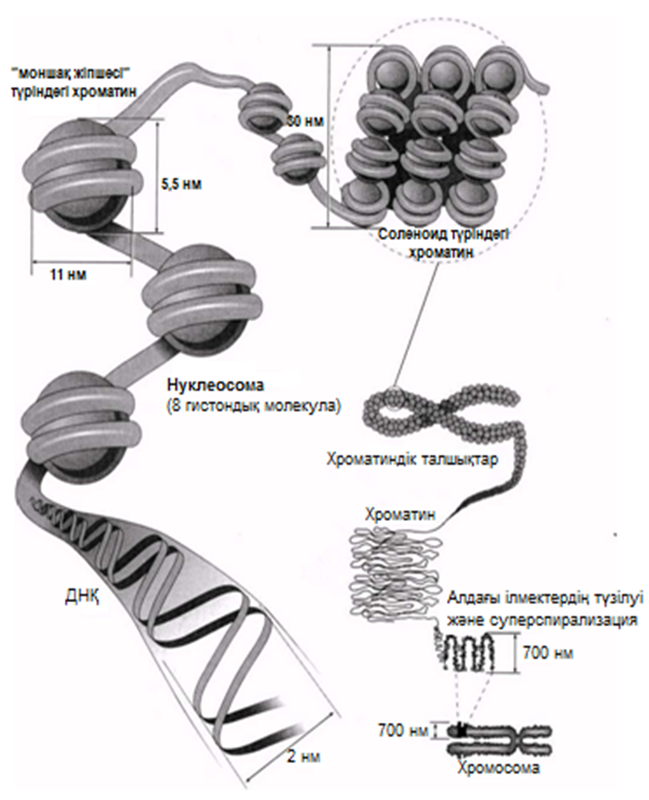
23.17. сур. Нуклеосоманың және оның хромосомамен және ДНҚ молекуласымен байланысының ұсынылған құрылымы.
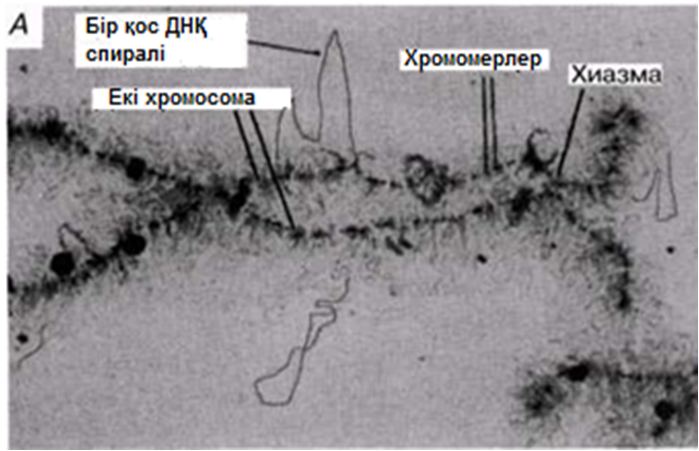
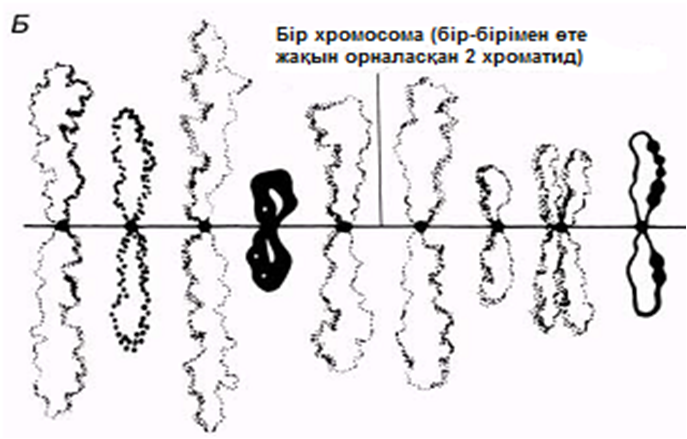
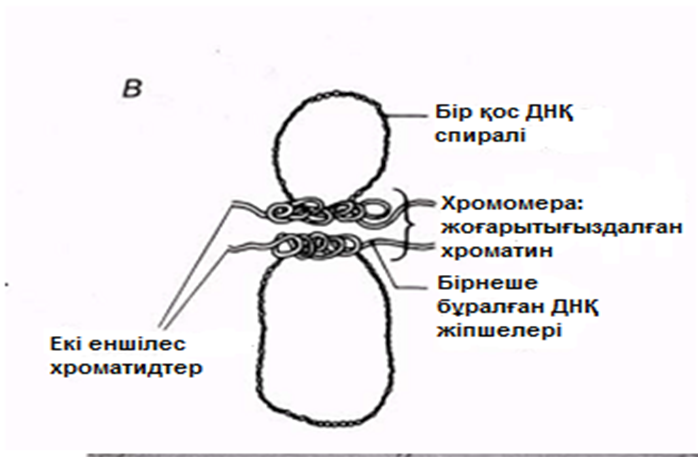
23.18.А. сур. Амфибия ооцитінен шамдық щетка тәрізді хромосома жұптары. Б және В. Хромосомалар,мРНҚ синтезі жүретін орталық ДНҚ жіпшесін және ДНҚ ілмегін көрсету үшін созылған.Тығыздалған аймақтар хромомерлерді құрайды. Әр хромомера және онымен байланысқан ілмек анықталған гендік локусқа сәйкес келетіндігі болжанады.( H.G.Callan, Int. Rev. Cytology, 1963, 15, 1).
23.6. ДНҚ
23.6.1. Тұқымқуалаушылықтағы ДНҚ-ның рөлін нұсқайтын ақпараттар
ХХ ғасырдың басында Саттон мен Бовери, хромосомалар ғана гендік ақпаратты ұрпақтан-ұрпаққа жеткізетіндігі туралы ой айтты. Бірақ, генетикалық материал ретінде ДНҚ немесе хромосома нәруызы қызмет атқаратыны анықталғанша, әлі де көп жылдар өтті. Ғалымдар, генетикалық материал ретінде қызмет атқару үшін, молекуласы жеткілікті құрылымдық алуан-түрлілікті қамтитын, бір ғана зат—нәруыз деп санауға бейімделген болатын.
Бактериялар туралы ақпараттар
1928 жылы ағылшын микробиологі Фредерик Гриффит болашақта бұл мәселенің шешілуінде маңызды рөл атқаратын, бақылау жасады. Антибиотиктер әлі болмаған кездері, пневмония өлім шығынына жиі әкелді. Гриффит пневмонияның бір түрінің қоздырғышы—пневмококкқа қарсы вакцина алуға әрекеттенді. Бұл бактерияның екі түрі белгілі болды, олардың бірі қоймалжың капсуламен қапталған және вирулентті(ауру тудырады), ал екіншісінің капсуласы болмайды және вирулентті емес. Көрінгені бойынша,капсула бактерияны белгілі бір тәсілмен адамның иммундық жүйесінен қорғады.
Гриффит, егер науқас ағзасына капсуласыз немесе қыздыру арқылы өлген қапталынған формасын енгізсе, оның ағзасы пневмония ауруынан сақтандыра алатын антиденелер түзеді деп үміттенді. Эксперименттер қатарында Гриффит тышқандарға бактерияның екі формасын да енгізіп, 23.3. кестесінде ұсынылғандай нәтижелер алған.
23.6. ДНҚ
23.6.1. Тұқымқуалаушылықтағы ДНҚ-ның рөлін нұсқайтын ақпараттар
ХХ ғасырдың басында Саттон мен Бовери, хромосомалар ғана гендік ақпаратты ұрпақтан-ұрпаққа жеткізетіндігі туралы ой айтты. Бірақ, генетикалық материал ретінде ДНҚ немесе хромосома нәруызы қызмет атқаратыны анықталғанша, әлі де көп жылдар өтті. Ғалымдар, генетикалық материал ретінде қызмет атқару үшін, молекуласы жеткілікті құрылымдық алуан-түрлілікті қамтитын, бір ғана зат—нәруыз деп санауға бейімделген болатын.
Бактериялар туралы ақпараттар
1928 жылы ағылшын микробиологі Фредерик Гриффит болашақта бұл мәселенің шешілуінде маңызды рөл атқаратын, бақылау жасады. Антибиотиктер әлі болмаған кездері, пневмония өлім шығынына жиі әкелді. Гриффит пневмонияның бір түрінің қоздырғышы—пневмококкқа қарсы вакцина алуға әрекеттенді. Бұл бактерияның екі түрі белгілі болды, олардың бірі қоймалжың капсуламен қапталған және вирулентті(ауру тудырады), ал екіншісінің капсуласы болмайды және вирулентті емес. Көрінгені бойынша,капсула бактерияны белгілі бір тәсілмен адамның иммундық жүйесінен қорғады.
Гриффит, егер науқас ағзасына капсуласыз немесе қыздыру арқылы өлген қапталынған формасын енгізсе, оның ағзасы пневмония ауруынан сақтандыра алатын антиденелер түзеді деп үміттенді. Эксперименттер қатарында Гриффит тышқандарға бактерияның екі формасын да енгізіп, 23.3. кестесінде ұсынылғандай нәтижелер алған.
23.3.кестесі. Гриффиттің эксперименттер нәтижелері

Өлген тышқандарды ашу кезінде, оларда тірі қапталынған формалар табылды. Бұл нәтижелер негізінде, Гриффит, қыздырудан өлген қапталынған формалардан, тірі капсуласыз формаларға, оларды капсулалар шығаруға және вирулентті болуға мәжбүрлейтін, қандай да бір фактордың анық берілетіндігі туралы қорытынды жасады. Бірақ бұл тасымалдаушы фактордың туындауы 1944 жылға, яғни оны айыру және анықтау іске асқанға дейін белгісіз болып қалды.
Ондаған жылдар бойы Эвери, Мак-Карти және Мак-Леод, қыздыру әсерінен өлген қапталынған пневмококк жасушасының құрамына кіретін молекулалардың бөлінуімен және тазартылуымен айналысты және олардың капсуласыз жасушаларды тасымалдау қабілетін зерттеді. Полисахаридтік капсуланың және жасушалық сығындыдағы нәруыздық фракцияның жойылуы, тасымалдауға ықпал еткен жоқ, бірақ ДНҚ-ны гидролиздейтін дизоксирибонуклеоаза ферментінің қосылуы оған кедергі келтірді. ДНҚ-ның тазалығы жоғары сығындысының, қапталынған жасушадан тасымалдау туғызу қабілеті, Гриффиттің тасымалдаушы факторы ДНҚ болғандығын көрсетті. Бұл нәтижелерге қарамастан, кейбір ғалымдар генетикалық материал ретінде нәруыз емес, ДНҚ қызмет атқаратынын мойымдаудан бас тартты. Елуінші жылдардың басында вирустарды зерттеу кезінде алынған көптеген қосымша ақпараттар, соңында, генетикалық ақпараттың тасымалдаушысы ретінде ДНҚ қызмет атқаратындығын көрсетті.
Вирустар туралы ақпараттар
ХХ ғасырдың 40-ыншы жылдары вирустар эксперименттік генетикалық зерттеулердің басты нысандарының бірі болды. Вирустық бөлшектер өте қарапайым құрылымды болады; олар нәруызды қабықшадан және ондағы нуклеин қышқылы молекулалары—ДНҚ немесе РНҚ (2.4.2. бөлім). Бұл оларды, генетикалық материал ретінде—нәруыз немесе нуклеин қышқылы қызмет атқаратындығы туралы сұрақты зерттеу үшін тамаша материал етеді. 1952 жылы Херши және Чейз бактериялық жасушаларды жұқтыратын және бактериофаг деп аталатын вирустың ерекше түріне арналған эксперименттер қатарына кірісті. Т2 бактериофагі адам ішегінде өмір сүретін ішек таяқшасының жасушасына енеді, және оны өте аз уақыт ішінде көптеген Т2 фагі бөлшектерін түзуге мәжбүрлейді. Херши және Чейз Т2 фагі бөлшектерін, радиоактивті изотоптар күкірт(35S) немесе фосформен(32P) бір ортада өсетін E.coli жасушасында дамытты.
Фаг нәруыздары құрамында күкірт бар, бірақ фосфор жоқ, ал ДНҚ құрамында фосфор бар, бірақ күкірт жоқ. Сондықтан да радиоактивті күкіртпен таңбаланған, E.coli-де түзілген фаг бөлшектері оны өзінің нәруызды қабықшасына қосып алды, радиоактивті фосформен таңбаланған, E.coli-де сонымен қатар түзілген бөлшектер құрамында 32P-мен таңбаланған ДНҚ болды.
Т2 таңбаланған фаг бөлшектері, таңбаланбаған E.coli жасушаларына жұқтырды және бірнеше минут өткен соң, фагтік бөлшектерді бактериялық жасушалар іргесінен бөліп алу үшін, бұл жасушаларды араластырғышта сілкіп алды. Содан соң бактерияларды инкубациялады және радиоактивтілігін анықтау үшін сынады. Нәтижелер 23.19. суретінде ұсынылған.
Алынған нәтижелер негізінде Херши мен Чейз, бактериялық жасушаға нәруыз емес, көптеген фагтік ұрпақтарға бастама беретін, фагтік ДНҚ енетіні жөнінде қорытынды жасады. Бұл эксперименттер тұқымқуалаушылық материал ретінде ДНҚ қызмет атқаратындығын көрсетті. Электронды-микроскоптық зерттеулер нәтижелері және вирустардың өмірлік циклі туралы толығырақ ақпараттар, бактериялық жасушаға тек қана фагтік ДНҚ енетіндігін дәлелдейді. Вирустық және фагтік бөлшектердің өмірлік циклі 2.4.3. және 2.4.5.бөлімдерінде сипатталған.
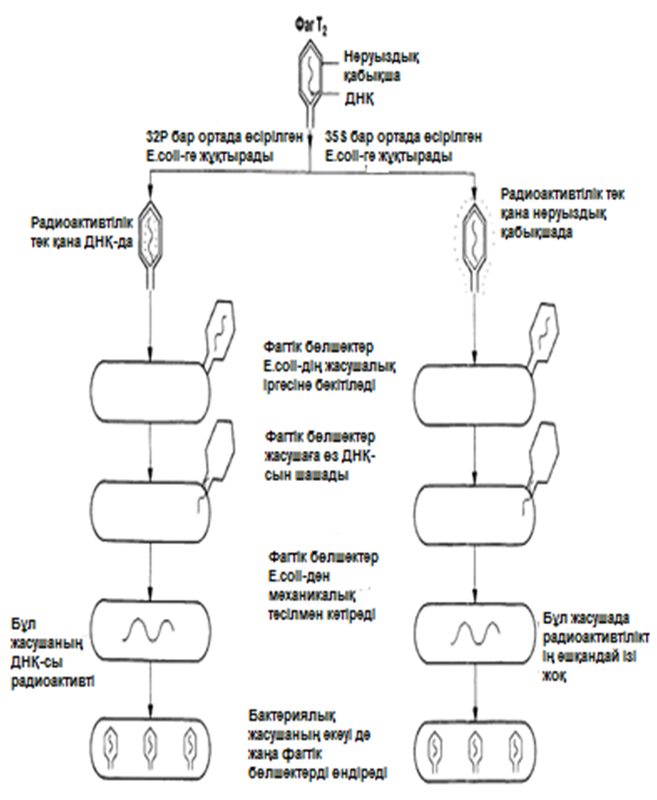
23.19.сур. Альфред Херши және Марта Чейздің Т2 фагіне және E.colі-ге жасаған эксперименттерінің сызбасы
23.6.2. ДНҚ репликациясы
Уотсонмен және Крикпен ұсынылған екі спираль түріндегі ДНҚ құрылымының моделі, 3.6.3.бөлімінде сипатталған. Бұл модельдің тартымды ерекшеліктерінің бірі, оның бір мезгілде, ДНҚ репликациясының қандай тәсілмен болатындығын көрсете алатындығында. Уотсон және Крик, спираль құрайтын екі тізбектің, тарқатыла және бөліне алатындығы жөнінде болжам білдірді; сонымен қатар олар, негіздердің жұптасу жолы арқылы, нуклеотидтердің комплементарлық тізбекшесі қосылатын, матрица ретінде қызмет атқарады. Осылайша ДНҚ-ның әр бастапқы молекуласынан, бірдей құрылымды екі көшірме пайда болады.
1956 жылы Корнбергке матрица ретінде жалғыз ДНҚ тізбегін қолдана отырып, ДНҚ молекуласының синтезін пробиркада көрсетуге мүмкіндік туды. Корнберг E.coli-ден ДНҚ комплементарлы тізбегі түзілу арқылы, энергия көзі түрінде АТФ болған кезде, бір-бірімен бос нуклеотидтер байланыстыра алатын ферменттерді бөліп алды және тазалады. Ол бұл ферментті ДНҚ-полимераза деп атады. Алдағы эксперименттердің көрсетуі бойынша, жасушада қолданылатын нуклеотидтер құрамында екі қосымша фосфаттық топтар бар. Ол нуклеотидтерді белсендендіреді. Әр нуклеотидтің, ДНҚ-ның өсіп жатқан тізбегіне бекітілуіне қарай, екі қосымша фосфаттық топтар жұлынады. Бұдан босап шыққан энергия, қалған нуклеотидтің фосфаттық тобымен бірге, көршілес нуклеотид молекуласындағы секер қалдығымен байланыс құру үшін пайдаланылады. Репликация процесі 23.20. суретінде көрсетілген. Ол геликаза ферментімен басқарылатын, ДНҚ-ның екі спиралінің тарқатылуынан басталады. Содан соң ДНҚ-полимераза біртізбекшелі ДНҚ-ға бекітіледі және тізбек бойымен жылжи бастайды. Ол кезекті ДНҚ тізбегіндегі негізге дейін жеткен сайын, бос нуклеотидтер тізбекке жақындайды және олардың құрамында комплементарлық негізі бар түрі, олармен сутектік байланыс түзеді. Бос нуклеотид алдыңғы нуклеотидке қосылғанға дейін осы тәсілмен жаңа ДНҚ тізбегін өсіру арқылы,орнында ферментпен ұсталып алынады. Бұл өсу тек қана 5’→3’ бағытында жүруі мүмкін. 23.20. суретінде көруге болатындай, ДНҚ полимераза, тарқатылатын фермент бағытымен бір бағытта қозғалатын болғандықтан, бұл бір ғана ДНҚ тізбегінің үздіксіз көшірмеленуінің мүмкіндігін білдіреді. Бұл процессті үздіксіз репликация деп атайды. Басқа тізбектің көшірмеленуі әр кезде қайтадан басталуы керек, себебі ДНҚ- полимераза тарқатылатын ферменттен 5’→3’ бағытында алшақтау керек. Нәтижесінде, тізбекте кішкене арақашықтықтықтар пайда болады, себебі ДНҚ-полимераза жаңадан синтезделген бір ДНҚ аймағының 3’-ұшын, келесісінің 5’-ұшымен байланыстыра алмайды. Бұл арақашықтықтарды жою үшін, ДНҚ-лигаза деп аталатын басқа фермент қажет. Мұндай репликацияны үзілісті деп атайды.
Жартылай консервативті репликация механизмінің пайдасына ақпараттар
ДНҚ репликациясының Уотсонмен және Крикпен ұсынылған және 23.20. суретінде көрсетілген тәсілі, осы кезде әр жаңа екі спираль, бастапқы екі ДНҚ спиралі тізбегінің екеуінің біреуін сақтайтын болғандықтан, жартылай консервативті репликация атымен белгілі. Бұл механизм 1958 жылы Мезелсонмен және Стальмен классикалық эксперименттер сериясында алынған ақпараттарға негізделген. E.coli жасушасы құрамында бір сақина тәріздес хромосома бар; бұл жасушаларды көптеген ұрпақтар ағымында, құрамында ауыр азот(15N) изотопы бар ортада өсіру кезінде, олардың барлық ДНҚ-счы осы изотоппен таңбаланған болып шықты. Таңбаланған ДНҚ-сы бар жасушаларды, құрамында қарапайым азот изотопы бар 14N бар ортаға ауыстырды. E.coli генерациясы уақытына сәйкес келетін (50мин 36ᵒС)—бір жасушалық бөлінуге және бір ДНҚ репликациясына қажетті уақыттың жылдамдығы өткеннен кейін,—жасуша үлгілерін таңдайды, олардан ДНҚ-ны бөледі және 20 сағ ішінде 40000g-та, цезий хлориді( CsCl) ерітіндісінде центрифугалайды(орталықтан тебу). Сонымен қатар ауыр CsCl молекулалары, пробирканың жоғарғы жағынан түбіне дейін өсетін тығыздық градиентін қалыптастыра отырып, тұна бастайды. ДНҚ тығыздығы CsCl ерітіндісінің тығыздығына тең деңгейге жиналады. Ультрафиолеттегі зерттеулер кезінде, орталықтан тепкіш пробиркадағы ДНҚ, тар жолақ түрінде болады. 15N және 14N орталарында өсірілген, жасушадан бөлінген ДНҚ жолақтарының орналасуы және алынған ақпараттардың талдануы 23.21.суретінде ұсынылған. Жартылай консервативті репликация сызбасы 23.22. суретіне қоса берілген. Бұл эксперименттер ДНҚ репликациясы жартылай консервативті тәсілмен болатындығын көрсетті.
Уотсонмен және Крикпен ұсынылған екі спираль түріндегі ДНҚ құрылымының моделі, 3.6.3.бөлімінде сипатталған. Бұл модельдің тартымды ерекшеліктерінің бірі, оның бір мезгілде, ДНҚ репликациясының қандай тәсілмен болатындығын көрсете алатындығында. Уотсон және Крик, спираль құрайтын екі тізбектің, тарқатыла және бөліне алатындығы жөнінде болжам білдірді; сонымен қатар олар, негіздердің жұптасу жолы арқылы, нуклеотидтердің комплементарлық тізбекшесі қосылатын, матрица ретінде қызмет атқарады. Осылайша ДНҚ-ның әр бастапқы молекуласынан, бірдей құрылымды екі көшірме пайда болады.
1956 жылы Корнбергке матрица ретінде жалғыз ДНҚ тізбегін қолдана отырып, ДНҚ молекуласының синтезін пробиркада көрсетуге мүмкіндік туды. Корнберг E.coli-ден ДНҚ комплементарлы тізбегі түзілу арқылы, энергия көзі түрінде АТФ болған кезде, бір-бірімен бос нуклеотидтер байланыстыра алатын ферменттерді бөліп алды және тазалады. Ол бұл ферментті ДНҚ-полимераза деп атады. Алдағы эксперименттердің көрсетуі бойынша, жасушада қолданылатын нуклеотидтер құрамында екі қосымша фосфаттық топтар бар. Ол нуклеотидтерді белсендендіреді. Әр нуклеотидтің, ДНҚ-ның өсіп жатқан тізбегіне бекітілуіне қарай, екі қосымша фосфаттық топтар жұлынады. Бұдан босап шыққан энергия, қалған нуклеотидтің фосфаттық тобымен бірге, көршілес нуклеотид молекуласындағы секер қалдығымен байланыс құру үшін пайдаланылады. Репликация процесі 23.20. суретінде көрсетілген. Ол геликаза ферментімен басқарылатын, ДНҚ-ның екі спиралінің тарқатылуынан басталады. Содан соң ДНҚ-полимераза біртізбекшелі ДНҚ-ға бекітіледі және тізбек бойымен жылжи бастайды. Ол кезекті ДНҚ тізбегіндегі негізге дейін жеткен сайын, бос нуклеотидтер тізбекке жақындайды және олардың құрамында комплементарлық негізі бар түрі, олармен сутектік байланыс түзеді. Бос нуклеотид алдыңғы нуклеотидке қосылғанға дейін осы тәсілмен жаңа ДНҚ тізбегін өсіру арқылы,орнында ферментпен ұсталып алынады. Бұл өсу тек қана 5’→3’ бағытында жүруі мүмкін. 23.20. суретінде көруге болатындай, ДНҚ полимераза, тарқатылатын фермент бағытымен бір бағытта қозғалатын болғандықтан, бұл бір ғана ДНҚ тізбегінің үздіксіз көшірмеленуінің мүмкіндігін білдіреді. Бұл процессті үздіксіз репликация деп атайды. Басқа тізбектің көшірмеленуі әр кезде қайтадан басталуы керек, себебі ДНҚ- полимераза тарқатылатын ферменттен 5’→3’ бағытында алшақтау керек. Нәтижесінде, тізбекте кішкене арақашықтықтықтар пайда болады, себебі ДНҚ-полимераза жаңадан синтезделген бір ДНҚ аймағының 3’-ұшын, келесісінің 5’-ұшымен байланыстыра алмайды. Бұл арақашықтықтарды жою үшін, ДНҚ-лигаза деп аталатын басқа фермент қажет. Мұндай репликацияны үзілісті деп атайды.
Жартылай консервативті репликация механизмінің пайдасына ақпараттар
ДНҚ репликациясының Уотсонмен және Крикпен ұсынылған және 23.20. суретінде көрсетілген тәсілі, осы кезде әр жаңа екі спираль, бастапқы екі ДНҚ спиралі тізбегінің екеуінің біреуін сақтайтын болғандықтан, жартылай консервативті репликация атымен белгілі. Бұл механизм 1958 жылы Мезелсонмен және Стальмен классикалық эксперименттер сериясында алынған ақпараттарға негізделген. E.coli жасушасы құрамында бір сақина тәріздес хромосома бар; бұл жасушаларды көптеген ұрпақтар ағымында, құрамында ауыр азот(15N) изотопы бар ортада өсіру кезінде, олардың барлық ДНҚ-счы осы изотоппен таңбаланған болып шықты. Таңбаланған ДНҚ-сы бар жасушаларды, құрамында қарапайым азот изотопы бар 14N бар ортаға ауыстырды. E.coli генерациясы уақытына сәйкес келетін (50мин 36ᵒС)—бір жасушалық бөлінуге және бір ДНҚ репликациясына қажетті уақыттың жылдамдығы өткеннен кейін,—жасуша үлгілерін таңдайды, олардан ДНҚ-ны бөледі және 20 сағ ішінде 40000g-та, цезий хлориді( CsCl) ерітіндісінде центрифугалайды(орталықтан тебу). Сонымен қатар ауыр CsCl молекулалары, пробирканың жоғарғы жағынан түбіне дейін өсетін тығыздық градиентін қалыптастыра отырып, тұна бастайды. ДНҚ тығыздығы CsCl ерітіндісінің тығыздығына тең деңгейге жиналады. Ультрафиолеттегі зерттеулер кезінде, орталықтан тепкіш пробиркадағы ДНҚ, тар жолақ түрінде болады. 15N және 14N орталарында өсірілген, жасушадан бөлінген ДНҚ жолақтарының орналасуы және алынған ақпараттардың талдануы 23.21.суретінде ұсынылған. Жартылай консервативті репликация сызбасы 23.22. суретіне қоса берілген. Бұл эксперименттер ДНҚ репликациясы жартылай консервативті тәсілмен болатындығын көрсетті.
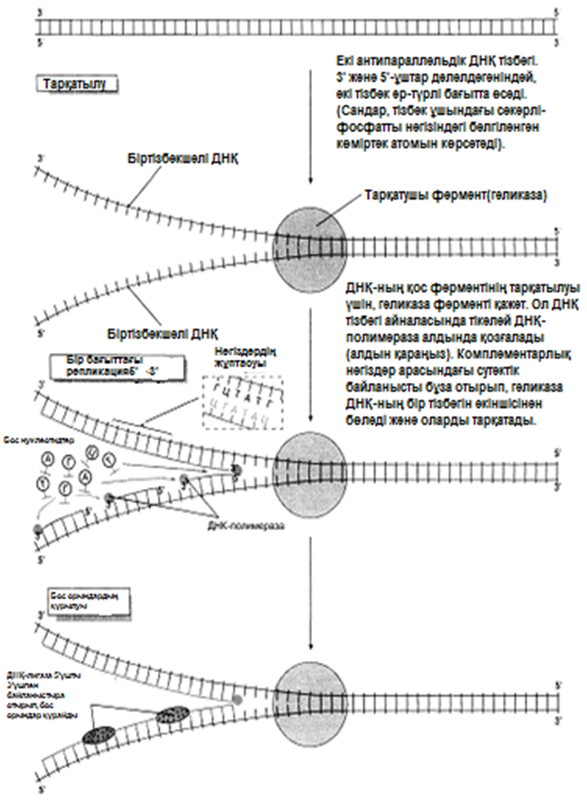
23.20. сур. ДНҚ репликациясы. Басты принциптер көрсетілген, оңайлатылған сызба. Репликацияда сызбада көрсетілмегенбасқа да ферменттер және молекулалар қатысады.
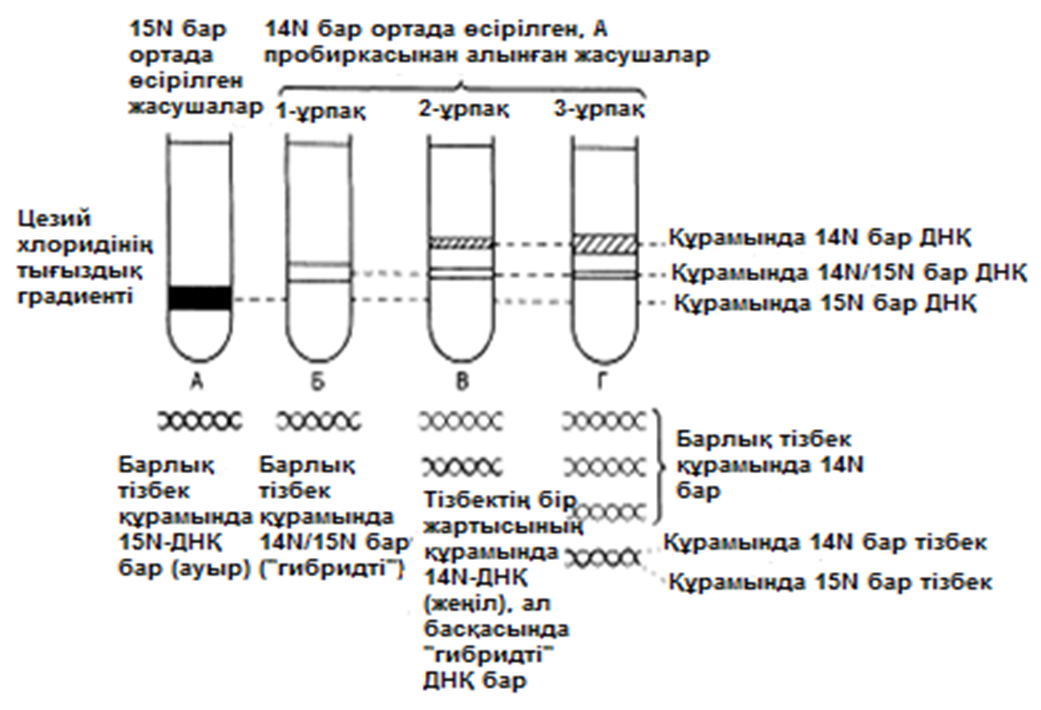
23.21.сур.Мезелсон мен Стальдың эксперименттернәтижәтижелері және олардың интерпретациясы. Орталықтан тепкіш пробиркадағы ДНҚ сызығының ені, әр түрлердегі ДНҚ молекуласының қатынас мөлшерін көрсетеді. В пробиркасындағы сызықтаренінің қатынасы 1:1, ал Г пробиркасында 3:1.
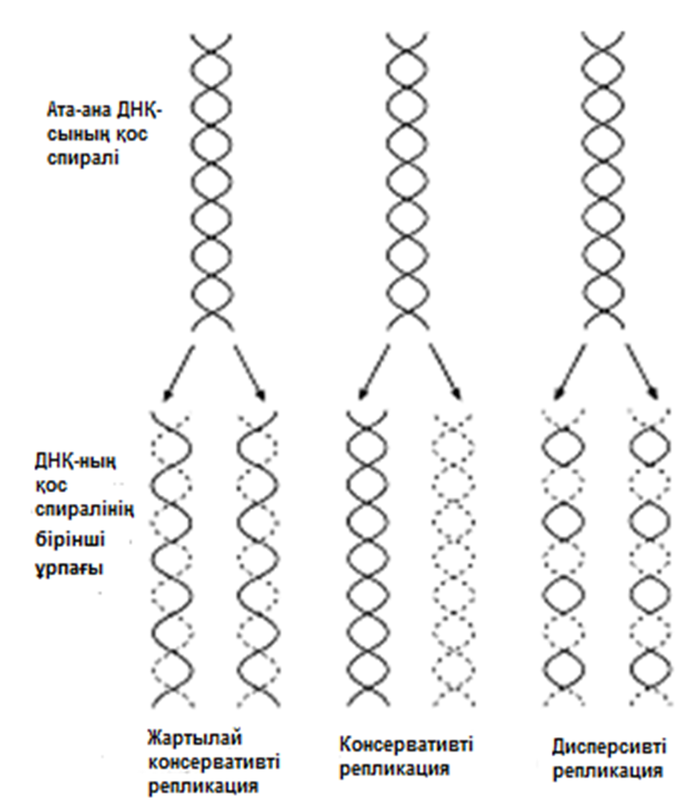
23.22.сур. ДНҚ репликациясының үш теориясын түсіндіретін сызба

23.7. Ген табиғаты
23.7.1. Ген дегеніміз не?
1866 жылы Мендель, ағзаның белгілері ол «элемент» деп атаған, тұқымқуалайтын бірліктермен анықталады деген болжам айтты. Кейіннен оларды ген деп атай бастады, және олардың гендерді ұрпақтан –ұрпаққа беретін хромосомаларда орналасқандығы көрсетілді. Сондықтан да Мендель, генді тұқымқуалаушылық бірлігі ретінде анықтауы мүмкін еді. Бұл түгелдей келісуге боларлық анықтама, бірақ бұл бізге геннің физикалық табиғаты туралы еш нәрсе айтпайды.
Төменде біз геннің физикалық табиғатын, оның екі анықтамасы нәтижесінде түсіндіруге тырысамыз.
РЕКОМБИНАЦИЯ БІРЛІГІ РЕТІНДЕГІ ГЕН. Морган өзінің Drosophila хромосомдық картасын құрастыру жұмыстарында, ген-кроссинговер нәтижесінде оған қосылатын аймақтардан бөлектенуі мүмкін, хромосоманың ең қысқа аймағы екендігін жазбаша таратты. Осы анықтамаға сәйкес, ген, ағзаның белгілі бір белгісін анықтайтын, әлдебір нақты хромосома кесіндісін көрсетеді.
ФУНКЦИЯ БІРЛІГІ РЕТІНДЕГІ ГЕН. Гендердің, ағзаның құрылымдық, физиологиялық және биохимиялық белгілерін анықтайтындығы белгілі болғандықтан, белгілі өнім синтезіне жағдай туғызатын, хромосоманың ең кіші аймағы ретінде, генді анықтау ұсынылды. Енді біз гендердің нәруыз синтезін код арқылы реттейтінін білеміз. Сондықтан да генді белгілі нәруызды код арқылы реттейтін ДНҚ аймағы ретінде анықтауға болады. Қанша дегенмен, кейбір нәруыздар бір емес, бірнеше полипептидтік тізбектен тұратындықтан, сондықтан да бір генге қарағанда, көбірек код арқылы реттелетіндіктен, бұл анықтаманы, генді нақтыланған полипепетидті код арқылы реттейтін ДНҚ аймағы деп атап, одан әрі растауға болады.
23.7.2. Генетикалық код— бұл негіздердің реттілігі
1953 жылы Уотсон мен Крик спиральді ДНҚ құрылымын жариялаған кезде, олар сонымен қатар, ұрпақтан-ұрпаққа берілетін және жасушалардың өміршеңдігін бақылайтын генетикалық ақпараттың, ДНҚ молекуласында, негіздердің реттілігі түрінде жатқандығы туралы да болжам айтқан. ДНҚ-ның нәруыз молекулалары синтезін код арқылы реттейтіндігі көрсетілгеннен кейін ДНҚ-дағы негіздердің реттілігі, нәруыз молекуларындағы аминқышқылдарының реттілігін, код арқылы реттеу тиістігі анықталды. Бұл негіздер мен аминқышқылдары арасындағы байланыс генетикалық код атымен белгілі. 1953 жылы шешілмеген мәселелер қатары қалды: оны түсіндіріп және ДНҚ-дағы негіздердің реттілігі, нәруыз молекуласындағы аминқышқылдар реттілігіне қандай тәсілмен ауысатындығын орнату керек болды.
23.7.3. Үштік код
ДНҚ молекуласы төрт түрдің негіздерінен салынған аденин(А), гуанин(Г), тимин(Т) және цитозин(Ц) (3.6. бөлім). Әр негіз нуклеотид бөлігін құрайды, ал нуклеотидтер жартылай нуклеотидті тізбекке байланысқан; олар өз атауларының бастапқы әрпімен белгіленеді және алфавиттің төрт әрпі, синтездеу нұсқаулығы, әр-түрлі нәруыз молекулаларының потенциалды шексіз санын жазуға мүмкіндік береді. Нәруыздар құрылған және ДНҚ құрамына кіретін негіздермен код арқылы реттелуі тиіс, 20 амнқышқылы бар. Егер де қандай да бір нәруыздың бірінші реттік құрылымындағы бір аминқышқылының орналасқан жері, бір негізді анықтайтын болса, онда бұл нәруыз құрамында тек төрт әр-түрлі аминқышқылы болар еді. Егер де әр аминқышқылы код арқылы реттелетін болса, онда мұндай код көмегімен 16 аминқышқылын анықтауға болатын еді.

Нәруыз молекуласына барлық 20 аминқышқылын қосу, тек қана үш негізден тұратын кодты қамтамасыз ете алады. Мұндай код жеткілікті мөлшерден де көп, негіздердің 64 сәйкестігін бере алады. Сондықтан да Уотсон және Крик, код үштік(триплоидты) болуы тиіс екендігін болжады.

Кейінірек кодтың шынында да триплоидты(үштік) екендігі, яғни әр аминқышқылдың үш негізбен код арқылы реттелетіндігі дәлелденді.
Кодтың триплоидтылығын(үштіктілігін) растайтын ақпараттар
Мұндай ақпараттар 1961 жылы Фрэнсис Крикпен берілген. Ол Т4 фагынан негіздердің қосылуынан немесе жойылуынан шақырылған мутациялар алды. Қандай да бір негіздің қосылуы немесе жойылуы(делеция), өзгеріс болған нүктеден кейін кодтың «оқылуын» өзгертеді(23.23. сур.). Мұндай өзгерістер нәтижесінде пайда болған мутацияны «санау жақтаушасының жылжуымен» бірге жүретін мутация деп атайды. Бұл мутация, бұрынғы аминқышқылдар реттілігімен нәруыз молекулаларының синтезін қамтамасыз ете алатын, негіздердің триплоидты(үштік) реттілігінің тууына мүмкіндік бермейді. Белгілі нүктелердегі бір негіздің қосылуы немесе жойылуы ғана негіздердің бастапқы реттілігіне әкелуі мүмкін. Мұндай қалпына келу Т4 эксперименттік фагтары арасында мутанттарының пайда болуын болдырмайтын еді. Бір негіздің қосылуы мутацияның (+) тәрізді, ал бір негіздің жойылуы мутацияның (-) тәрізді атауын алды. (+)(-) тәрізді мутациялар дұрыс санауды қалпына келтіреді. Қос мутанттарда да (++) немесе (--) кемшілігі бар нәруыздарды синтездейтін мутанттардың пайда болуына әкелетін, санау жақтаушасының жылжуы болады. Бірақ та (+++) немес (---) мутанттарында нәруыз синтезінің ешқандай өзгерісі бақыланған жоқ. Криктің ойынша, бұл, мұндай мутациялар, жақтаушаның жылжуын шақырмайтындығымен, ал тек нәруыз функциясына жиі әсер етпейтін, бір аминқышқылдың қосылуына немесе жойылуына ғана әкелетіндігімен түсіндіріледі. Бұл, кодтың бірден үш негізбен, яғни триплоидтармен(үштіктермен) саналатындығын білдіреді.
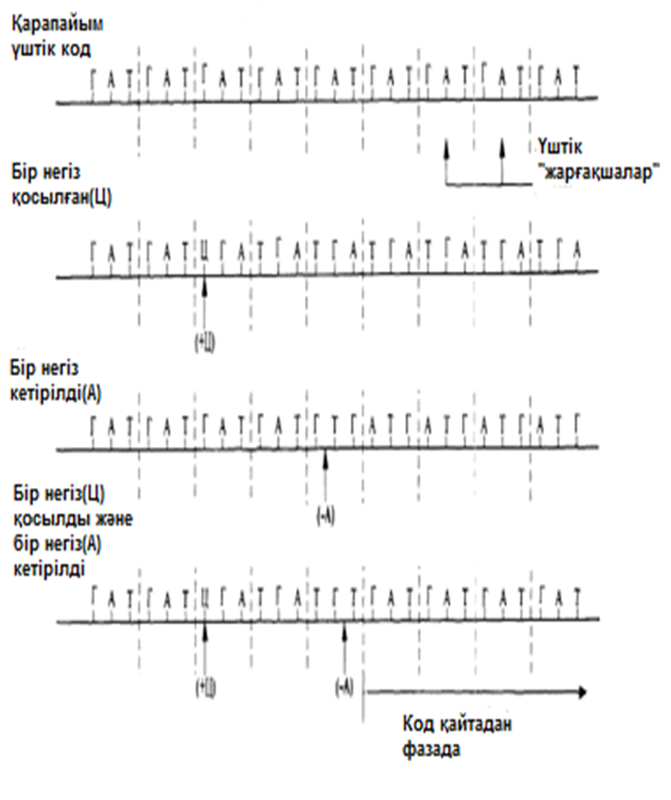
23.23.сур. Үштік кодтағы негіздердің қосылуы немесе кетірілуі нәтижесін түсіндіретін сызба. Ц негізін қосу, санау жақтаушасыныңжылжуына әкеледі, сондықтан бастапқы хабарлама ГАТ, ГАТ... , ТГА, ТГА... –ға айналады. А негізінің кетірілуі, бастапқы хабарлама ГАТ, ГАТ –ның АТ, АТГ..-ға ауысуына әкелетін, жақтаушаның жылжуын тудырады. Сызбада көрсетілген Ц негізінің қосылуы және А негізінің кетірілуі,бастапқы ГАТ, ГАТ... хабарламасын қалпына келтіреді. (По F. Н. С. Crick, The genetic code I, 1962, Scientific American Offprint, N123, Wm. Saunders and Co.)
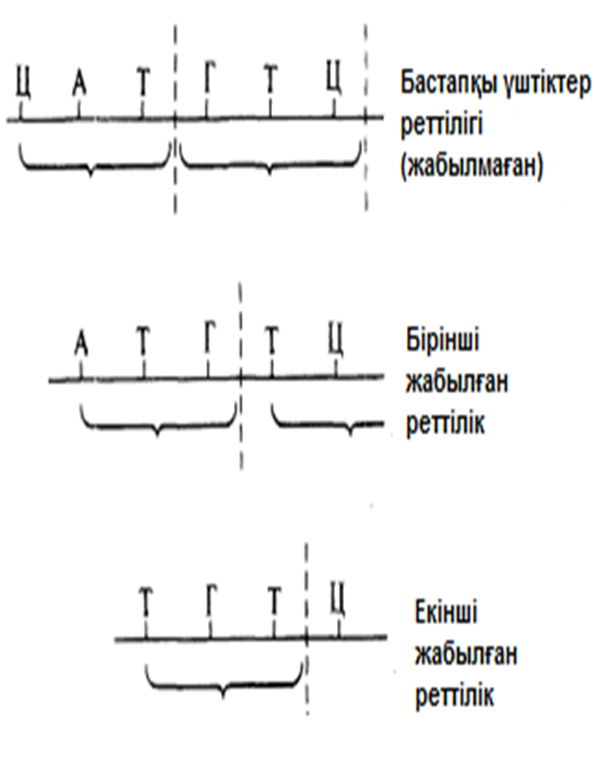
23.24. сур. Жабылған және жабылмаған кодтардағы үштіктер реттілігі.
Бұл эксперименттер триплоидтардың(үштіктердің) жабылмайтындығын, яғни әр негіздің тек бір триплоидқа тиесілі екендігін де көрсетті. Берілген триплоидқа(үштікке) кіретін негіздердің ешқайсысы, көршілес триплоидтың(үштіктің) бөлігі болып табылмайды.(23.24. сур.).

23.7.4. Кодтың мағынасын ашу
Кодтың триплоидтығы(үштігі) орнатылғаннан кейін, аминқышқылдардың әр ақпаратын қандай триплоидтардың(үштіктердің) код арқылы реттейтінін білу, басқаша сөзбен айтқанда—кодтың мағынасын ашу қалды. Эксперименттік процедураларда сонымен қатар қолданылғандарды білу үшін, үштік кодтың нәруыз молекуласына аударылуына көмектесетін механизмнің жалпы ерекшеліктерін елестету керек.
Нәруыз синтезінде бір-бірімен әрекеттесетін екі түрлі нуклеин қышқылдары қатысады: дезоксирибонуклеин қышқылы(ДНҚ) және рибонуклеин қышқылы (РНҚ). РНҚ-ның басты үш түрі бар:ақпараттық немесе матрицалық РНҚ (мРНҚ), рибосомдық РНҚ(рРНҚ) және транспорттық РНҚ(тРНҚ). ДНҚ негіздерінің реттілігі, содан соң ядродан цитоплазмаға түсетін матрицалық РНҚ тізбегінде жазылады (транскрипцияланады). Мұнда ол тізбектер рибосомаларға бекітіледі және мРНҚ реттілігі аминқышқылдар реттілігіне ауысады. Әр аминқышқылы, комплементарлық мРНҚ негізі үштігіне бекітілетін, сәйкес тРНҚ- мен байланысады. Осының нәтижесінде бір-бірінің қасында кенеттен пайда болған амнқышқылдар, полипептидтік тізбек құрып бірігеді. Осылайша, нәруыздық синтезге ДНҚ, мРНҚ, рибосомалар, тРНҚ, аминқышқылдар, энергия қоры ретінде АТФ, сонымен қатар бұл процесстің әр кезеңін катализдейтін әр-түрлі ферменттер және кофакторлар керек.
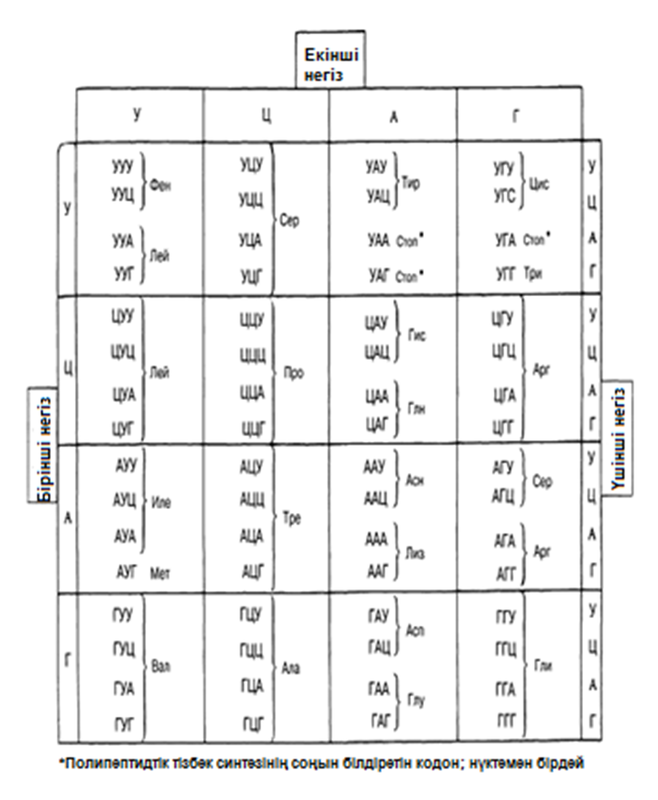
Ниренберг ХХ ғаысрдың 50-жылдарының соңында табылған бұл ақпараттарды және әр-түрлі әдістерді, кодтың мағынасын ашу мақсатында қойылған эксперименттер қатарын жасау үшін қолданған. Оның эксперименттерінің мәні, алдын-ала белгілі негіздер реттілігі бар мРНҚ-ны қолдану үшін, осы мРНҚ-ның қатысуында синтезделетін, полипептидтік тізбектегі аминқышқылдар реттілігін анықтауға әкелді. Ниренберг бірнеше рет қайталанатын үштік УУУ-дан құралатын мРНҚ синтездеуге қол жеткізді. Бұл жартылай урацилді қышқыл(жартылай (У)) деп аталған байланыс, код ретінде қолданылады. 20 пробирканың әрқайсысына құрамында рибосомалар, тРНҚ, АТФ, ферменттер және қандай-да бір таңбаланған аминқышқыл бар жасушасыз экстракт E.coli орналастырды. Содан соң әр пробиркаға жартылай(У) қосқан және полипептид синтезі жүруі үшін, біршама уақытқа қойған. Пробирканың құрамындағылардың анализі, құрамында фенилаланин аминқышқылы бар пробиркада ғана полипептид түзілгенін көрсетті. Бұл генетикалық кодтың мағынасын ашудағы бірінші қадам болды. Ниренберг, мРНҚ құрамына кіретін негіздер үштігі немесе кодон УУУ-дың, фенилалатиннің полипептидтік тізбектегі орнын анықтайтындығын көрсетті. Содан соң Ниренберг және оның қызметкерлері, барлық 64 мүмкін кодондарға сәйкес келетін жартылай нуклеотидтердің синтетикалық молекулаларын жасаумен айналысты және 1964 жылға қарай барлық 20 аминқышқылы үшін кодтардың мағынасын ашты (23.4. кестесі).
23.4. кестесі. Үштік кодтағы негіздер реттілігі және үштіктермен код арқылы реттелетін аминқышқылдар.
Ескерту. ДНҚ-дағы емес, мРНҚ-дағы кодондар, яғни, негіздер реттілігі ұсынылған. ДНҚ-ның генетикалық коды комплементарлы негіздерден тұрады, ал У мұнда Т м-мен ауыстырылған.
23.7.5. Генетикалық кодтың сипаттамалары
ҮШТІКТІГІ
Бұған дейін көрсетілгендей, генетикалық код— бұл үштік код: ДНҚ молекуласындағы үш негіз кез-келген нәруыз молекуласындағы бір аминқышқылды код арқылы реттейді. ДНҚ-коды басында, осы ДНҚ-мен комплементарлы РНҚ матрицаға транскрипцияланады. мРНҚ-ның комплементарлық үштігін кодон деп атайды. Әр кодон үш негіз ұзындығына тең және бір аминқышқылды код арқылы реттейді. РНҚ кодонын ДНҚ негізінің комплементарлы үштігіне 23.5. кестесімен сәйкес аударса, әр аминқышқыл үшін ДНҚ кодын алуға болады.
23.5. кестесі. ДНҚ және РНҚ негіздері арасындағы комплементарлылық


Туынды код
23.4. кестесінде генетикалық код кодондары мысалға келтірілді. Кестеде көрсетілгендей, кейбір аминқышқылдар бір емес, бірнеше кодондармен реттеледі. Мұндай кодтарды туынды код деп атайды. Бұл код анализі сондай-ақ, көптеген аминқышқылдар үшін, көргеніміз бойынша, тек қана кодонның бастапқы екі әрпінің ғана маңызы бар екендігін көрсетеді.
Код құрамында тыныс белгілері бар
23.4. кестесінде ұсынылған кодондар арасындағы үшеуі «нүкте» ретінде қызмет атқарады, яғни хабарламаның аяқталғанын білдіреді. Мысал ретінде УАА үштігі қызмет атқарады. Мұндай кодондарды кейде «нонсенс-кодондар» деп атайды; олар аминқышқылдардың біреуін де код арқылы реттемейді. Көргеніміз бойынша, бұл кодондар берілген геннің соңын білдіреді, яғни трансляция кезінде полипептидтік тізбек синтезін тоқтататын «стоп-сигналдар» ретінде қызмет атқарады.
АУГ (метионин) тәрізді кейбір басқа кодондар, полипептидтік тізбектегі трансляция (ауысу) кіріспесіне нұсқай отырып, «старт-сигналдар» ретінде қызмет атқарады.
Код әмбебап болып табылады
Генетикалық кодтың елеулі ерекшеліктерінің бірі, бұл, көргеніміз бойынша, оның әмбебаптығында. Барлық тірі ағзаларда барлығында да бірдей 20 аминқышқыл және бірдей бес негіз болады (А, Г, Т, Ц және У).
Қазіргі уақытта молекулалық биологиядағы жетістіктер, бүтін гендер және бүтін ағзалар үшін негіздердің реттілігін анықтау мүмкін болатындай деңгейге жетті. Бүтін генетикалық кодының мағынасын ашу мүмкіндігі туған бірінші ағза, вирустардың бірі— фаг ϕХ174 болды. Бұл фагта бар болғаны 10 ген бар, ал оның толық генетикалық коды 5386 негізден тұрады. Бұл негіздердің реттілігін— алғаш рет нәруыздардың біреуінде аминқышқылдардың реттілігін ашқан зерттеуші— Фред Сенджер орнатты. Ол, бұл іргелі жаңалықтарының әрқайсысы үшін Нобель сыйлығынан алды. Енді, гендік инженерияда қолданыс табатын бүтін гендерді синтездеу мүмкін болды. ХХІ ғасырдың ең басында «Адам геномы» жобасы айналасында, ұзындығы, бағалау бойынша 3000 мм негіздер жұбына тең, адамның толық генетикалық кодының мағынасын ашу мүмкін болатындығын күту керек.(Геном— бұл берілген ағзаның барлық ДНҚ-сы. Қазіргі кезде E.coli, шыбын(Drosophila), нематодтардың(дөңгелек құрт) біреуі және лабороториялық тышқан геномдарының мағынасын ашу жөнінде жұмыстар жүргізіліп жатыр.
Резюме
Төмендегі қысқашада генетикалық кодтың негізгі ерекшеліктері тұжырымдалған.
23.8. Нәруыз синтезі
«ДНҚ РНҚ тудырады, ал РНҚ нәруыз тудырады»
Алдыңғы бөлімдерде айтылғандардың барлығынан, ДНҚ-ның тікелей бақылауында синтезделетін бірегей молекулалар — нәруыз екенін көруге болады. Нәруыздар кератин және коллаген тәрізді құрылымды, немесе инсулин, фибриноген және ең бастысы, жасушалық метаболизмнің реттелуіне жауапты ферменттер тәрізді функциональды бола алады. Берілген жасуша құрамындағы ферменттер жиынтығы ғана, оның қай жасуша түріне жататындығын анықтайды. Осы тәсілмен ДНҚ, жасушаның өмір сүру қабілетін бақылайды.
Ферменттер синтезіне және барлық басқа нәруыздарға қажетті «нұсқаулар», толықтай дерлік ядродаорналасқан ДНҚ-да орныққан. Бірақ, ХХ ғасырдың 50-жылдарының басында көрсетілгендей, синтез нақты цитоплазмада жүреді және оған рибосомалар қатысады. Генетикалық ақпаратты ядродан цитоплазмаға ауыстыратын, қандай да бір механизм болуы керек екендігі анықталды. 1961 жылы екі француз биохимигі Жакоб және Моно, өздері матрицалық РНҚ(мРНҚ) деп атаған ерекше РНҚ формасы бар екендігін жазбаша жариялады. Олардың ойлары дұрыс болып шықты. Нәруыз синтезі процесінде болатын оқиғалар реттелігі мына сөйлемде тұжырымдалған: «ДНҚ РНҚ тудырады, ал РНҚ нәруыз тудырады».
23.8.1. РНҚ рөлі
РНҚ барлық тірі жасушалар құрамында біртізбекшелі молекула түрінде болады. Ол ДНҚ-дан , құрамында пентоза ретінде рибоза (дезоксирибоза орнына) , ал пиримидиндік негіздердің бірі ретінде— урацил (тимин орнына) бар екендігімен ерекшеленеді. Жасуша құрамындағы РНҚ анализі, нәруыз молекулалары синтезіне қатысатын РНҚ-ның үш түрі бар екендігін көрсетті. Олар матрицалық, немесе ақпараттық РНҚ(мРНҚ), транспорттық РНҚ(тРНҚ) және рибосомдық РНҚ(рРНҚ). Барлық үш РНҚ тікелей ДНҚ-да синтезделеді, ал әр жасушадағы РНҚ мөлшері, осы жасушадан түзілітін нәруыз мөлшеріне тікелей байланысты болады.
23.8.2. Матрицалық РНҚ
Анализдердің көрсетуі бойынша, мРНҚ, барлық РНҚ жасушаларының 3-5% құрайды. Бұл, транскрипция деп аталатын процессте ДНҚ тізбегінің біреуінде қалыптасатын біртізбекшелі молекула. мРНҚ синтезі кезінде тек қана ДНҚ молекуласының бір тізбегі көшірмеленеді. мРНҚ синтезі кейінірек сипатталатын болады. мРНҚ-дағы негіздердің реттілігі, сәйкес келетін ДНҚ тізбегінің комплементарлы көшірмесін құрайды; оның ұзындығы, ол код арқылы реттейтін полипептидтік тізбек ұзындығына байланысты өзгеріп тұрады. мРНҚ-ның көп бөлігі жасушада тек қысқа уақыт ішінде болады. Бактерияларда бұл бар болғаны бірнеше минут болуы мүмкін, онда дамып жатырған эритроциттерде мРНҚ, гемоглобин синтезі үшін матрица ретінде бірнеше күндер ішінде қызмет ете алады.
28.8.3. Рибосомдық РНҚ
Рибосомдық РНҚ, шамамен, барлық хасуша РНҚ-сының 80%-ын құрайдв. Ол ядрошық ұйымдастырушысы атымен белгілі ядрошық аймағында орналасқан бірнеше хромосомалар ДНҚ-сындағы гендермен код арқылы реттеледі. рРНҚ-дағы негіздер реттілігі барлық ағзаларда ұқсас— бактериядан бастап, жоғары өсімдіктер мен жануарларға дейін. рРНҚ, нәруыз молекулаларымен бірге рибосомадеп аталатын жасуша органеллаларын түзе отырып байланысатын жер— цитоплазма құрамында болады (5.10.4 бөлім)
Рибосомаларда нәруыз синтезі болады. Бұл жерде мРНҚ-да орныққан «код», полипептидтік тізбектің аминқышқылдық реттілігіне тасымалданады.
28.8.4. Транспорттық РНҚ
Транспорттық РНҚ-ның(тРНҚ) бар екендігі Крикпен жазбаша жарияланды және 1855 жылы Хоглендпен көрсетілді. Әр аминқышқылда тРНҚ молекулаларының жеке отбасысы болады. Олар цитоплазма құрамындғы аминқышқылдарды рибосомаға жеткізеді. Осылайша, тРНҚ, мРНҚ-дағы үштік код және полипептидтік тізбектегі аминқышқылдық реттілік арасында, аралық молеула ретінде қызмет атқарады. тРНҚ үлесіне, шамамен, барлық жасушалық РНҚ-ның 15%-ы келед; Ең қысқа полинуклеотидтік тізбек, осы РНҚ-да— оған орташа есеппен 80 нуклеотид кіреді. Әр жеке жасуша құрамында 20-дан көбірек тРНҚ молекулалары болады(қазір 60-ы анықталған). Барлық тРНҚ молекулалары ұқсас негізгі құрылымдыболады (23.25. сур.қар.).
тРНҚ молекуласының 5’-соңында әрқашан гуанин, ал 3’-соңында— ЦЦА негізінің реттілігі болады. Молекуланың қалған бөлігіндегі нуклеотидтер реттілігі өзгеріп тұрады және инозин мен псевдоурацил тәрізді «ерекше» негіздер болуы мүмкін. Антикадон үштігіндегі негіздер реттілігі (23.25.сур.) берілген тРНҚ молекуласы тасымалдайтын аминқышқылға қатаң сәйкес келеді. Әр аминқышқыл өзіне тән тРНҚ-ның біреуіне, аминоацил-тРНҚ-синтетаза ферментінің қалыптасуымен бірігеді. Нәтижесінде, алда көршіліс аминқышқылмен байланыс пайда болуы үшін, ЦЦА үштігіндегі соңғы нуклеотид А және аминқышқыл арасындағы байланыс энергиясы жеткілікті болатын, аминоацил-тРНҚ ретінде белгілі тРНҚ-аминқышқылының кешені пайда болады. Осылайша полипептидтік тізбек синтезделеді.
23.4. кестесінде генетикалық код кодондары мысалға келтірілді. Кестеде көрсетілгендей, кейбір аминқышқылдар бір емес, бірнеше кодондармен реттеледі. Мұндай кодтарды туынды код деп атайды. Бұл код анализі сондай-ақ, көптеген аминқышқылдар үшін, көргеніміз бойынша, тек қана кодонның бастапқы екі әрпінің ғана маңызы бар екендігін көрсетеді.
Код құрамында тыныс белгілері бар
23.4. кестесінде ұсынылған кодондар арасындағы үшеуі «нүкте» ретінде қызмет атқарады, яғни хабарламаның аяқталғанын білдіреді. Мысал ретінде УАА үштігі қызмет атқарады. Мұндай кодондарды кейде «нонсенс-кодондар» деп атайды; олар аминқышқылдардың біреуін де код арқылы реттемейді. Көргеніміз бойынша, бұл кодондар берілген геннің соңын білдіреді, яғни трансляция кезінде полипептидтік тізбек синтезін тоқтататын «стоп-сигналдар» ретінде қызмет атқарады.
АУГ (метионин) тәрізді кейбір басқа кодондар, полипептидтік тізбектегі трансляция (ауысу) кіріспесіне нұсқай отырып, «старт-сигналдар» ретінде қызмет атқарады.
Код әмбебап болып табылады
Генетикалық кодтың елеулі ерекшеліктерінің бірі, бұл, көргеніміз бойынша, оның әмбебаптығында. Барлық тірі ағзаларда барлығында да бірдей 20 аминқышқыл және бірдей бес негіз болады (А, Г, Т, Ц және У).
Қазіргі уақытта молекулалық биологиядағы жетістіктер, бүтін гендер және бүтін ағзалар үшін негіздердің реттілігін анықтау мүмкін болатындай деңгейге жетті. Бүтін генетикалық кодының мағынасын ашу мүмкіндігі туған бірінші ағза, вирустардың бірі— фаг ϕХ174 болды. Бұл фагта бар болғаны 10 ген бар, ал оның толық генетикалық коды 5386 негізден тұрады. Бұл негіздердің реттілігін— алғаш рет нәруыздардың біреуінде аминқышқылдардың реттілігін ашқан зерттеуші— Фред Сенджер орнатты. Ол, бұл іргелі жаңалықтарының әрқайсысы үшін Нобель сыйлығынан алды. Енді, гендік инженерияда қолданыс табатын бүтін гендерді синтездеу мүмкін болды. ХХІ ғасырдың ең басында «Адам геномы» жобасы айналасында, ұзындығы, бағалау бойынша 3000 мм негіздер жұбына тең, адамның толық генетикалық кодының мағынасын ашу мүмкін болатындығын күту керек.(Геном— бұл берілген ағзаның барлық ДНҚ-сы. Қазіргі кезде E.coli, шыбын(Drosophila), нематодтардың(дөңгелек құрт) біреуі және лабороториялық тышқан геномдарының мағынасын ашу жөнінде жұмыстар жүргізіліп жатыр.
Резюме
Төмендегі қысқашада генетикалық кодтың негізгі ерекшеліктері тұжырымдалған.
- Аминқышқылдардың полипептидтік тізбекке қосылуын анықтайтын код ретінде, ДНҚ-ның полинуклеотидтік тізбегіндегі негіздер үштігі қызмет атқарады.
- Код әмбебап болып табылады: барлық ағзаларда бірдей үштіктер бірдей аминқышқылдарды код арқылы реттейді (митохондриялық ДНҚ-ның және кейбір ежелгі бактериялардың бірнеше үштік кодтарының әмбебап кодтап айырмашалаға бар).
- Туынды код: берілген аминқышқыл бір үштікке қарағанда көбірек үштікпен код арқылы реттеле алады.
- Жабылмайтын кодтар: мысалы АУГАГЦГЦА нуклеотидтерінен басталатын мРНҚ реттілігі, АУГ/УГА/ГАГ...(екі негізбен жабылу) немесе АУГ/ГАГ/ГЦГ...(бір негізбен жабылу) ретінде саналмайды. (Бірақ, бірнеше гендермен жабылу кейбір ағзаларда табылған, мысалы бактериофаг ϕХ174. Мұндай жағдайлар, өте сирек және өте аз гендер санындағы ДНҚ-ны үнемдеумен түсіндірілуі мүмкін).
23.8. Нәруыз синтезі
«ДНҚ РНҚ тудырады, ал РНҚ нәруыз тудырады»
Алдыңғы бөлімдерде айтылғандардың барлығынан, ДНҚ-ның тікелей бақылауында синтезделетін бірегей молекулалар — нәруыз екенін көруге болады. Нәруыздар кератин және коллаген тәрізді құрылымды, немесе инсулин, фибриноген және ең бастысы, жасушалық метаболизмнің реттелуіне жауапты ферменттер тәрізді функциональды бола алады. Берілген жасуша құрамындағы ферменттер жиынтығы ғана, оның қай жасуша түріне жататындығын анықтайды. Осы тәсілмен ДНҚ, жасушаның өмір сүру қабілетін бақылайды.
Ферменттер синтезіне және барлық басқа нәруыздарға қажетті «нұсқаулар», толықтай дерлік ядродаорналасқан ДНҚ-да орныққан. Бірақ, ХХ ғасырдың 50-жылдарының басында көрсетілгендей, синтез нақты цитоплазмада жүреді және оған рибосомалар қатысады. Генетикалық ақпаратты ядродан цитоплазмаға ауыстыратын, қандай да бір механизм болуы керек екендігі анықталды. 1961 жылы екі француз биохимигі Жакоб және Моно, өздері матрицалық РНҚ(мРНҚ) деп атаған ерекше РНҚ формасы бар екендігін жазбаша жариялады. Олардың ойлары дұрыс болып шықты. Нәруыз синтезі процесінде болатын оқиғалар реттелігі мына сөйлемде тұжырымдалған: «ДНҚ РНҚ тудырады, ал РНҚ нәруыз тудырады».
23.8.1. РНҚ рөлі
РНҚ барлық тірі жасушалар құрамында біртізбекшелі молекула түрінде болады. Ол ДНҚ-дан , құрамында пентоза ретінде рибоза (дезоксирибоза орнына) , ал пиримидиндік негіздердің бірі ретінде— урацил (тимин орнына) бар екендігімен ерекшеленеді. Жасуша құрамындағы РНҚ анализі, нәруыз молекулалары синтезіне қатысатын РНҚ-ның үш түрі бар екендігін көрсетті. Олар матрицалық, немесе ақпараттық РНҚ(мРНҚ), транспорттық РНҚ(тРНҚ) және рибосомдық РНҚ(рРНҚ). Барлық үш РНҚ тікелей ДНҚ-да синтезделеді, ал әр жасушадағы РНҚ мөлшері, осы жасушадан түзілітін нәруыз мөлшеріне тікелей байланысты болады.
23.8.2. Матрицалық РНҚ
Анализдердің көрсетуі бойынша, мРНҚ, барлық РНҚ жасушаларының 3-5% құрайды. Бұл, транскрипция деп аталатын процессте ДНҚ тізбегінің біреуінде қалыптасатын біртізбекшелі молекула. мРНҚ синтезі кезінде тек қана ДНҚ молекуласының бір тізбегі көшірмеленеді. мРНҚ синтезі кейінірек сипатталатын болады. мРНҚ-дағы негіздердің реттілігі, сәйкес келетін ДНҚ тізбегінің комплементарлы көшірмесін құрайды; оның ұзындығы, ол код арқылы реттейтін полипептидтік тізбек ұзындығына байланысты өзгеріп тұрады. мРНҚ-ның көп бөлігі жасушада тек қысқа уақыт ішінде болады. Бактерияларда бұл бар болғаны бірнеше минут болуы мүмкін, онда дамып жатырған эритроциттерде мРНҚ, гемоглобин синтезі үшін матрица ретінде бірнеше күндер ішінде қызмет ете алады.
28.8.3. Рибосомдық РНҚ
Рибосомдық РНҚ, шамамен, барлық хасуша РНҚ-сының 80%-ын құрайдв. Ол ядрошық ұйымдастырушысы атымен белгілі ядрошық аймағында орналасқан бірнеше хромосомалар ДНҚ-сындағы гендермен код арқылы реттеледі. рРНҚ-дағы негіздер реттілігі барлық ағзаларда ұқсас— бактериядан бастап, жоғары өсімдіктер мен жануарларға дейін. рРНҚ, нәруыз молекулаларымен бірге рибосомадеп аталатын жасуша органеллаларын түзе отырып байланысатын жер— цитоплазма құрамында болады (5.10.4 бөлім)
Рибосомаларда нәруыз синтезі болады. Бұл жерде мРНҚ-да орныққан «код», полипептидтік тізбектің аминқышқылдық реттілігіне тасымалданады.
28.8.4. Транспорттық РНҚ
Транспорттық РНҚ-ның(тРНҚ) бар екендігі Крикпен жазбаша жарияланды және 1855 жылы Хоглендпен көрсетілді. Әр аминқышқылда тРНҚ молекулаларының жеке отбасысы болады. Олар цитоплазма құрамындғы аминқышқылдарды рибосомаға жеткізеді. Осылайша, тРНҚ, мРНҚ-дағы үштік код және полипептидтік тізбектегі аминқышқылдық реттілік арасында, аралық молеула ретінде қызмет атқарады. тРНҚ үлесіне, шамамен, барлық жасушалық РНҚ-ның 15%-ы келед; Ең қысқа полинуклеотидтік тізбек, осы РНҚ-да— оған орташа есеппен 80 нуклеотид кіреді. Әр жеке жасуша құрамында 20-дан көбірек тРНҚ молекулалары болады(қазір 60-ы анықталған). Барлық тРНҚ молекулалары ұқсас негізгі құрылымдыболады (23.25. сур.қар.).
тРНҚ молекуласының 5’-соңында әрқашан гуанин, ал 3’-соңында— ЦЦА негізінің реттілігі болады. Молекуланың қалған бөлігіндегі нуклеотидтер реттілігі өзгеріп тұрады және инозин мен псевдоурацил тәрізді «ерекше» негіздер болуы мүмкін. Антикадон үштігіндегі негіздер реттілігі (23.25.сур.) берілген тРНҚ молекуласы тасымалдайтын аминқышқылға қатаң сәйкес келеді. Әр аминқышқыл өзіне тән тРНҚ-ның біреуіне, аминоацил-тРНҚ-синтетаза ферментінің қалыптасуымен бірігеді. Нәтижесінде, алда көршіліс аминқышқылмен байланыс пайда болуы үшін, ЦЦА үштігіндегі соңғы нуклеотид А және аминқышқыл арасындағы байланыс энергиясы жеткілікті болатын, аминоацил-тРНҚ ретінде белгілі тРНҚ-аминқышқылының кешені пайда болады. Осылайша полипептидтік тізбек синтезделеді.
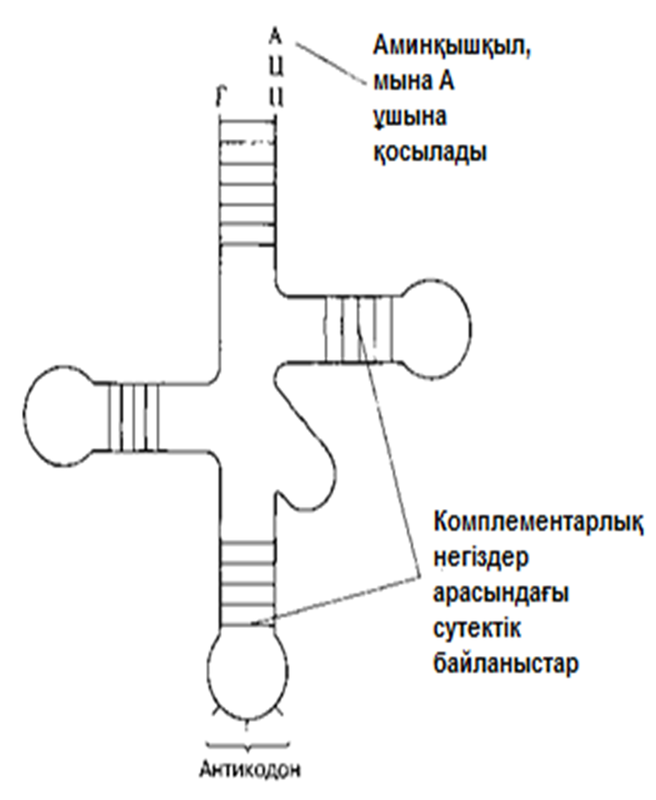
23.25.сур. Транспорттық РНҚ (тРНҚ) құрылымының ұсынылған модельдерінің бірі. Молекула 80 нуклеотидтен тұрады, бірақ негіздердің комплементарлық байланысуы нәтижесінде түзілген жұптар саны тек қана 21.
23.8.5. Резюме
Нәруыз синтезі процесі 23.26. суретінде сызба түрінде ұсынылған екі қадамнан тұрады.
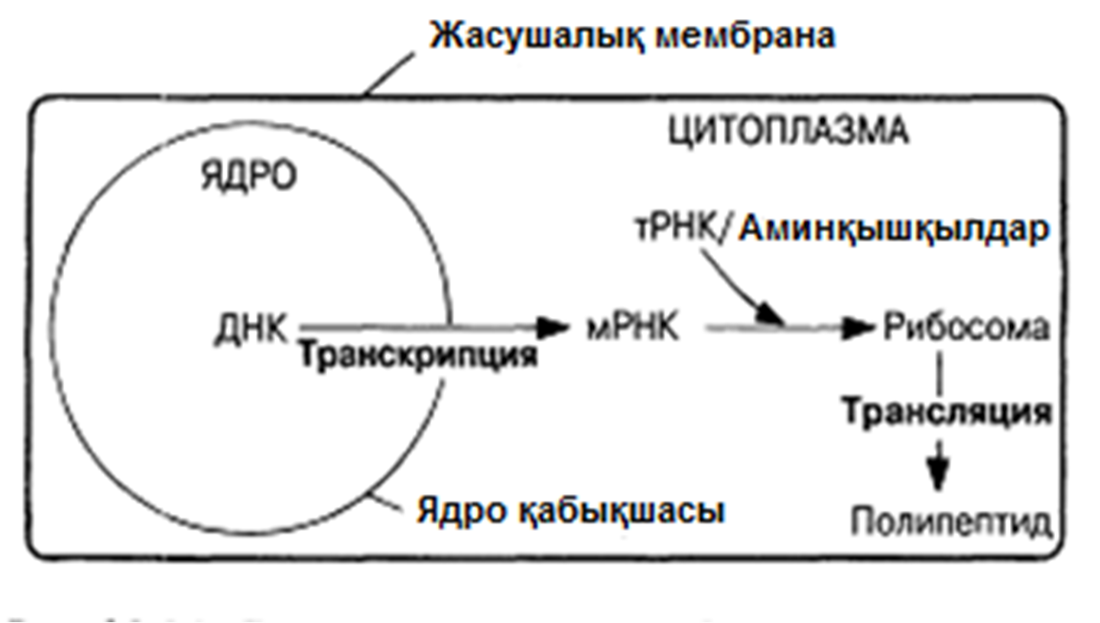
23.26. сур. Нәруыз синтезінің басты кезеңдерінің сызбасы
1. Транскрипция— мРНҚ-ның ДНҚ-ға синтезі. Сонымен қатар, ДНҚ-ның кейбір аймағы (ген) мРНҚ түзілуімен көшірмеленеді.
2. Трансляция— мРНҚ молекуласындағы негіздер реттілігінің, нәруыз молекуласындағы аминқышқылдар реттілігіне ауысуы.
23.8.6. Транскрипция
Транскрипция деп, ДНҚ тізбегі аймағыныңі біреуіндегі негіздер реттілігінің, мРНҚ негізінің комплементарлық реттілігіне «көшіріліп жазылуы» көмегімен жүретін процессті айтады. ДНҚ-ның қос спиралі, екі жалғыз ДНҚ тізбектері негіздерінің арасындағы салыстырмалы әлсіз сутектік байланыстардың үзілуі нәтижесінде тарқатылады. Бұл тізбектің біреуі ғана, мРНҚ-ның комплементарлы жалғыз тізбегі түзілуі үшін матрица ретінде таңдалуы мүмкін. Бұл молекула, ДНҚ және РНҚ негіздерінің жұптасу ережелерімен сәйкес болғандағы РНҚ-полимеразалары әсерінен, еркін рибонуклеотидтердің бір-бірімен байланысуы нәтижесінде түзіледі (23.5. кестесі және 23.27 суреті).
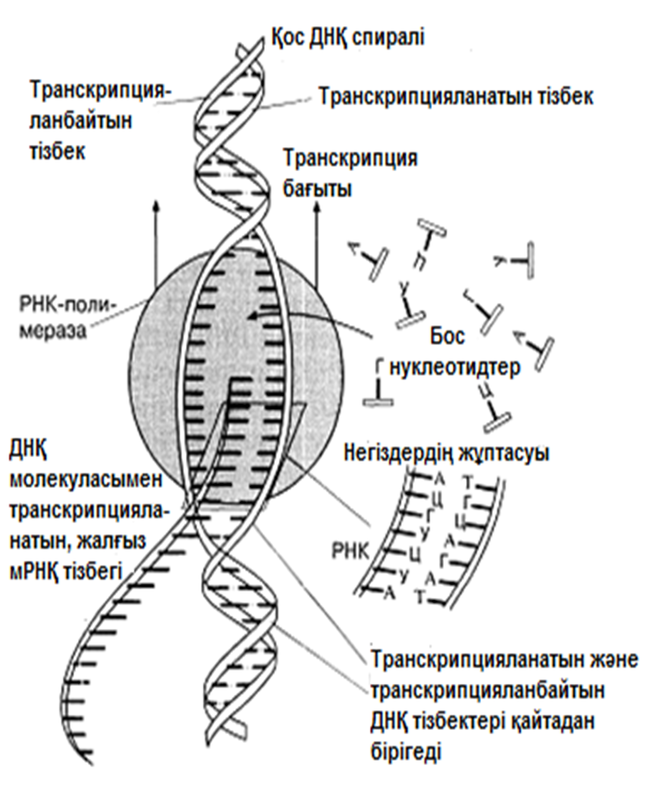
23.27.сур. Транскрипция механизмінің сызбасы. РНҚ-полимераза болғанда қос ДНҚ спиралі, комплементарлы негіздер арасындағы сутектік байланыстың үзілуі нәтижесінде тарқатылады.Бос рибонуклеотидтерден түзілген жалғыз ДНҚ тізбегінде, мРНҚ-ный полинуклеотидтік тізбегі пайда құрылады. мРНҚ негіздері ДНҚ-ның комплементарлы негіздеріне қарсы орналасады. ( Е. J. Ambrose, D. М. Easty, Cell Biology, 1977, 2nd ed. Nelson. бойынша)
РНҚ негізіндегі ДНҚ негіздерінің транскрипциясы дәл қандай тәсілмен жүретіндігі, тек қана бір нуклеотид түрінен— тимидиннен(ТТТ...) тұратын синтетикалық ДНҚ тәжірибелерінде көрсетілді. Бұл ДНҚ-ны құрамында ДНҚ-полимераза және барлық 4 нуклеотид(А, У, Г және Ц) бар жасушасыз жүйеге енгізгенде, құрамында тек қана комплементарлы адионуклеотидтер бар мРНҚ синтезделді.
Синтезделген мРНҚ молекулалары ядродан ядролық тесіктер арқылы шығады және генетикалық ақпаратты рибосомаларға апарады. Транскрипция процесінде мРНҚ молекулаларының жеткілікті мөлшері түзілгеннен кейін, РНҚ-полимераза ДНҚ-ны қалдырып кетеді («замок» бекітіледі)— қос спираль қалпына келтіріледі.
23.8.7. Трансляция
Трансляция деп РНҚ молекуласындағы негіздер реттілігі, полипептидтік тізбектегі аминқышқылдар реттілігіне тасымалдануы көмегімен жүретін процессті атайды. Бұл процесс рибосомаларда жүреді. Бірнеше рибосома мРНҚ түрінде, полирибосома немесе полисома деп аталатын құрылым түзе отырып бірігеді. Полисомаларды электрондық микроскоп көмегімен көруге болады (23.28. сур.). Полисомдық ұйымның артықшылығы, бұл кезде, бір уақытта бірнеше полипептидтердің синтезі мүмкін болатындығында (23.8.3. бөлім). Әр рибосома екі суббірліктен тұрады—үлкен және кіші-және пішіні бойынша саңырауқұлақты еске түсіреді (5.27. бөлім). мРНҚ-ның алғашқы екі кодоны (қосындысы 6 негіз) рибосомаға кіреді, 23.29.А. суретінде көрсетілгендей. Бірінші кодон, құрамында оған комплементарлы антикодон бар және синтезделіп жатқан полипептидтің бірінші аминқышқылын (әдетте, бұл, метионин) тасмалдайтын аминоацил-тРНҚ молекуласын байланыстырады. Содан соң екінші кодон, өзіне,құрамында оған комплементарлы антикодоны бар амиоацил-тРНҚ молекуласын қосып алады (23.29.Ажәне Б сур.). Рибосома функциясы, трансляция процесіне қатысатын мРНҚ, тРНҚ және ферменттерді, көршілес аминқышқылдар арасында пептидтік байланыс түзілгенге дейін қажетті жағдайда ұстап тұруында болып табылады.
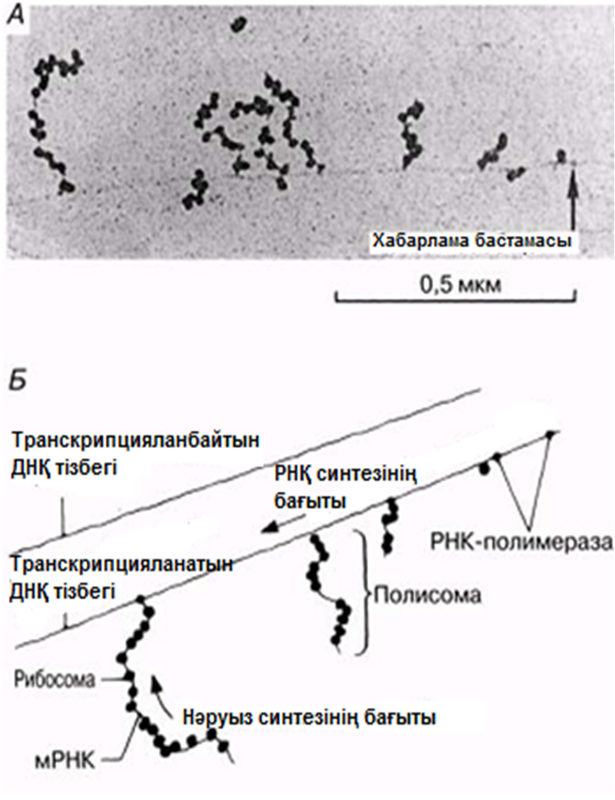
23.28.сур. Бактериядағы транскрипция және полисома түзілуі процесстері. Қанша дегенмен, бактерияда ядро болмайтындығына назар аударыңыз.Олар да РНҚ ДНҚ-дан бөлінбеуі керек. А. мРНҚ түзілу кезеңдерін және рибосомалардың қосылуын көруге болатын, бактериялық ДНҚ аймағының электронды микрофотосуреті. Б. А микрофотосуретіндегі көрсетілген құрылымның сызба түріндегі суреті.
Жаңа аминқышқыл өсіп жатқан полипептидтік тізбекке қосылғаннан соң бірден, рибосомаға иРНҚ-мен бір кодонға орын ауыстырады. Бұған дейін полипептидтік тізбекпен байланысқан тРНҚ молекуласы рибосоманы қалдырып кетеді және аминоацил-тРНҚ жаңа кешенін қалыптастыру үшін цитоплазмаға қайтып келеді (23.29.В. сур.).
Мұндай мРНҚ кодына орныққан рибосомамен ретті түрде «санау» және «трансляция», процесс стоп-кодондардың біріне жеткенге дейін созылады. Мұндай кодондар ретінде УАА, УАГ және УГА үштіктері қызмет атқарады. Бұл кезеңде, ДНҚ анықталған бірінші реттік құрылым-полипептидтік тізбек, рибосоманы қалдырып кетеді және трансляция аяқталады.
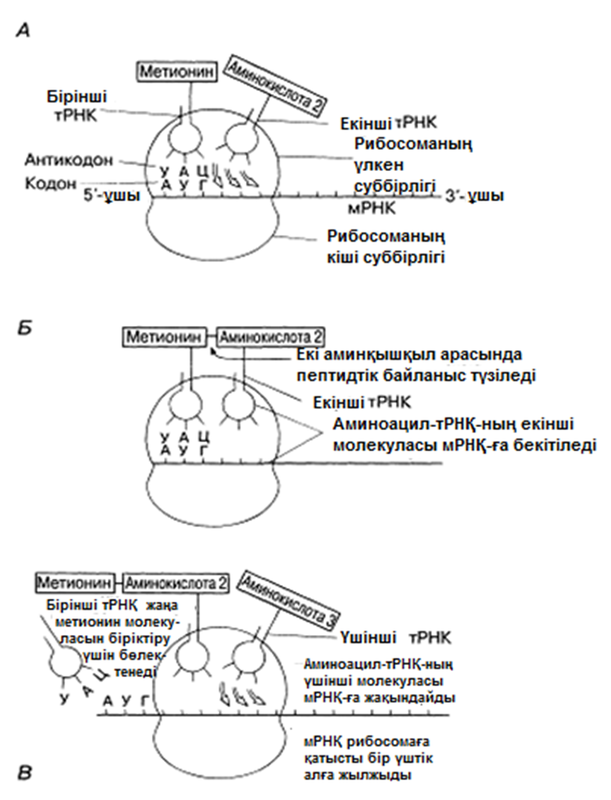
23.29. А және Б суреті . Аминоацил-тРНҚ молекуласының, өздерінің антикодондарымен бірге мРНҚ кодондарына бекітілу процесінің кезеңдері және көршілес аминқышқылдар арасында пептидтік байланыстың түзілуі. В. мРНҚ және рибосоманың , нәтижесінде аминоацил-тРНҚ молекулаларын байланыстыру үшін жаңа үштік ашылатын (жақтауша) қосылуы. Бірінші т-РНҚ молекуласы рибосомадан бөлектенеді және аминоацил-тРНҚ жаңа комплексі түзілуін мүмкін ете отырып, оны ферменттер реактивтейтін, цитоплазмаға қайта келеді.
Трансляция процесінің басты кезеңдері:
1) мРНҚ-ның рибосомаға қосылуы (бірігуі);
2) аминқышқылдың белсенденуі және оның тРНҚ-ға қосылуы (бірігуі);
3) полипептидтік тізбектің иницияциясы (синтездің басталуы);
4) тізбек элонгациясы (ұзаруы);
5) тізбек терминациясы (синтездің аяқталуы);
6) мРНҚ-ның одан әрі қолданылуы немесе оның бұзылуы.
Трансляция процесі сызба түрінде 23.30. суретінде көрсетілген.
Полипептидтік тізбектер рибосомалардан ажыратылғаннан кейін, олар сол бойда өздеріне тән екінші реттік, үшінші реттік немесе төртінші реттік құрылымға ие болады (3.5.3. бөлім). Егер рибосома эндоплазмалық ретикулумға(ЭР) бекітілген болса, онда нәруыз, белгіленген жерге тасымалданатын ЭР-ға түседі. Өсіп жатқан полипептидтік тізбектің бірінші кезеңі, рибосоманың ЭР-ға бірігуін қамтамасыз ететін ЭР мембранасындағы белгілі рецепторға сәйкес келетін аминқышқылдардың «сигналды реттілігінен» тұрады. Өсіп жатқан нәруыздық молекула рецептор арқылы ЭР-ға түседі(23.31.сур.). Нәруыз ЭР-ның ішінде болғаннан кейін, сигналды реттілік одан ажырайды және нәруыздық молекула өзінің соңғы пішіне ие бола отырып қысқарады.
Аминқышқылдың полипептидтік тізбекке қосылуы, аминқышқылдың өзімен емес, мРНҚ кодоны және тРНҚ антикодоны арасындағы негіздердің комплементарлы жұптасуымен анықталатындының дәлелдемесі, келесі эксперименттен алынған болатын. тРНҚ –цистеин молекуласы, әдетте, мРНҚ кодоны УГУ-мен бірге , өзінің антикодоны АЦА көмегімен жұптасады. Бұл кешенге Реней никелі-катализаторы әсер еткен кезде, цистеин аланин аминқышқылына айналады. Мұндай тРНҚ –аланин жаңа кешенін (тРНҚ-цистеиннің антикодонын тасымалдайтын), құрамында жатрылай(УГУ)-мРНҚ бар жасушасыз жүйеге орналастырғанда, синтезделген полипептидтік тізбек тек қана аланиннен тұрды. Бұл эксперимент, генетикалық код трансляциясындағы мРНҚ-кодон — тРНҚ-антикодон кешенінің түзілуі механизмінің маңызды рөлін тұжырымдады.
Нәруыз синтезі процесі түзілетін барлық оқиғалар реттілігі, сызба түрінде 23.32. суретінде ұсынылған.
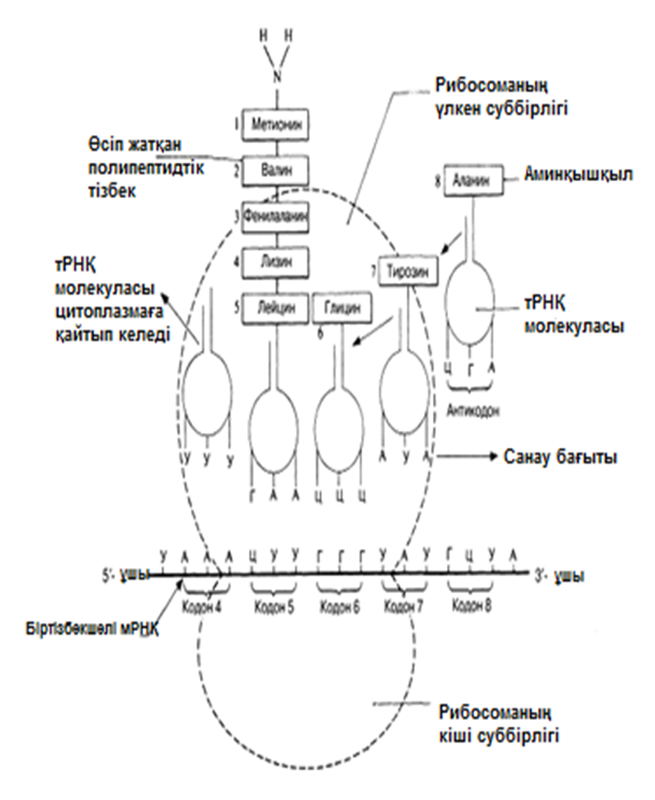
23.30.сур. Трансляция процесінің сызба түріндегі суреті.Әр жеке молекулалардың антикодоны аминоацил-тРНҚ, оған комплементарлы, рибосомадағы мРНҚ кодонымен жұптасады. Бұл жерде суреттелген жағдайда, оның артынан лейцин мен глицин арасындағы пептидтік байланыс түзілуі жүреді және осылайша өсіп жатқан полипептидтік тізбекке тағы бір аминқышқыл қосылады.
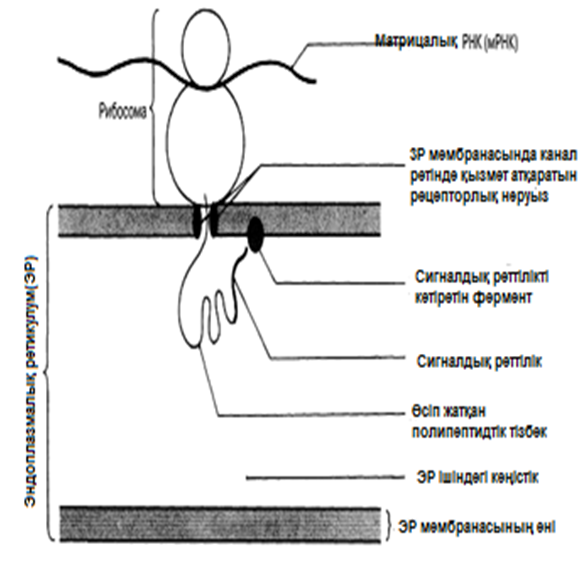
23.31.сур. Жаңадан түзілген нәруыздың, эндоплазмалық ретикулумға түсуі.

23.32.сур.Нәруыздық синтезге қатысатын негізгі құрылымдар мен процесстердің оңайлатылған сызбасы.
23.8.8. Код арқылы реттемейтін ДНҚ
Адам ДНҚ-сы құрамында, шамамен, 3000 млн жұп негіз және, бағалау бойынша, 100000 ген бар, бірақ қазіргі кезе бұл бағалаулар тек қана, шамамен болуы мүмкін. Мәселе, ДНҚ-ның көп бөлігі(шамамен 95%), код арқылы реттеуге қатыспайтындықтан, көргеніміз бойынша, қандай да бір айқын функциялардан айырылғандығында. Басқаша айтқанда, ол нәруыз синтезін де, РНҚ синтезін де код арқылы реттемейді. Кейде оны «жарамсыз ДНҚ» деп атайды, бірақ функцияның жоқтығы тек қана бұл функциялардың белгісіздігінде екенін болжау дұрыс емес. Мүмкін, бұл ДНҚ бөлігі, қазіргі уақытта ешқандай пайдалы мақсатта қызмет атқармайтын бұрынғы гендерді құрайды. Басқа бөлігі, мысалға, хромосомаларды қаптауға қатысу арқылы, құрылымдық функцияларды орындай алады. Бұл ДНҚ-ның, шамамен, 30-40%-ы бірнеше рет қайталанатын қысқа негіздер реттілігінен тұрады. ДНҚ-ның бұл бөлігіне, «генетикалық дактилоскопиядағы» рөлі 25.7.12. бөлімінде сипатталған, сателлитті ДНҚ жатады. ДНҚ-ның, интрон деп аталатын кейбір аймақтары, төменде қарастырылады.
Интрондар және экзондар
1977 жылы биологтар, эукариоттық ағзалардың ДНҚ-сынан, сәйкес мРНҚ-ға қарағанда ұзындау гендерді анықтау арқылы таңғалды. мРНҚ, геннің көшірмесі болғандықтан, генмен дәл бірдей болуы тиіс еді. мРНҚ синтезі аяқталғаннан кейін,сол бойда, оның молекуласының кейбір аймақтары ол трансляцияда қолданылғанға дейін қиылатындығы анықталды.
23.8.8. Код арқылы реттемейтін ДНҚ
Адам ДНҚ-сы құрамында, шамамен, 3000 млн жұп негіз және, бағалау бойынша, 100000 ген бар, бірақ қазіргі кезе бұл бағалаулар тек қана, шамамен болуы мүмкін. Мәселе, ДНҚ-ның көп бөлігі(шамамен 95%), код арқылы реттеуге қатыспайтындықтан, көргеніміз бойынша, қандай да бір айқын функциялардан айырылғандығында. Басқаша айтқанда, ол нәруыз синтезін де, РНҚ синтезін де код арқылы реттемейді. Кейде оны «жарамсыз ДНҚ» деп атайды, бірақ функцияның жоқтығы тек қана бұл функциялардың белгісіздігінде екенін болжау дұрыс емес. Мүмкін, бұл ДНҚ бөлігі, қазіргі уақытта ешқандай пайдалы мақсатта қызмет атқармайтын бұрынғы гендерді құрайды. Басқа бөлігі, мысалға, хромосомаларды қаптауға қатысу арқылы, құрылымдық функцияларды орындай алады. Бұл ДНҚ-ның, шамамен, 30-40%-ы бірнеше рет қайталанатын қысқа негіздер реттілігінен тұрады. ДНҚ-ның бұл бөлігіне, «генетикалық дактилоскопиядағы» рөлі 25.7.12. бөлімінде сипатталған, сателлитті ДНҚ жатады. ДНҚ-ның, интрон деп аталатын кейбір аймақтары, төменде қарастырылады.
Интрондар және экзондар
1977 жылы биологтар, эукариоттық ағзалардың ДНҚ-сынан, сәйкес мРНҚ-ға қарағанда ұзындау гендерді анықтау арқылы таңғалды. мРНҚ, геннің көшірмесі болғандықтан, генмен дәл бірдей болуы тиіс еді. мРНҚ синтезі аяқталғаннан кейін,сол бойда, оның молекуласының кейбір аймақтары ол трансляцияда қолданылғанға дейін қиылатындығы анықталды.
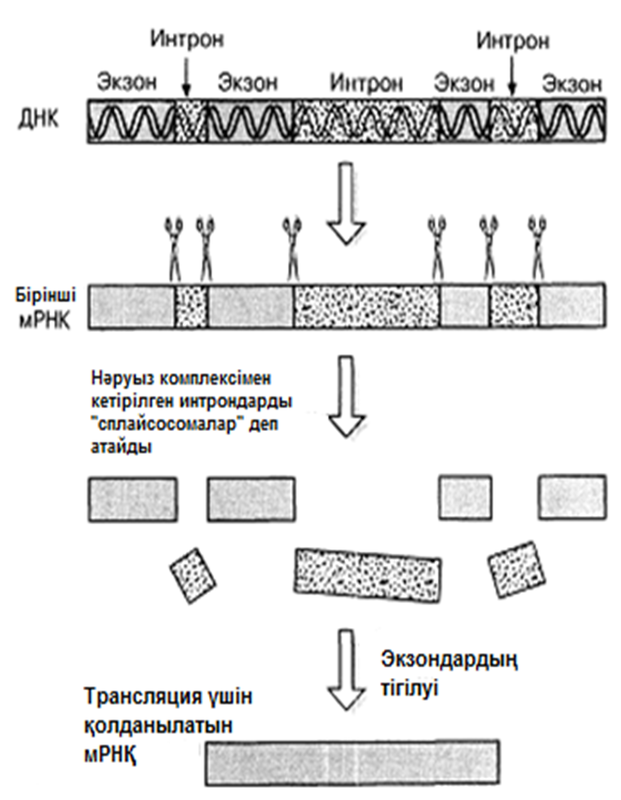
23.33.сур. Экзондар, интрондар және интрондардың кесілуі.
Бұл қолданылмайтын РНҚ аймақтарын код арқылы реттейтін ген аймақтарын, интрондар деп атаған. Сәйкес нәруыз ды код арқылы реттейтін геннің қалған аймақтарын, экзондар деп атаған (23.33. сур.). Интрондардың өлшемі және орналасуы өте алуан түрлі және берілген генге тән. Прокариоттарда интрондар болмайды.
Интрондардың мүмкін функцияларының бірі, әр түрлі жасушалардағы бңрдей мРНҚ-дан әртүрлі интрондар кетірілуі мүмкін болып шыққан кезде анықталды. Осылайша, ген құрамында альтернативті интрон болуы және әртүрлі, бірақ ұқсас нәруыздарда да код арқылы реттеуі мүмкін. Бұл оның потенциалды қолданылуын арттырады. Мысал ретінде кальцитонин гені қызмет атқарады. Бұл ген, қандай интрон кетірілгеніне байланысты, мРНҚ-ның екі түрлі формасын шығара алады. Олардың біреу іқалақанша безде түзіледі және 32 аминқышқылдан тұратын кальцитонин нәруызы синтезін код арқылы реттейді. Кальцитонин— бұл қандағы кальций деңгейін төмендететін гормон. мРНҚ-ның басқа формасы гипоталамуста синтезделеді және 37 аминқышқылдан тұратын нәруызды код арқылы реттейді; бұл пептид деп аталған, кальцитонин генімен байланысқан нәруыз, кальцитонинге ұқсас және тамырды кеңейтетін күшті әсерге ие. Ол , сонымен қатар, перифериялық жүйке жүйесінің кейбір бөліктеріндегі жүйке ұштарынан да бөлінеді.
23.9. Гендік белсенділіктің реттелуі
Генетикалық кодтың мағынасы ашылғаннан және ДНҚ құрылымы орнатылғаннан кейін, генетика саласындағы зерттеулер алға қарай дами түсті. Әсіресе молекулалық генетиктерді ойландырған басты мәселелердің бірі, бұл гендер белсенділігінің реттелуі қандай тәсілмен әр жасушамен бірдей даму программасының және оның өзіндік функцияларының орындалуын қамтамасыз ететіні туралы сұрақ.
Берілген ағзаның барлық соматикалық жасушаларында бірдей гендер болады, яғни құрамында бірдей аллельдерді таситын бірдей хромосома саны бар және қанша дегенмен көпжасушалы ағзалар жасушасы, құрылымы және функциялары бойынша алуан түрлі. Тіпті бірдей жасушаларда нәруыздың молекулалар синтезінің жылдамдығы, жағдайларға және қажеттілікке байланысты әр-түрлі болуы мүмкін. Жасушадағы гендер белсенділігін реттейтін механизмдер туралы ақпараттар, алғаш рет, E.coli-дегі ферменттер синтезі реттелуін зерттеу кезінде алынды.
1961 жылы Жакоб және Моно, E.coli-дегі ферменттер синтезінің индукция табиғатын түсінуді қалап, эксперименттер қатарын жүргізді. Бұл бактерия жасушасында, шамамен, 800 фермент синтезделетіндігін болжайды. Олардың кейбірінің синтезі үздіксіз жүреді және оларды конститутивтік ферменттер деп атайды; басқалары берілген ферменттің субстраты болмауы да мүмкін тиісті индуктор бар болғанда ғана түзіледі. Мұндай мысал ретінде ᵦ -гаметозидаза қызмет атқаратын ферменттерді индуцибельдік ферменттер деп атайды.
Е. coli , құрамында глюкоза бар мәдени ортада тез өседі. Жасушаларды глюкоза орнына лактозасы бар ортаға ауыстырғанда, өсу бірден емес, қысқа кідірістен соң басталады, бірақ содан соң глюкозалы ортадағы сияқты жылдамдықпен жалғасуда. Жүргізілген зерттеулер, лактоза ортасында өсуге, әдетте Е. coli синтездемейтін екі зат болуы қажет екендігін көрсетті: лактозаны глюкоза және галактозаға дейін гидролиздейтін ᵦ -галактозидазалар, және жасушаны ортадан лактозаны тез жұтып алуға қабілетті қылатын лактозопермеазалар. Бұл, орта жағдайларындағы өзгерістер (глюкозаның лактозамен ауысуы) анықталған ферменттің синтезін тудыратындығына мысал ретінде қызмет атқарады. Е. coli –мен жасалған басқа эксперименттер, ортадағы триптофан аминқышқылының жоғары құрамы, триптофансинтазаның— триптофан синтезіне қажет ферменттің шығарылуын басатындығын көрсетті. ᵦ -галактозидаза синтезі фермент индукциясының, ал триптофансинтаза синтезі фермент репрессиясының мысалы ретінде қызмет атқарады. Бұл бақылаулар негізінде Жакоб және Моно индукцияны және репрессияны түсіндіретін механизм — гендерді «қосу және «өшіру» механизмдерін ұсынды.
23.9.1. Жакоб және Моно болжамдары
Нәруыздардан жоғарыда аталған аминқышқыл реттілігін анықтайтын генетикалық нұсқаулар құрылымдық гендерде орныққан. ᵦ -галактозидаза және лактозопермеаза үшін нұсқаулықтар бір хромосомада тығыз бекітілген. Бұл гендердің белсенділігі тағы да бір генмен, болжам бойынша, құрылымдық гендердің белсенді жағдайға ауысуына кедергі келтіретін реттеуші-генмен реттеледі. Реттеуші-ген құрылымдық гендерден біршама қашықтықта орналаса алады. Оның бар екендігінің дәлелдемесі, осы геннен айырылған және сондықтан да, ᵦ -галактозидазаны үздіксіз шығаратын мутанттық жасушалар E.coli –ді зерттеу кезінде алынған.
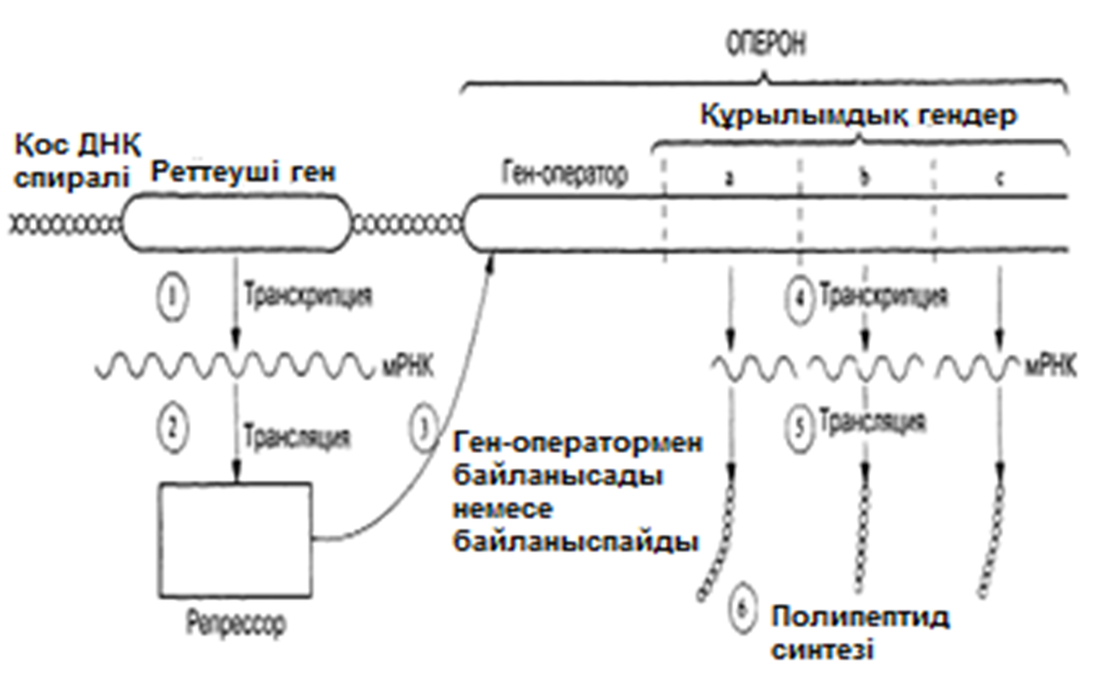
23.34.сур. Жакоб және Моно болжамдарына сәйкес, нәуыз синтезі регуляциясында қатысатын негізгі құрылымдар және процесстер. Сандармен оқиғалардың реттілігі көрсетілген.
Реттеуші ген құрамында құрылымдық гендердің белсенділігіне кедергі келтіретін репрессор синтезі үшін генетикалық ақпарат бар. Репрессор құрылымдың гендерге тікелей емес, құрылымдық гендерге кіретін аймаққа ықпал ете отырып жанама әсер етеді және ол ген-оператор деп аталады. Оператор және құрылымдық гендерді жиынтықта оперон деп атайды(23.34 сур.).
Репрессор, ген-оператормен оның белсенділігін баса отырып байланысатын (оны «өшіреді»), не ген-оператормен оған байланыспайтын (оны «қосады») маңызды аллостериялық нәруызды құрайды. Оператор «өшірулі» болған кезде, мРНҚ түзілмейді және онымен код арқылы реттелетін полипептидтер синтезделмейді (24.34.сур.).
Аллостериялық нәруыздың ген-операторға қосылуы немесе қосылмауы анықталатын механизм қарапайым, бірақ сонымен қатар жасуша ішіндегі өзгерістерге сезімтал. Репрессор молекуласында, олардың, 23.9.4. бөлімінде сипатталғандағыдан қазіргі уақыттағы концентрациясына байланысты, индуктор молекуласы, не репрессор молекуласы қосылуы мүмкін кем дегенде екі белсенді аймағы болатындығы болжанады.
23.9.2. Ферменттер индукциясы
Индуктор молекуласының, оның репрессор молекуласындағы белсенді аймағына қосылуы, үшінші реттік репрессор құрылымын өзертеді, (аллостериялық әсер; 4.4.4. бөлімін қараңыз), сондықтан да ол ген-оператормен байланыса алмайды және оны репрессиялай алмайды. Ген –оператор белсенді болады және құрылымдық гендерді «қосады».
Е. coli –ді глюкозасы бар ортада дамыту кезінде, реттеуші-ген, ген-оператормен байланысатын және оны «өшіретін» репрессор құрамына ие нәруызды шығарады. Сонымен қатар, құрылымдық гендер белсендендірілмейді және ᵦ -галактозидаза, не лактопермаза синтезделмейді. Бактерияларды лактозасы бар ортаға ауыстырған кезде, соңғысы, болжам бойынша, репрессор молекуласына біріге және оның ген-оператормен байланысуына кедергі келтіре отырып, нәруыз синтезінің индукторы ретінде әсер етеді; мРНҚ шығарылады және нәруыздар синтезделеді. Осылайша лактоза өзіндік ұсақтауды тудырады.
23.9.3. Ферменттер репрессиясы
Егер қандай-да бір корепрессордың молекуласы, репрессор молекуласымен бірге өзінің белсенді аймағымен байланысса, онда ол репрессордың ген-опратормен байланысынан пайда болатын әсерді күшейтеді. Сонымен қатар құрылымдық гендердің «қосылуына» нақты кедергі келтіретін, ген-операторының ішкі белбенденуі жүреді.
Е. coli триптофан аминқышқылын, триптофансинтаза ферменті қатысуымен синтездейді. Жасуша құрамында артық триптофан болған кезде, оның кейбір бөліктері репрессор молекуласымен байланыса отырып, фермент синтезінің корепрессоры ретінде әсер етеді. Корепрессор және репрессор молекулалары ген-операторға бірігеді және оның белсенділігін басады. Құрылымдық гендер «өшіріледі», м РНҚ түзілмейді және триптофансинтаза синтезі тоқтатылады. Бұл, гендік жеңгейдегі кері байланыс тәсілімен ингибирлеу мысалы.
23.9.4. Метаболиялық жолдардың реттелуі
Жоғарыда сипатталған индукцияның және репрессияның қос механизмі жасуша метаболизмінің жақсы реттелуін қамтамасыз ету үшін цитоплазма және ядро арасындағы біріге әрекет етуді мүмкін қылады. Қандай-да бір қарапайым метаболиялық жолдар болған жағдайда, бастапқы субстрат және соңғы өнім, сәйкесінше , индуктор және репрессор рөлін ойнай алады. Осының әсерінен жасуша, қазіргі уақытта соңғы өнім мөлшерін төменгі деңгейде ұстап тұру үшін қажетті мөлшерде фермент синтездей алады . Мұндай метаболизм регуляциясының тәсілі өте үнемді. Бірінші ферменттің соңғы өніммен байланысы нәтижесінде, бірінші ферментті ішкі белсендендіру жолымен жүзеге асыратын теріс кері байланыс, берілген метаболиялық жолды тез жауып тастайды, бірақ басқа ферменттредің синтезіне кедергі келтірмейді. Жакоб және Мономен ұсынылған модельде, соңғы өнім, репрессор молекуласына оның ген-операторға ингибирлік әсрін күшейту үшін біріге отырып, барлық, ферментер синтезін басады және берілген метаболиялық жолды өшіреді.
23.9.5. Оперон болжамының модификациясы
1961 жылы Жакоб және Моно гендердің қосылуы және өшірілуі жүзеге асатын механизм ұсынғаннан кейін, бұл механизмнің алуан-түрлі аспектін анықтауға мүмкіндік беретін басқа анықтамалар алынды. Генетикалық анықтамалар негізінде, ген-оператордың жанында орналасатын және онымен және реттеуші-ген арасында әрекет ететін ген-промотордың бар екендігі жөнінде болжам айтылды. Ген-проматор екі функциядан тұратындығы болжанады. Біріншіден, ол ДНҚ-полимераза, транскрипция процесінде ДНҚ айналасына орын ауыстыруды бастамастан бұрын бірігетін жер ретінде, қызмет атқарады, яғни синтез кезінде мРНҚ құрылымдың генде болады. Бұл орын ауыстыру , әрине , ген-оператордың белсенді жағдайда тұру-тұрмауына байланысты. Екіншіден, промотор гендері негіздерінің реттілігі, ДНҚ-ның қос спиралі тізбегінің қайсысы өзіне РНҚ-полимеразаны қосып алатындығын анықтайды, яғни мРНҚ транскрипциясы үшін матрица ретінде қызмет атқарады.
24
ӨЗГЕРГІШТІК ЖӘНЕ ГЕНЕТИКА
Генетиканың биологияның ең маңызды бөлігінің бірі болып саналуына құқығы бар. Мыңдаған жылдар бойы адамдар генетикалық тәсілдерді, бұл тәсілдер негізінде жататын құрылғылар жайында білмей, үй жануарларын және өсірілетін өсімдіктерді жақсарту үшін қолданды. Алуан түрлі археологиялық мәліметтерге қарағанда, 6000 жыл бұрын адамдар кейбір физикалық белгілер бір ұрпақтан екінші ұрпаққа берілетіндігін түсінген. Табиғи популяциядан белгілі ағзаларды іріктеп және оларды өзара шағылыстырып адам, оған қажетті құрылымдарға ие жақсартылған өсімдіктердің сортын және жануарлардың тұқымдарын жасады.
Алайда ХХ ғасырдың басында ғана ғалымдар толықтай түрде тұқымқуалаушылықтың заңдарының және оның құрылғыларының маңыздылығын түсінді. Микроскопия сәттіліктері тұқымқуалайтын белгілердің ұрпақтан ұрпаққа спермий және жұмыртқа жасуша арқылы берілетіндігін анықтауға мүмкіндік берсе де, биологиялық материялдың өте кішкентай бөлшектері өзінде бөлек ағза құралатын өте көп белгілерді қалай алып жүретіндігі белгісіз болып қалды.
Тұқымқуалаушылық жайындағы зерттеулерге бастаманы австралиялық монах, 1866 жылы заманауи генетиканың негізін қалаған мақаланы жариялаған, Грегор Менднль қойды. Менднль тұқымқуалайтын белгілердің араласпайтынын, ата-анадан ұрпақтарына дискретті (жекешеленген) бірліктер түрінде берілетіндігін көрсетті. Ұрпақтарда жұп болып көрсетілген бұл бірліктер, дискретті болып қалады және келесі ұрпаққа әрқайсысының құрамында әр жұптан бір бірліктен болатын аталық және аналық гаметалардан беріледі. 1909 жылы дат ботанигі Иогансен бұл бірліктерді гендер деп атады, ал 1912 жылы американдық генетик Морган олар хромосомаларда орналасатындығын көрсетті. Сол кезден бастап генетика, тұқымқуалаушылық табиғатын ағза деңгейінде және ген деңгейінде түсіндіруден үлкен жетістіктерге жетті.
24.1. Мендельдің зерттеулері
Грегор Мендель 1822 жылы туылды. 1843 жылы ол Брюндегі (қазір Брно, Чехия) августицтердің монастыріне түседі, ол бұл жерде діни атақ қабылдайды. Кейінірек ол Венаға кетеді, ол бұл жердегі университетте тарих және математиканы үйреніп екі жыл өткізді, ал 1853 жылы монастырге қайтып оралады. Мендельмен таңдалған пәндер оның кейінгі асбұршақтан белгілердің тұқымқуалауы жөніндегі жұмыстарына сөз жоқ маңызды әсер етті. Венада болғанының өзінде Мендель өсімдіктердегі будандастыру процесіне және атап айтқанда гибридті ұрпақтардың әр түрлі типтеріне және олардың арақатынасына қызығушылық танытты. Бұл мәселелер Мендельдің 1856 жылы жазда бастаған ғылыми зерттеулерінің мән-мағынасы болды.
Мендельмен жетілген жетістіктер, тәжірибеге сәтті нысанды – егістік асбұршағын немесе кәдімгі асбұршақты (Pisum sativum) таңдаумен жартылай байланысты. Мендель асбұршақтың басқа түрлеріне қарағанда бұл түрінің артықшылықтарға ие екендігіне көз жеткізді.
Мендельге дейін де ғалымдар өсімдіктерге осындай тәжірибелер жүргізді, бірақ олардың бір де біреуі мұндай нақты және толық анықтамалар алған жоқ; сонымен қатар, олар өздерінің нәтижелерін тұқымқуалаушылық механизмдері тұрғысынан түсіндіре алған жоқ. Мендельдің жетістікке жетуін қамтамасыз еткен факторларды, кез-келген ғылыми зерттеулерді жүргізуге қажетті шарттар деп мойымдау керек. Бұл шарттар төменде тұжырымдалған:
Мендель жазғандай, «кез-келген тәжірибенің сенімділігі мен пайдалылығы таңдалған материалдың ол қолданылатын мақсаттарда жарамдылығымен анықталады».
Алайда тәжірибелік нысанды таңдау кезінде Мендельдің бір нәрседен жолы болғандығын атап өту керек: ол таңдап алған тұқымқуалаушылық белгілердің ішінде басымдылық, немесе қосбасымдылық (24.7.1 бөлімі), бір жұптан көп генге тәуелділік (24.7.6 бөлімі), гендердің қиылысуы (24.3 бөлімі) сияқты қиынырақ генетикалық ерекшеліктер қатары болған жоқ.
24.1.1. Моногибридті шағылыстыру кезіндегі тұқымқуалаушылық және ажырау заңы
Өзінің алғашқы тәжірибелеріне Мендель екі түрлі сұрыпты, белгілі бір белгісі бойынша, мысалы гүлдердің орналасуы бойынша анық ажыратылатын өсімдіктерді таңдады: гүлдер бүкіл сабақ бойымен бөлінген болуы мүмкін (қуыстық) немесе сабақтың ұшында орналасу мүмкін (төбелік). Бір жұп альтернативті белгілермен ажыратылатын өсімдіктерді, Мендель ұрпақтар қатары ұзақтығында өсірді. Қуыс гүлді өсімдіктерден алынған тұқымдар әрқашан қуыс гүлді өсімдіктер берді, ал төбе гүлді өсімдіктерден алынған тұқымдар – төбе гүлді өсімдіктер берді. Осылайша, Мендель, өзінің таңдаған өсімдіктері тазалықта көбейетіндігіне көз жеткізді. Бұл оларды будандастыру бойынша сынақтар (тәжірибелік шағылыстыру) жүргізуге жарамды етті. Мендельдің тәсілі келесіде: ол бір сұрыптың өсімдіктер қатарынан тозаңдарды, өздігінен тозаңдану болғанға дейін алып тастады. Бұл өсімдіктерді Мендель «аналық» деп атады. Содан соң, қылқалам қолдана отырып, ол бұл «аналық гүлдер» аузына басқа сұрып өсімдіктердің тозаңдығынан тозаңшаны жақты. Содан кейін ол жасанды тозаңданған гүлдерге, олардың ауыздарына басқа өсімдік тозаңдары түсіп кету мүмкіндігін болдырмау үшін кішкентай қалпақшалар кигізді. Мендель ілеспелі шағылыстыруды жүргізді – тозаңды қуыс гүлдерден төбе гүлдерге және төбе гүденден қуыс гүлдерге ауыстырды. Барлық жағдайларда алынған будандардан жиналған тұқымдардан қуыс гүлді өсімдіктер өсіп шықты. Бірінші будандық ұрпақ өсімдіктерінде (кейінірек, 1902 жылы Бэтсон және Сондерс оны F1 белгісімен белгілей бастады) бақыланатын бұл – «қуыс гүлдер» белгісін, Мендель доминантты деп атады. F1 –дің бір де бір өсімдігінде төбе гүлдер болған жоқ.
F1 өсімдігінің гүлдеріне Мендель қалпақшалар кигізді (айқас тозаңдануды болдырмау үшін) және оларға өздігінен тозаңдануға мүмкіндік берді. Бұл F1 өсімдігінен жиналған тұқымдар түгенделді және келесі көктемде екінші будандық ұрпақ, немесе F2 ұрпағын алу үшін егілді. (F2 ұрпағы – бұл әрқашан F1 ұрпағындағы инбридинг нәтижесі немесе, берілген жағдайдағы сияқты өздігінен тозаңдану) Бұл өсімдіктер гүлдеген кезде, олардың біреулерінде қуыс гүлдер түзілді, ал басқаларында – төбе гүлдер. Басқаша айтқанда F1 ұрпағында болмаған төбе гүлдер белгісі F2 ұрпағында қайта пайда болды. Мендель бұл белгінің F1 ұрпағында жасырын түрде болып, бірақ шыға алмаған деген қорытындыға келді, сондықтан да оны рецессивті деп атады. F2 –де Мендельмен алынған 858 өсімдіктің, 651-інде қуыс гүлдер, ал 207-сінде төбе гүлдер болды. Мендель әрдайым альтернативті белгілер жұптарын қолдана отырып, ұқсас сынақтар қатарын жүргізді. Осындай белгілердің жеті жұбы бойынша тәжірибелік шағылыстыру нәтижелері 24.1 кестесінде ұсынылған. Барлық жағдайларда нәтижелер анализі, F2 ұрпағында доминантты белгілердің рецессивті белгілерге қатынасы шамамен 3:1 құрағаныдығын көрсетті.
Жоғарыда ұсынылған мысал, бір ғана белгінің тұқымқуалауы (моногибридті шағылыстыру) зерттелген Мендельдің барлық тәжірибелеріне ортақ. Қысқаша ол мынаған саяды:
Алайда ХХ ғасырдың басында ғана ғалымдар толықтай түрде тұқымқуалаушылықтың заңдарының және оның құрылғыларының маңыздылығын түсінді. Микроскопия сәттіліктері тұқымқуалайтын белгілердің ұрпақтан ұрпаққа спермий және жұмыртқа жасуша арқылы берілетіндігін анықтауға мүмкіндік берсе де, биологиялық материялдың өте кішкентай бөлшектері өзінде бөлек ағза құралатын өте көп белгілерді қалай алып жүретіндігі белгісіз болып қалды.
Тұқымқуалаушылық жайындағы зерттеулерге бастаманы австралиялық монах, 1866 жылы заманауи генетиканың негізін қалаған мақаланы жариялаған, Грегор Менднль қойды. Менднль тұқымқуалайтын белгілердің араласпайтынын, ата-анадан ұрпақтарына дискретті (жекешеленген) бірліктер түрінде берілетіндігін көрсетті. Ұрпақтарда жұп болып көрсетілген бұл бірліктер, дискретті болып қалады және келесі ұрпаққа әрқайсысының құрамында әр жұптан бір бірліктен болатын аталық және аналық гаметалардан беріледі. 1909 жылы дат ботанигі Иогансен бұл бірліктерді гендер деп атады, ал 1912 жылы американдық генетик Морган олар хромосомаларда орналасатындығын көрсетті. Сол кезден бастап генетика, тұқымқуалаушылық табиғатын ағза деңгейінде және ген деңгейінде түсіндіруден үлкен жетістіктерге жетті.
24.1. Мендельдің зерттеулері
Грегор Мендель 1822 жылы туылды. 1843 жылы ол Брюндегі (қазір Брно, Чехия) августицтердің монастыріне түседі, ол бұл жерде діни атақ қабылдайды. Кейінірек ол Венаға кетеді, ол бұл жердегі университетте тарих және математиканы үйреніп екі жыл өткізді, ал 1853 жылы монастырге қайтып оралады. Мендельмен таңдалған пәндер оның кейінгі асбұршақтан белгілердің тұқымқуалауы жөніндегі жұмыстарына сөз жоқ маңызды әсер етті. Венада болғанының өзінде Мендель өсімдіктердегі будандастыру процесіне және атап айтқанда гибридті ұрпақтардың әр түрлі типтеріне және олардың арақатынасына қызығушылық танытты. Бұл мәселелер Мендельдің 1856 жылы жазда бастаған ғылыми зерттеулерінің мән-мағынасы болды.
Мендельмен жетілген жетістіктер, тәжірибеге сәтті нысанды – егістік асбұршағын немесе кәдімгі асбұршақты (Pisum sativum) таңдаумен жартылай байланысты. Мендель асбұршақтың басқа түрлеріне қарағанда бұл түрінің артықшылықтарға ие екендігіне көз жеткізді.
- Оның белгілер қатары бойынша анық ажыратылатын көп сұрпы бар.
- Өсімдікті өсіру оңай.
- Репродуктивті органдар толықтай күлтелермен жабылған, сол себептен өсімдік әдетте өздігінен тозаңданады. Сондықтан барлық сұрыптар тазалықта көбейеді, яғни олардың белгілері ұрпақтан-ұрпаққа өзгеріссіз беріледі.
- Жасанды шағылыстыру арқылы фертильді гибридті ұрпақ алуға болады. Асбұршақтың 34 сұрпынан Мендель белгілер қатары бойынша анық белгіленген айырмашылықтарға ие 22 сұрпын таңдап алды, және оларды өзінің шағылыстыруларында қолданды. Мендельді жеті басты белгі қызықтырды: сабақтың ұзындығы, тұқымның пішіні және түсі, жемістің пішіні және түсі және гүлдердің орналасуы және түсі.
Мендельге дейін де ғалымдар өсімдіктерге осындай тәжірибелер жүргізді, бірақ олардың бір де біреуі мұндай нақты және толық анықтамалар алған жоқ; сонымен қатар, олар өздерінің нәтижелерін тұқымқуалаушылық механизмдері тұрғысынан түсіндіре алған жоқ. Мендельдің жетістікке жетуін қамтамасыз еткен факторларды, кез-келген ғылыми зерттеулерді жүргізуге қажетті шарттар деп мойымдау керек. Бұл шарттар төменде тұжырымдалған:
- Тәжірибелік нысанмен танысу үшін алдын-ала зерттеулер жүргізу.
- Барлық тәжірибелерді бір айнымалыға аудару бақылау жүргізуді жеңілдетеді.
- Нәтижелерді бұрмалайтын айнымалылардың енгізілу мүмкіндігін болғызбау үшін тәжірибе тәртібін қатаң ұстану.
- Барлық тәжірибелерді және алынған нәтижелерді қатаң құжаттау.
- Статистикалық сенімді деп санай алатындай жеткілікті мөлшердегі ақпараттарды алу.
Мендель жазғандай, «кез-келген тәжірибенің сенімділігі мен пайдалылығы таңдалған материалдың ол қолданылатын мақсаттарда жарамдылығымен анықталады».
Алайда тәжірибелік нысанды таңдау кезінде Мендельдің бір нәрседен жолы болғандығын атап өту керек: ол таңдап алған тұқымқуалаушылық белгілердің ішінде басымдылық, немесе қосбасымдылық (24.7.1 бөлімі), бір жұптан көп генге тәуелділік (24.7.6 бөлімі), гендердің қиылысуы (24.3 бөлімі) сияқты қиынырақ генетикалық ерекшеліктер қатары болған жоқ.
24.1.1. Моногибридті шағылыстыру кезіндегі тұқымқуалаушылық және ажырау заңы
Өзінің алғашқы тәжірибелеріне Мендель екі түрлі сұрыпты, белгілі бір белгісі бойынша, мысалы гүлдердің орналасуы бойынша анық ажыратылатын өсімдіктерді таңдады: гүлдер бүкіл сабақ бойымен бөлінген болуы мүмкін (қуыстық) немесе сабақтың ұшында орналасу мүмкін (төбелік). Бір жұп альтернативті белгілермен ажыратылатын өсімдіктерді, Мендель ұрпақтар қатары ұзақтығында өсірді. Қуыс гүлді өсімдіктерден алынған тұқымдар әрқашан қуыс гүлді өсімдіктер берді, ал төбе гүлді өсімдіктерден алынған тұқымдар – төбе гүлді өсімдіктер берді. Осылайша, Мендель, өзінің таңдаған өсімдіктері тазалықта көбейетіндігіне көз жеткізді. Бұл оларды будандастыру бойынша сынақтар (тәжірибелік шағылыстыру) жүргізуге жарамды етті. Мендельдің тәсілі келесіде: ол бір сұрыптың өсімдіктер қатарынан тозаңдарды, өздігінен тозаңдану болғанға дейін алып тастады. Бұл өсімдіктерді Мендель «аналық» деп атады. Содан соң, қылқалам қолдана отырып, ол бұл «аналық гүлдер» аузына басқа сұрып өсімдіктердің тозаңдығынан тозаңшаны жақты. Содан кейін ол жасанды тозаңданған гүлдерге, олардың ауыздарына басқа өсімдік тозаңдары түсіп кету мүмкіндігін болдырмау үшін кішкентай қалпақшалар кигізді. Мендель ілеспелі шағылыстыруды жүргізді – тозаңды қуыс гүлдерден төбе гүлдерге және төбе гүденден қуыс гүлдерге ауыстырды. Барлық жағдайларда алынған будандардан жиналған тұқымдардан қуыс гүлді өсімдіктер өсіп шықты. Бірінші будандық ұрпақ өсімдіктерінде (кейінірек, 1902 жылы Бэтсон және Сондерс оны F1 белгісімен белгілей бастады) бақыланатын бұл – «қуыс гүлдер» белгісін, Мендель доминантты деп атады. F1 –дің бір де бір өсімдігінде төбе гүлдер болған жоқ.
F1 өсімдігінің гүлдеріне Мендель қалпақшалар кигізді (айқас тозаңдануды болдырмау үшін) және оларға өздігінен тозаңдануға мүмкіндік берді. Бұл F1 өсімдігінен жиналған тұқымдар түгенделді және келесі көктемде екінші будандық ұрпақ, немесе F2 ұрпағын алу үшін егілді. (F2 ұрпағы – бұл әрқашан F1 ұрпағындағы инбридинг нәтижесі немесе, берілген жағдайдағы сияқты өздігінен тозаңдану) Бұл өсімдіктер гүлдеген кезде, олардың біреулерінде қуыс гүлдер түзілді, ал басқаларында – төбе гүлдер. Басқаша айтқанда F1 ұрпағында болмаған төбе гүлдер белгісі F2 ұрпағында қайта пайда болды. Мендель бұл белгінің F1 ұрпағында жасырын түрде болып, бірақ шыға алмаған деген қорытындыға келді, сондықтан да оны рецессивті деп атады. F2 –де Мендельмен алынған 858 өсімдіктің, 651-інде қуыс гүлдер, ал 207-сінде төбе гүлдер болды. Мендель әрдайым альтернативті белгілер жұптарын қолдана отырып, ұқсас сынақтар қатарын жүргізді. Осындай белгілердің жеті жұбы бойынша тәжірибелік шағылыстыру нәтижелері 24.1 кестесінде ұсынылған. Барлық жағдайларда нәтижелер анализі, F2 ұрпағында доминантты белгілердің рецессивті белгілерге қатынасы шамамен 3:1 құрағаныдығын көрсетті.
Жоғарыда ұсынылған мысал, бір ғана белгінің тұқымқуалауы (моногибридті шағылыстыру) зерттелген Мендельдің барлық тәжірибелеріне ортақ. Қысқаша ол мынаған саяды:
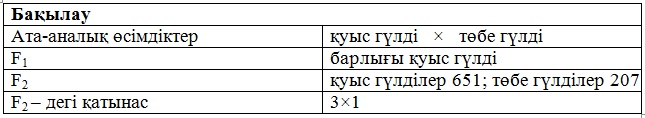
Осы және осыған ұқсас нәтижелерге негізделе, Мендель келесі қорытындыларға келді:
24.1кестесі. Альтернативті белгілердің жеті жұбының тұқымқуалауы бойынша Мендель тәжірибелерінің нәтижелері. (Доминантты және рецессивті белгілердің бақыланатын қатынасы теориялық түрде күтілетін 3:1 қатынасына жақын)
- Қанша дегенмен бастапқы ата-ананық сұрыптар тазалықта көбейгендіктен, қуыс гүлді сұрыпта екі «қуыс факторы» болуы қажет, ал төбе гүлді сұрыпта – екі «төбе факторы» болуы қажет.
- F1 өсімдіктері құрамында әр ата-аналық өсімдіктен гаметалар арқылы алынған бір-бір фактордан болды.
- Бұл факторлар F1-де араласпайды, өзінің жекешелігін сақтайды.
- «Қуыс» фактор, баслыңқы «төбе» фактордан басым болады.
24.1кестесі. Альтернативті белгілердің жеті жұбының тұқымқуалауы бойынша Мендель тәжірибелерінің нәтижелері. (Доминантты және рецессивті белгілердің бақыланатын қатынасы теориялық түрде күтілетін 3:1 қатынасына жақын)
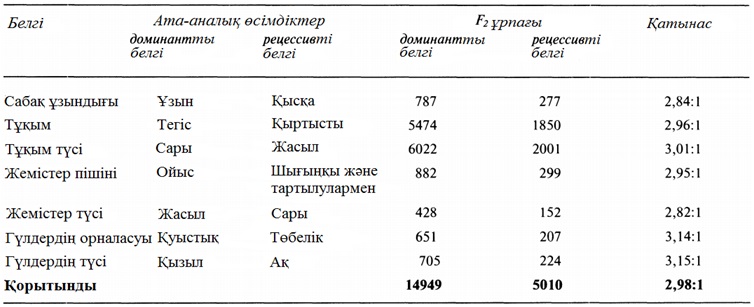
Гаметалар түзілуі кезіндегі ата-аналық факторлар жұбының, нәтижесінде әр гаметада олардың тек біреуі ғана қалатындай ажырауы, Мендельдің бірінші заңы, немесе ажырау заңы атымен белгілі. Бұл заңға сәйкес, берілген ағзаның барлық белгілері қандай да бір ішкі факторлар жұбымен анықталады. Бір гаметада мұндай факторлар жұбының біреуі ғана көрсетілуі мүмкін.
Белгілерді анықтайтын жорамал факторлар, гендер деп аталатын хромосома бөліктерін құрайды (мысалы, гүлдердің орналасуы және түсі, тұқымдардың пішіні және түсі және т.б.).
Жоғарыда сипатталған альтернативті белгілердің бір жұбының тұқымқуылаушылығын зерттеуде Мендельмен жүргізілген тәжірибелер моногибридті шағылысудың мысалы ретінде қызмет атқарады. Оны белгілер көмегімен сипаттауға болады және гаметалардың қалыптасуы және ұрықтану туралы заманауи болжамдармен байланыстыруға болады. Генді доминантты белгіні сипаттайтын сөздің бірінші әрпәмен белгілеу қабыладанды, яғни үлкен әріппен (мысалы, А) доминантты аллельді белгілейді, ал кіші әріппен (мысалы, а) – рецессивті аллельді белгілейді. Жоғарыда қарастырылған барлық терминдер және белгілер 24.2 кестесінде түсіндірілген.
24.1 суретінде моногибридті шағылысуды сипаттаудың дұрыс тәсілі немесе альтернативті белгілердің бір жұбының тұқымқуалауы бойынша генетикалық есептерді шешу көрсетілген.
24.2 кестесі. Көбірек қолданылатын генетикалық терминдер (24.1 суретінде ұсынылған шағылысулар мысалы бойынша түсіндірулерімен)
Белгілерді анықтайтын жорамал факторлар, гендер деп аталатын хромосома бөліктерін құрайды (мысалы, гүлдердің орналасуы және түсі, тұқымдардың пішіні және түсі және т.б.).
Жоғарыда сипатталған альтернативті белгілердің бір жұбының тұқымқуылаушылығын зерттеуде Мендельмен жүргізілген тәжірибелер моногибридті шағылысудың мысалы ретінде қызмет атқарады. Оны белгілер көмегімен сипаттауға болады және гаметалардың қалыптасуы және ұрықтану туралы заманауи болжамдармен байланыстыруға болады. Генді доминантты белгіні сипаттайтын сөздің бірінші әрпәмен белгілеу қабыладанды, яғни үлкен әріппен (мысалы, А) доминантты аллельді белгілейді, ал кіші әріппен (мысалы, а) – рецессивті аллельді белгілейді. Жоғарыда қарастырылған барлық терминдер және белгілер 24.2 кестесінде түсіндірілген.
24.1 суретінде моногибридті шағылысуды сипаттаудың дұрыс тәсілі немесе альтернативті белгілердің бір жұбының тұқымқуалауы бойынша генетикалық есептерді шешу көрсетілген.
24.2 кестесі. Көбірек қолданылатын генетикалық терминдер (24.1 суретінде ұсынылған шағылысулар мысалы бойынша түсіндірулерімен)

3:1 сияқты доминантты фенотиптердің рецессивтіге қатынасы, моногибридті шағылысудағы F2 ұрпағына тән. Мендельдің әр гамета бойынша бір аллельдің тасымалдануы және оның фенотиптік көрінуі туралы қорытындылары анықталған заңдарға сәйкес келеді. F1 гетерозиготалы ата-аналық дарақтан түзілген гаметаның доминантты аллель А немесе рецессивті аллель а-ны тасымалдау ықтималдылығы 50% немесе ½-ге тең. Егер берілген ата-аналық дарақ гаметаларының арасында, әр екі типтің гаметалары ½ ықтималдылығымен кездесетін болса, ұрықтандырудағы гаметалардың төрт мүмкін комбинациялардың әрқайсысының ықтималдылығы ½ × ½ = ¼ құрайды. Сондықтан да F2- нің 4 генотипі мүмкін. Кездейсоқ ұрықтанудағы құрамында А және а аллельдері болатын гаметалардың кездесуінің статистикалық ықтималдығы 24.2 суретінде көрсетілген. Доминирлеу салдарынан ұрпақтар арасынадағы фенотиптер қатынасы 1 рецессивті фенотипке 3 доминантты фенотипті құрайды. 24.1 кестесінде көрсетілгендей, Мендельмен оның шағылыстыру жөніндегі тәжірибелерінде алынған нәтижелер, бұл теориялық түрде шығарылған қатынасты дәлелдейді.
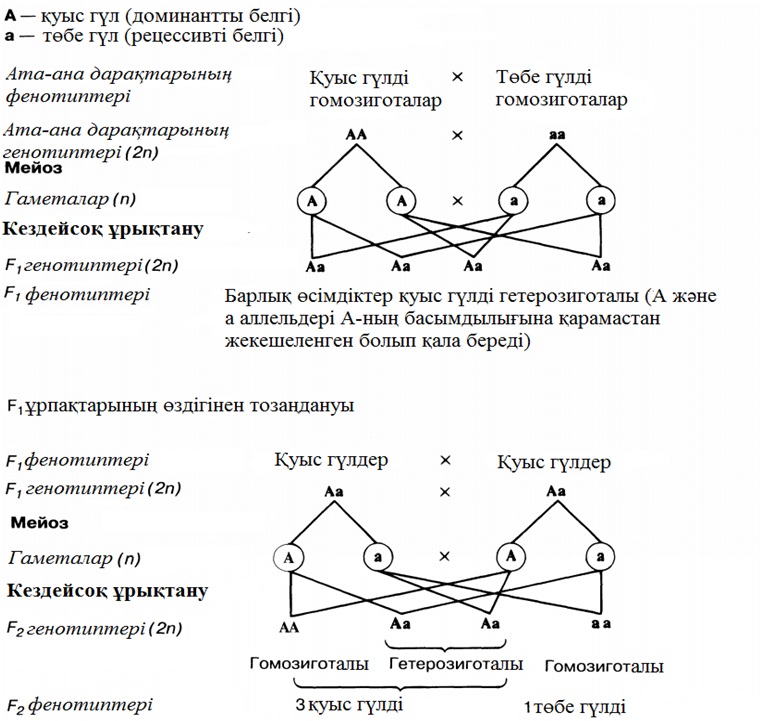
24.1 суреті. Мендельмен жүргізілген моногибридті шағылыстырулардың бірінің толық генетикалық түсіндірмесі.(2n-диплоидтық жағдай, n-гаплоидтық жағдай;23.1.2 бөлімі)

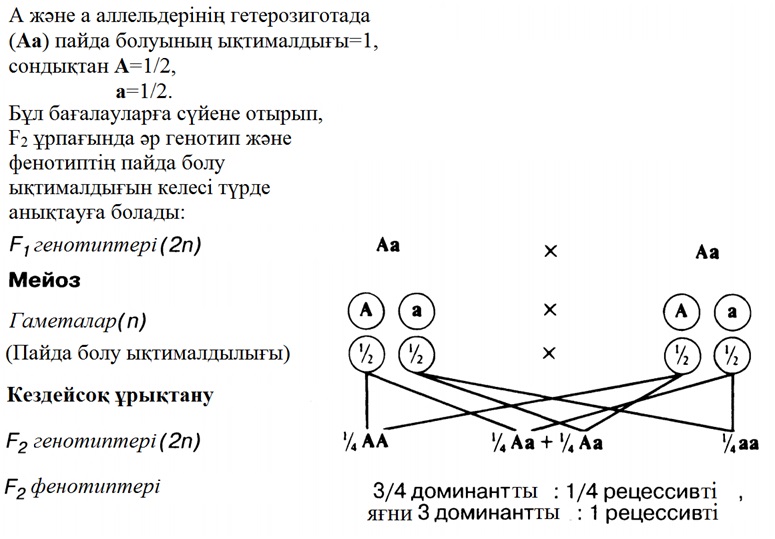
24.2 суреті. Моногибридті шағылысудағы мендельдік қатынас 3:1-дің ықтимал түсіндірмесі.
24.1.2. Анализдейтін шағылыстыру
Гомозиготалы доминантты және гомозиготалы рецессивті дарақтар арасында шағылыстырудан алынған F1 ұрпағының ағзасы гетерозиготалы, бірақ доминантты фенотипке ие. Рецессивті фенотип көрінуі үшін, ағза рецессивті ген арқылы гомозиготалы болуы керек. F2 ұрпағында доминантты фенотипті дарақтар гомозиготалы да, гетерозиготалы да бола алады. Егер селекционерге мұндай дарақтың генотипін анықтау керекболса, онда мұны істеудің бір ғана жолы – анализдейтін шағылыстыру жүргізу. Генотипі белгісіз ағзаны, рецессивті ген арқылы гомозиготалы ағзамен шағылыстыра отырып, бұл генотипті шағылыстырумен бірге анықтауға болады. Мысалы, топырақ шыбыны Drosophila-да ұзын қанаттары өскіндіге басымдылық көрсетеді. Ұзын қанатты дарақ гомозиготалы (LL) немесе гетерозиготалы (LI) бола алады. Оның генотипін анықтау үшін бұл шыбын мен рецессивті аллель (ІІ) бойынша гомозиготалы шыбын арасында анализдейтін шағылыстыру жүргізу қажет. Бұл шағылыстырудан барлық ұрпақта ұзын қанат болса, демек, белгісіз генотипті дарақдоминантты аллель бойынша гомозиготалы болады. Ұзын және өскін қанатты ұрпақтардың сандық қатынасы 1:1, белгісіз генотипті дарақтың гетерозиготалы екендігін көрсетеді (24.3 суреті).
Гомозиготалы доминантты және гомозиготалы рецессивті дарақтар арасында шағылыстырудан алынған F1 ұрпағының ағзасы гетерозиготалы, бірақ доминантты фенотипке ие. Рецессивті фенотип көрінуі үшін, ағза рецессивті ген арқылы гомозиготалы болуы керек. F2 ұрпағында доминантты фенотипті дарақтар гомозиготалы да, гетерозиготалы да бола алады. Егер селекционерге мұндай дарақтың генотипін анықтау керекболса, онда мұны істеудің бір ғана жолы – анализдейтін шағылыстыру жүргізу. Генотипі белгісіз ағзаны, рецессивті ген арқылы гомозиготалы ағзамен шағылыстыра отырып, бұл генотипті шағылыстырумен бірге анықтауға болады. Мысалы, топырақ шыбыны Drosophila-да ұзын қанаттары өскіндіге басымдылық көрсетеді. Ұзын қанатты дарақ гомозиготалы (LL) немесе гетерозиготалы (LI) бола алады. Оның генотипін анықтау үшін бұл шыбын мен рецессивті аллель (ІІ) бойынша гомозиготалы шыбын арасында анализдейтін шағылыстыру жүргізу қажет. Бұл шағылыстырудан барлық ұрпақта ұзын қанат болса, демек, белгісіз генотипті дарақдоминантты аллель бойынша гомозиготалы болады. Ұзын және өскін қанатты ұрпақтардың сандық қатынасы 1:1, белгісіз генотипті дарақтың гетерозиготалы екендігін көрсетеді (24.3 суреті).
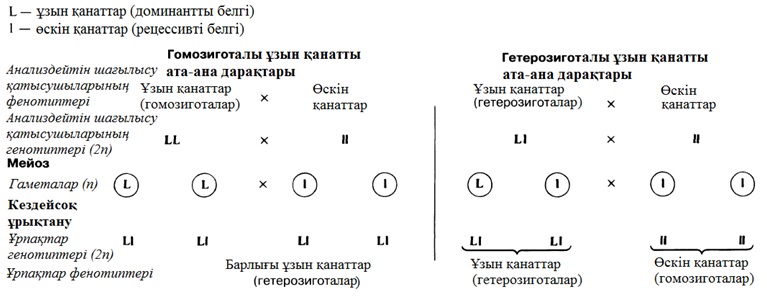
24.3 суреті. Қандай да бір белгісі бойынша доминантты болып табылатын ағзаның генотипін анықтауға мүмкіндік беретін тәсілдің толық генетикалық түсіндірмесі. Бұл тәсіл анализдейтін шағылысу атымен белгілі. Бұдан алынатын ұрпақтардың фенотиптері көрсетілген.

24.1.3. Дигибридті шағылыстырудағы тұқымқуалаушылық және тәуелсіз ажырау заңы
Альтернативті белгілердің бір жұбы бойынша шағылыстыру нәтижелерін болжау мүмкіндігін орната отырып, Мендель мұндай белгілердің екі жұбының тұқымқуалауын зерттеуге көшті. Екі белгі бойынша ажыратылатын дарақтар арасындағы шағылыстыру, дигибридті деп аталады.
Өзінің тәжірибелерінің бірінде Мендель пішіні және тұқымының түсі бойынша ажыратылатын асбұршақ өсімдігін қолданды. 24.1.1 бөлімінде сипатталған тәсілді қолдана отырып, ол тегіс сары тұқымды таза сұрыпты (гомозиготалы) өсімдікті және қыртысты жасыл тұқымды таза сұрыпты өсімдікті өзара шағыластырды.Барлық F1 өсімдіктерде тұқымдары тегіс және сары болды. Бұрын жүргізілген моногибридті шағылыстыру нәтижелері бойынша Мендель бұл белгілердің доминантты екендігін білді; енді, алайда, F1 өсімдігінен өздігінен тозаңдану нәтижесінде алынған F2 ұрпағындағы әр типтің тұқымдарының сипаты және қатынасы қызықтырды. F2 өсімдігінен ол барлығы 556 тұқым жинады, олардың ішінде:
Тегіс сары 315
Қыртысты сары 101
Тегіс жасыл 108
Қыртысты жасыл 32
Әртүрлі фенотиптер қатынасы шамамен 9:3:3:1 қатынасын құрады (дигибридті ажырау). Алынған нәтижелер негізінде Мендель екі қорытынды жасады.
Бұл нәтижелер Мендельге, тұқымқуалайтын қабілеттері F1 ұрпағына біріккен белгілердің екі жұбының (пішіні және тұқымның түсі), келесі ұрпақтарда ажырайтындығын және өздерін бір-біріне тәуелсіз ұстайтындығын дәлелдеуге мүмкіндік берді Мендельдің екінші заңы – тәуелсіз ажырау тәсілі осында негізделген, оған сәйкес:
белгілердің бір жұбының әр белгісі, басқа жұптың кез-келген белгісімен үйлесе алады.
Жоғарыда ұсынылған тәжірибені біз белгілі генетикалық белгілер көмегімен 24.4.А суретінде көрсетілгендей сипаттауға болады. Аллельдердің бөлінуінің (сегрегация) (R, r, Y және y) және олардың тәуелсіз ажырауының (рекомбинация) нәтижесінде, аталық және аналық гаметалардың әрқайсысында аллельдердің әрқайсысында аллельдердің төрт үйлесімінің біреуі мүмкін. Кездейсоқ ұрықтанудапайда болатын, гаметалардың барлық мүмкін үйлесімділігін көрсету үшін, Петтен торы пішініндегі жазбаны қолданады, ол бұлай кембридж генетигі Р. Пеннеттің атымен аталған. Ол гаметалардың барлық мүмкін үйлесімділігінің тізімін құруда тууы мүмкін қателерді минимумға әкеледі. Пеннет торын толтыру кезінде алдымен барлық «аталық» гаметаларды вертикальды баған торшаларына, ал содан соң барлық «аналық» гаметаларды – горизонталь жолдағы торшаларға қою ұсынылады. Сонымен қатар, F2 дарақтарының фенотиптерін анықтай отырып, ұқсас фенотптерді қандай да бір жақсы ажыратылатын таңбалармен белгілеу пайдалы (24.4.Б суретіндегі сияқты). 24.4.А және Б суреттерінде көрсетілгендей, Мендельдің бірінші және екінші заңдарымен сәйкес, F1-дің әр аталық және аналық гаметаларында, аллельдердің келесі үйлесімділігімен гаметалардың түзілуі мүмкін:
R тек қана Y немесе y-мен үйлесімділікте кездесе алады (бірақ r-мен емес), яғни RY немесе Ry түрінде.
r тек қана Y немесе y-мен үйлесімділікте кездесе алады (бірақ R-мен емес), яғни rY немесе ry түрінде.
Осылайша, кез-келген гаметаға осында көрсетілген аллельдер үйлесімділігінің төртеуінің қандай да бір біреуін алу мүмкіндігі 4-тен 1-ге тең.
Альтернативті белгілердің бір жұбы бойынша шағылыстыру нәтижелерін болжау мүмкіндігін орната отырып, Мендель мұндай белгілердің екі жұбының тұқымқуалауын зерттеуге көшті. Екі белгі бойынша ажыратылатын дарақтар арасындағы шағылыстыру, дигибридті деп аталады.
Өзінің тәжірибелерінің бірінде Мендель пішіні және тұқымының түсі бойынша ажыратылатын асбұршақ өсімдігін қолданды. 24.1.1 бөлімінде сипатталған тәсілді қолдана отырып, ол тегіс сары тұқымды таза сұрыпты (гомозиготалы) өсімдікті және қыртысты жасыл тұқымды таза сұрыпты өсімдікті өзара шағыластырды.Барлық F1 өсімдіктерде тұқымдары тегіс және сары болды. Бұрын жүргізілген моногибридті шағылыстыру нәтижелері бойынша Мендель бұл белгілердің доминантты екендігін білді; енді, алайда, F1 өсімдігінен өздігінен тозаңдану нәтижесінде алынған F2 ұрпағындағы әр типтің тұқымдарының сипаты және қатынасы қызықтырды. F2 өсімдігінен ол барлығы 556 тұқым жинады, олардың ішінде:
Тегіс сары 315
Қыртысты сары 101
Тегіс жасыл 108
Қыртысты жасыл 32
Әртүрлі фенотиптер қатынасы шамамен 9:3:3:1 қатынасын құрады (дигибридті ажырау). Алынған нәтижелер негізінде Мендель екі қорытынды жасады.
- F2 ұрпағында екі жаңа белгілердің үйлесімі пайда болды: қыртысты және сары; тегіс және жасыл.
- Аллеломорфты белгілердің (әр-түрлі аллельдермен анықталатын белгілердің) әр жұбының қатынасы 3:1 құрады, бұл моногибридті шағылысуға тән: 133 қыртыстыға 423 тегіс, 140 жасылға 416 сары.
Бұл нәтижелер Мендельге, тұқымқуалайтын қабілеттері F1 ұрпағына біріккен белгілердің екі жұбының (пішіні және тұқымның түсі), келесі ұрпақтарда ажырайтындығын және өздерін бір-біріне тәуелсіз ұстайтындығын дәлелдеуге мүмкіндік берді Мендельдің екінші заңы – тәуелсіз ажырау тәсілі осында негізделген, оған сәйкес:
белгілердің бір жұбының әр белгісі, басқа жұптың кез-келген белгісімен үйлесе алады.
Жоғарыда ұсынылған тәжірибені біз белгілі генетикалық белгілер көмегімен 24.4.А суретінде көрсетілгендей сипаттауға болады. Аллельдердің бөлінуінің (сегрегация) (R, r, Y және y) және олардың тәуелсіз ажырауының (рекомбинация) нәтижесінде, аталық және аналық гаметалардың әрқайсысында аллельдердің әрқайсысында аллельдердің төрт үйлесімінің біреуі мүмкін. Кездейсоқ ұрықтанудапайда болатын, гаметалардың барлық мүмкін үйлесімділігін көрсету үшін, Петтен торы пішініндегі жазбаны қолданады, ол бұлай кембридж генетигі Р. Пеннеттің атымен аталған. Ол гаметалардың барлық мүмкін үйлесімділігінің тізімін құруда тууы мүмкін қателерді минимумға әкеледі. Пеннет торын толтыру кезінде алдымен барлық «аталық» гаметаларды вертикальды баған торшаларына, ал содан соң барлық «аналық» гаметаларды – горизонталь жолдағы торшаларға қою ұсынылады. Сонымен қатар, F2 дарақтарының фенотиптерін анықтай отырып, ұқсас фенотптерді қандай да бір жақсы ажыратылатын таңбалармен белгілеу пайдалы (24.4.Б суретіндегі сияқты). 24.4.А және Б суреттерінде көрсетілгендей, Мендельдің бірінші және екінші заңдарымен сәйкес, F1-дің әр аталық және аналық гаметаларында, аллельдердің келесі үйлесімділігімен гаметалардың түзілуі мүмкін:
R тек қана Y немесе y-мен үйлесімділікте кездесе алады (бірақ r-мен емес), яғни RY немесе Ry түрінде.
r тек қана Y немесе y-мен үйлесімділікте кездесе алады (бірақ R-мен емес), яғни rY немесе ry түрінде.
Осылайша, кез-келген гаметаға осында көрсетілген аллельдер үйлесімділігінің төртеуінің қандай да бір біреуін алу мүмкіндігі 4-тен 1-ге тең.

24.4. А суреті. Гомозиготалы ата-ана дарақтары арасындағы шағылысудан алынған F1 фенотиптерінің қалыптасуы. Бұл дигибридті шағылысудың мысалы – контрасттаушы белгілердің екі жұбы қарастырылады. Б. F2 фенотиптерінің түзілуі кезіндегі барлық мүмкін гаметалар үйлесімділігін көрсету үшін Пеннет торының қолданылуы.
Қанша дегенмен моногибридті шағылысуда F2 ұрпағының ¾-інде доминантты аллель, ал ¼-інде− рецессивті аллель байқалатындықтан, біздермен қарастырылған 4 аллельдің F2-де байқалу мүмкіндігі мынаған тең:
тегіс (доминантты) ¾
сары (доминантты) ¾
қыртысты (рецессивті) ¼
жасыл (рецессивті) ¼
Бұдан аллельдердің келесі үйлесімділігінің F2 фенотипінде байқалуы мынаған тең:
тегіс және сары = ¾ × ¾ = 9/16
тегіс және сары = ¾ × ¼ = 3/16
қыртысты және сары = ¼ × ¾ = 3/16
қыртысты және жасыл = ¼ × ¼ = 1/16
Альтернативті белгілердің екі жұбы бойынша ажыратылатын сұрыптарды шағылысуы бойынша жүргізілген Мендельдің тәжірибелерінің нәтижелері, осы теориялық есептеулер нәтижелерімен жақын.
тегіс (доминантты) ¾
сары (доминантты) ¾
қыртысты (рецессивті) ¼
жасыл (рецессивті) ¼
Бұдан аллельдердің келесі үйлесімділігінің F2 фенотипінде байқалуы мынаған тең:
тегіс және сары = ¾ × ¾ = 9/16
тегіс және сары = ¾ × ¼ = 3/16
қыртысты және сары = ¼ × ¾ = 3/16
қыртысты және жасыл = ¼ × ¼ = 1/16
Альтернативті белгілердің екі жұбы бойынша ажыратылатын сұрыптарды шағылысуы бойынша жүргізілген Мендельдің тәжірибелерінің нәтижелері, осы теориялық есептеулер нәтижелерімен жақын.

24.1.4. Мендельдің заңдарының қысқаша баяндамасы
Төменде тұжырымдалған ережелерде заманауи генетикада қабылданған терминдер қолданылады.
NB. Дигибридті шағылысу кезіндегі мұнда сипатталған шағылысу механизмі, келтірілген мысалдар және қарапайым қатынас 9:3:3:1, әр түрлі хромосомаларда орналасқан гендермен бақыланатын белгілерге ғана тән. Егер де гендер бір хромосомада орналасса, онда мұндай типтің тәуелсіз ажырауы барлық жағдайларда байқала бермейді (24.3 бөлімін қараңыз).
24.2. Тұқымқуалаушылықтың хромосомалық теориясы
Мендель өзінің зерттеулерінің нәтижелерін және өз гипотезаларын 1866 жылы көптеген елдердің ғылыми қоғамдарына жіберілетін «Брюн қоғамының табиғи зерттеушілерінің еңбектері» журналына жариялады. Алайда ғалымдар оның жаңалығының мағынасына баға бере алмады; мүмкін, бұл сол уақытта Мендельдің анықтамаларын қандай да бір гаметалардағы тұқымқуалайтын факторлар ата-анадан ұрпақтарына беріле алатын нақты құрылымдармен байланыстыруға болмайтындығымен түсіндіріледі.
ХІХ ғасырдың соңына қарай микроскоптардың оптикалық сипатының елеулі түрде жақсаруы және цитологиялық тәсілдердің жетілдірілуі арқасында гаметалардағы және зиготалардағы хромосомалардың тәртібін бақылау мүмкін болды. 1875 жылы Гертвиг теңіз кірпісінің жұмыртқаларын ұрықтандыру кезінде екі ядроның – спермий ядросының және жұмыртқа жасушасы ядросының бірігуі жүретіндігіне назар аударды. 1902 жылы Бовери ағза белгілерінің дамуының реттелуіндегі ядроның маңызды рөлін көрсетті, ал 1882 жылы Флемминг митоз процесіндегі хромосомалардың тәртібін сипаттады.
1900 жылы Мендельдің заңдары екінші рет бір уақытта дерлік үш ғалыммен ашылды. Олар Мендельдің заңдарын лайықты түрде бағалаған – де Фриз Корренс және Черман. Корренс Мендельдің қорытындыларын бізге үйреншікті формада екі заң түрінде тұжырымдады және Мендельдің тұқымқуалаушылық бірлігін сипаттау үшін қолданған «элемент» сөзінің орнына «фактор» терминін енгізді. Ал американдық зерттеуші Уильям Сэттон гаметалардың түзілуі және ұрықтану кезіндегі хромосомалардың тәртібі мен мендельдік тұқымқуалаушылық факторлардың берілуінің арасындағы таңғажайып ұқсастықты байқады (24.3 кестесі).
24.3 кестесі. Мейозда және ұрықтандыруда, Мендель болжамында болатын жағдайлардың сәйкестігі
Төменде тұжырымдалған ережелерде заманауи генетикада қабылданған терминдер қолданылады.
- Берілген ағзаның әр белгісі аллельдер жұбымен бақыланады.
- Егер ағза құрамында берілген белгі бойынша екі әр түрлі аллель болса, онда олардың біреуі (доминантты) екінші (рецессивті) белгінің байқалуын толығымен баса отырып, байқала алады.
- Мейоз кезінде аллельдердің әр жұбы бөлінеді (ажырайды) және әр гамета аллельдердің әр жұбынан бір-бірден алады (ажырау заңы).
- Аталық және аналық гаметалар түзілуі кезінде олардың әрқайсысына бір жұптан кез-келген аллель, басқа жұптан кез-келген басқасымен бірге тұсе алады (тәуелсіз ажырау заңы).
- Әр аллель ұрпақтан-ұрпаққа дискретті өзгермейтін бірлік ретінде беріледі.
- Әр ағза ата-аналық дарақтардың әрқайсысынан бір аллельден (әр белгіге) иеленеді.
NB. Дигибридті шағылысу кезіндегі мұнда сипатталған шағылысу механизмі, келтірілген мысалдар және қарапайым қатынас 9:3:3:1, әр түрлі хромосомаларда орналасқан гендермен бақыланатын белгілерге ғана тән. Егер де гендер бір хромосомада орналасса, онда мұндай типтің тәуелсіз ажырауы барлық жағдайларда байқала бермейді (24.3 бөлімін қараңыз).
24.2. Тұқымқуалаушылықтың хромосомалық теориясы
Мендель өзінің зерттеулерінің нәтижелерін және өз гипотезаларын 1866 жылы көптеген елдердің ғылыми қоғамдарына жіберілетін «Брюн қоғамының табиғи зерттеушілерінің еңбектері» журналына жариялады. Алайда ғалымдар оның жаңалығының мағынасына баға бере алмады; мүмкін, бұл сол уақытта Мендельдің анықтамаларын қандай да бір гаметалардағы тұқымқуалайтын факторлар ата-анадан ұрпақтарына беріле алатын нақты құрылымдармен байланыстыруға болмайтындығымен түсіндіріледі.
ХІХ ғасырдың соңына қарай микроскоптардың оптикалық сипатының елеулі түрде жақсаруы және цитологиялық тәсілдердің жетілдірілуі арқасында гаметалардағы және зиготалардағы хромосомалардың тәртібін бақылау мүмкін болды. 1875 жылы Гертвиг теңіз кірпісінің жұмыртқаларын ұрықтандыру кезінде екі ядроның – спермий ядросының және жұмыртқа жасушасы ядросының бірігуі жүретіндігіне назар аударды. 1902 жылы Бовери ағза белгілерінің дамуының реттелуіндегі ядроның маңызды рөлін көрсетті, ал 1882 жылы Флемминг митоз процесіндегі хромосомалардың тәртібін сипаттады.
1900 жылы Мендельдің заңдары екінші рет бір уақытта дерлік үш ғалыммен ашылды. Олар Мендельдің заңдарын лайықты түрде бағалаған – де Фриз Корренс және Черман. Корренс Мендельдің қорытындыларын бізге үйреншікті формада екі заң түрінде тұжырымдады және Мендельдің тұқымқуалаушылық бірлігін сипаттау үшін қолданған «элемент» сөзінің орнына «фактор» терминін енгізді. Ал американдық зерттеуші Уильям Сэттон гаметалардың түзілуі және ұрықтану кезіндегі хромосомалардың тәртібі мен мендельдік тұқымқуалаушылық факторлардың берілуінің арасындағы таңғажайып ұқсастықты байқады (24.3 кестесі).
24.3 кестесі. Мейозда және ұрықтандыруда, Мендель болжамында болатын жағдайлардың сәйкестігі

24.5 суреті. Екі гомологиялық хромосомалар жұбы бар жасуша. Екі әр-түрлі гендік локустардың орналасуы қара дөңгелектермен көрсетілген. Берілген жағдайда екі локус гомологиялық хромосомалардың әр-түрлі жұптарында орналасқан, және әр ген екі аллельмен ұсынылған.
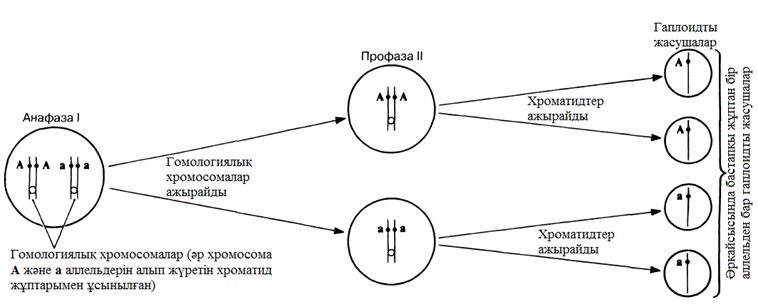
24.6 суреті. Мейоздағы гомологиялық хромосомалардың ажырау нәтижесі ретінде, А және а факторларының (аллельдерінің) мендельдік ажырау заңын түсіндіру.
Жоғарыда баяндалған барлық анықтамалар негізінде Сэттон және Бовери хромосомалардың мендельдік фактордың тасымалдаушысы ретінде қызмет атқаратындығы туралы болжам айтты, және тұқымқуалаушылықтың хромосомалық теориясы деп аталатын теорияны тұжырымдады. Бұл теорияға сәйкес, факторлардың әр жұбы гомологиялық хромосомалардың жұбында орналасқан, сонымен қоса әр хромосома бұл факторлардан біреуден алып жүреді. Қанша дегенмен кез-келген ағзадағы белгілер саны оның микроскоп астынан көрінетін хромосомаларының санынан бірнеше есе көп болғандықтан, әр хромосома құрамында көптеген факторлар болуы қажет.
1909 жылы Иогансен тұқымқуалаушылықтың негізгі бірлігін білдіретін фактор терминін ген терминімен алмастырды. Геннің, оның фенотипте көрінуін анықтайтын альтернативті формаларын аллельдер деп атады. Аллельдер – бұл ген түсіндіріле алатын нақты формалар; олар гомологиялық хромосомаларда бірдей орынды (локусты) алады (24.5 суреті).
Мендельдің факторлардың ажырау заңын енді мейоздың анафаза І-інде болатын гомологиялық хромосомалардың ажырауымен, және аллельдердің гаметалар арасындағы кездейсоқ бөлінуімен түсіндіруге болады. Бұл оқиғалар схемалық түрде 24.6 суретінде ұсынылған.
24.2.1. Тәуелсіз ажыраудың негізі ретінде хромосомалардың тәртібі
Мендельдің тәуелсіз ажырау заңын да мейоз кезіндегі хромосомалардың орын ауыстыруының ерекшеліктері ретінде түсіндіруге болады. Гаметалар түзілуі кезінде, олардың арасында берілген гомологиялық хромосомалар жұптарынан алынған аллельдердің ажырауы, басқа жұптардан алынған аллельдердің ажырауынан толықтай тәуелсіз түрде болады (24.7 суреті). Мейоздың метафаза І-індегі веретено экваторында гомологиялық хромосомалардың кездейсоқ орналасуы және олардың кейінгі анафаза І-дегі ажырауы, гаметалардағы аллельдердің рекомбинацияларының әртүрлілігіне әкеледі. Аталық немесе аналық гаметалардағы мүмкін үйлесімділіктердің санын 2n-нің ортақ формуласы бойынша анықтауға болады, мұндағы n – хромосомалардың гаплоидты саны. Адамда n = 23, ал әртүрлі үйлесімдердің мүмкін саны 223=8 388 608-ді құрайды.
1909 жылы Иогансен тұқымқуалаушылықтың негізгі бірлігін білдіретін фактор терминін ген терминімен алмастырды. Геннің, оның фенотипте көрінуін анықтайтын альтернативті формаларын аллельдер деп атады. Аллельдер – бұл ген түсіндіріле алатын нақты формалар; олар гомологиялық хромосомаларда бірдей орынды (локусты) алады (24.5 суреті).
Мендельдің факторлардың ажырау заңын енді мейоздың анафаза І-інде болатын гомологиялық хромосомалардың ажырауымен, және аллельдердің гаметалар арасындағы кездейсоқ бөлінуімен түсіндіруге болады. Бұл оқиғалар схемалық түрде 24.6 суретінде ұсынылған.
24.2.1. Тәуелсіз ажыраудың негізі ретінде хромосомалардың тәртібі
Мендельдің тәуелсіз ажырау заңын да мейоз кезіндегі хромосомалардың орын ауыстыруының ерекшеліктері ретінде түсіндіруге болады. Гаметалар түзілуі кезінде, олардың арасында берілген гомологиялық хромосомалар жұптарынан алынған аллельдердің ажырауы, басқа жұптардан алынған аллельдердің ажырауынан толықтай тәуелсіз түрде болады (24.7 суреті). Мейоздың метафаза І-індегі веретено экваторында гомологиялық хромосомалардың кездейсоқ орналасуы және олардың кейінгі анафаза І-дегі ажырауы, гаметалардағы аллельдердің рекомбинацияларының әртүрлілігіне әкеледі. Аталық немесе аналық гаметалардағы мүмкін үйлесімділіктердің санын 2n-нің ортақ формуласы бойынша анықтауға болады, мұндағы n – хромосомалардың гаплоидты саны. Адамда n = 23, ал әртүрлі үйлесімдердің мүмкін саны 223=8 388 608-ді құрайды.
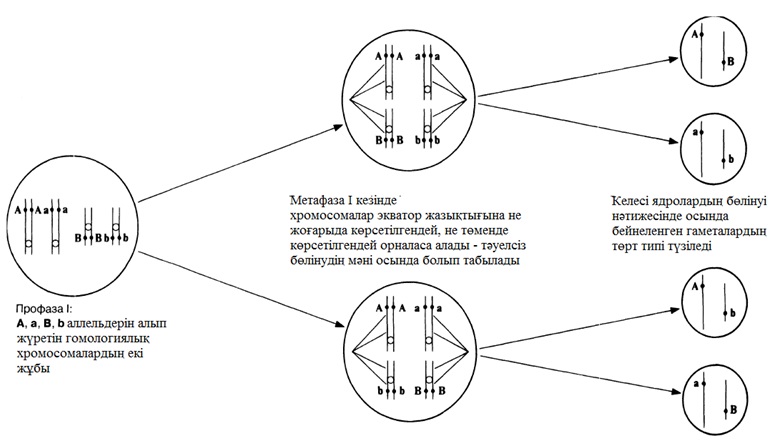
24.7 суреті. Мейоздағы гомологиялық хромосомалардың әр-түрлі жұптарының тәуелсіз ажырауының нәтижесі ретінде, А, а, В, b факторларының (аллельдерінің) тәуелсіз бөлінуінің мендельдік заңын түсіндіру (23.16 суретімен салыстырыңыз).

24.3. Тіркес
Бұған дейін осы тарауда талқыланған барлық жағдайлар және мысалдар, әртүрлі хромосомаларда орналасатын гендердің тұқымқуалауына қатысты болды. Цитологиялық зерттеулер деректеріне сәйкес, адамда барлық сомалық жасушалар құрамында 46 хромосомадан болады. Қанша дегенмен адам қан тобы, көздің түсі, инсулинді секреттеу қабілеті және т.б. сияқты мыңдаған әр-түрлі белгілерге ие болғандықтан, әр хромосомада гендердің көп саны болуы керек.
Бір хромосомада орналасқан гендерді тіркескен деп атайды. Бір хромосоманың барлық гендері тіркес тобын құрайды; олар әдетте бір гаметаға түседі және бірге тұқымқуалайды. Осылайша тіркестің бір тобына жататын гендер, әдетте Мендельдің тәуелсіз ажырау заңына бағынбайды. Сондықтан дигибридті шағылысу кезінде бұл гендер күтілетін қатынас 9:3:3:1-ді бермейді. Мұндай жағдайларда ең әр-түрлі қатынастар алынады, оларды енді бізге Мендельмен ашылған заңдылықтар белгілі болған соң біршама оңай түсіндіруге болады. (Мұнда тағы да бір айта кететін нәрсе, Мендельге белгілер жұптарының тұқымқуылауын зерттеу үшін, әр-түрлі хромосомаларда орналасқан гендерді таңдаудың сәті түсті.) Дрозофилада дене түсін және қанат ұзындығын қадағалайтын гендер, келесі аллеломорфтарға (әр-түрлі аллельдермен анықталатын фенотиптік белгілер) ие: сұр дене - қара дене, ұзын қанаттар – өскін (қысқа) қанаттар. Сұр дене және ұзын қанаттар басымдылық көрсетеді. Сұр денелі және ұзын қанатты гомозиготалы шыбынды және қара денелі және өскін қанатты гомозиготалы шыбынды шағылыстырған кезде F2-дегі күтілетін фенотиптер қатынасы, 9:3:3:1-ді құрауы керек. Бұл әр-түрлі гомологиялық емес хромосомаларда орналасатын дене түсі және қанат ұзындығы гендерінің кездейсоқ ажырауымен қамтамасыз етілетін дигибридті шағылыстырудағы қарапайым мендельдік тұқымқуалауды көрсеткен болар еді. Алайда оның орнына F2-де негізінен шамамен 3:1 қатынасындағы ата-аналық фенотиптер алынған болатын. Мұны, дене түсі және қанат зындығы гендерін бір хромосомада орналасқан, яғни тіркескен деп болжау арқылы түсіндіруге болады.
Бұған дейін осы тарауда талқыланған барлық жағдайлар және мысалдар, әртүрлі хромосомаларда орналасатын гендердің тұқымқуалауына қатысты болды. Цитологиялық зерттеулер деректеріне сәйкес, адамда барлық сомалық жасушалар құрамында 46 хромосомадан болады. Қанша дегенмен адам қан тобы, көздің түсі, инсулинді секреттеу қабілеті және т.б. сияқты мыңдаған әр-түрлі белгілерге ие болғандықтан, әр хромосомада гендердің көп саны болуы керек.
Бір хромосомада орналасқан гендерді тіркескен деп атайды. Бір хромосоманың барлық гендері тіркес тобын құрайды; олар әдетте бір гаметаға түседі және бірге тұқымқуалайды. Осылайша тіркестің бір тобына жататын гендер, әдетте Мендельдің тәуелсіз ажырау заңына бағынбайды. Сондықтан дигибридті шағылысу кезінде бұл гендер күтілетін қатынас 9:3:3:1-ді бермейді. Мұндай жағдайларда ең әр-түрлі қатынастар алынады, оларды енді бізге Мендельмен ашылған заңдылықтар белгілі болған соң біршама оңай түсіндіруге болады. (Мұнда тағы да бір айта кететін нәрсе, Мендельге белгілер жұптарының тұқымқуылауын зерттеу үшін, әр-түрлі хромосомаларда орналасқан гендерді таңдаудың сәті түсті.) Дрозофилада дене түсін және қанат ұзындығын қадағалайтын гендер, келесі аллеломорфтарға (әр-түрлі аллельдермен анықталатын фенотиптік белгілер) ие: сұр дене - қара дене, ұзын қанаттар – өскін (қысқа) қанаттар. Сұр дене және ұзын қанаттар басымдылық көрсетеді. Сұр денелі және ұзын қанатты гомозиготалы шыбынды және қара денелі және өскін қанатты гомозиготалы шыбынды шағылыстырған кезде F2-дегі күтілетін фенотиптер қатынасы, 9:3:3:1-ді құрауы керек. Бұл әр-түрлі гомологиялық емес хромосомаларда орналасатын дене түсі және қанат ұзындығы гендерінің кездейсоқ ажырауымен қамтамасыз етілетін дигибридті шағылыстырудағы қарапайым мендельдік тұқымқуалауды көрсеткен болар еді. Алайда оның орнына F2-де негізінен шамамен 3:1 қатынасындағы ата-аналық фенотиптер алынған болатын. Мұны, дене түсі және қанат зындығы гендерін бір хромосомада орналасқан, яғни тіркескен деп болжау арқылы түсіндіруге болады.
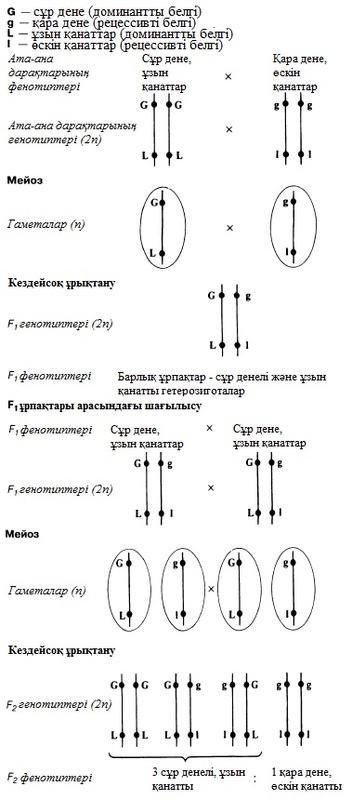
24.8 суреті. F2 фенотиптері арасындағы 3:1 қатынасын тіркес нәтижесі ретінде генетикалық түсіндіру.
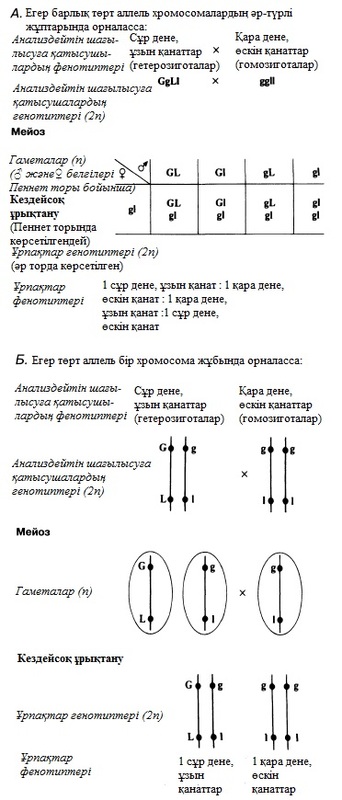
24.9 суреті. Морган болжамдарының генетикалық түсіндірмесі.
Іс жүзінде, алайда, 3:1қатынасы ешқашан шықпайды, ал F2-де шарасыз барлық төрт фенотип байқалады. Бұл толық тіркестің өте сирек кездесетіндігімен түсіндіріледі. Шағылыстыру бойынша көптеген тәжірибелерде тіркестер болған кезде ата-аналық фенотипті шыбындардан бөлек, белгілердің жаңа үйлесімділігімен дарақтар табылады. Бұл жаңа фенотиптерді рекомбинантты деп атайды. Бұның барлығы тіркестің келесі анықтамасына мүмкіндік береді: егер жаңа генді комбинациялы ұрпақтар (рекомбинанттар) ата-аналық фенотиптерге қарағанда сирегірек кездесетін болса, екі немесе одан да көп гендерді тіркескен деп атайды.
Американдық зерттеуші Томас Морганды тіркестің ашылуына әкелген оқиғаларды оның сұр ұзынқанатты гетерозигота (F1 ұрпағынан 24.8 суретінде ұсынылған шағылысудан) және қара терілі және өскін қанатты рецессивті гомозигота арасындағы анализдейтін шағылыстырудың нәтижелерін болжаған, дрозофиладағы тәжірибелерінің бірімен түсіндіруге болады. Екі мүмкін нәтиже ұсынылған болатын.
1 қара дене, ұзын қанаттар: 1 сұр дене, өскін қанаттар.
2. Егер дене түсін және қанаттар ұзындығын анықтайтын аллельдер, бір хромосома жұбына тиесілі болса (яғни тіркескен), онда фенотиптер қатынасы басқа болады:
1 сұр дене, ұзын қанаттар: 1 қара дене, өскін қанаттар.
Бұл болжамдардың түсіндірмесі 24.9 суретінде ұсынылған.
Морган бұл анализдейтін шағылыстыруды бірнеше рет өткізді және бір ретте болжанған нәтижелерді алған жоқ. Әр ретте ол келесі нәтижелерді алды:
41,5% − сұр дене, ұзын қанаттар
41,5% − қара дене, өскін қанаттар
8,5% − сұр дене, өскін қанаттар
8,5% −қана дене, ұзын қанаттар
Бұл нәтижелер негізінде Морган,
Ұрпақтардың 17%-ындағы аллельдердің рекомбинанты үйлесімінің байқалуы 4-пункт негізінде түсіндірілді. Бұл құбылыс кроссинговер (қиылыс) атауын алды.
Американдық зерттеуші Томас Морганды тіркестің ашылуына әкелген оқиғаларды оның сұр ұзынқанатты гетерозигота (F1 ұрпағынан 24.8 суретінде ұсынылған шағылысудан) және қара терілі және өскін қанатты рецессивті гомозигота арасындағы анализдейтін шағылыстырудың нәтижелерін болжаған, дрозофиладағы тәжірибелерінің бірімен түсіндіруге болады. Екі мүмкін нәтиже ұсынылған болатын.
- Егер дененің сұр немесе қара түсін және ұзын немесе өскін қанаттардыанықтайтын аллельдердің екі жұбы, әр түрлі хромосомаларда жататын болса (яғни тіркеспеген), онда олар фенотиптердің келесі қатынасында тәуелсіз ажырауы қажет:
1 қара дене, ұзын қанаттар: 1 сұр дене, өскін қанаттар.
2. Егер дене түсін және қанаттар ұзындығын анықтайтын аллельдер, бір хромосома жұбына тиесілі болса (яғни тіркескен), онда фенотиптер қатынасы басқа болады:
1 сұр дене, ұзын қанаттар: 1 қара дене, өскін қанаттар.
Бұл болжамдардың түсіндірмесі 24.9 суретінде ұсынылған.
Морган бұл анализдейтін шағылыстыруды бірнеше рет өткізді және бір ретте болжанған нәтижелерді алған жоқ. Әр ретте ол келесі нәтижелерді алды:
41,5% − сұр дене, ұзын қанаттар
41,5% − қара дене, өскін қанаттар
8,5% − сұр дене, өскін қанаттар
8,5% −қана дене, ұзын қанаттар
Бұл нәтижелер негізінде Морган,
- зерттелетін гендер хромосомаларда орналасқан;
- екі ген де бір хромосомада орналасқан, яғни тіркескен;
- әр геннің аллельдері гомологиялық хромосомаларда орналасқан;
- мейоз уақытында гомологиялық хромосомалар арасында аллельдермен алмасу жүреді,-деп жариялады.
Ұрпақтардың 17%-ындағы аллельдердің рекомбинанты үйлесімінің байқалуы 4-пункт негізінде түсіндірілді. Бұл құбылыс кроссинговер (қиылыс) атауын алды.

24.3.1. Кроссинговер және рекомбинациялар жиілігі
1909 жылы бельгиялық цитолог Янсенс мейоздың профаза І-індегі хиазмалардың түзілуін байқады (23.4 бөлімі).Бұл процесстің генетикалық шығу тегін Морган кроссинговерді (аллельдермен алмасу) хиазмалар түзілуі кезіндегі айырылу және гомологиялық хромосомалардың рекомбинациясы нәтижесінде болатындығы туралы ойын айта отырып түсіндірді. Мұнан былай микроскоптық зерттеулер деректерді рекомбинантты фенотиптер қатынасы туралы деректермен салыстыру, генетикалық материалмен алмасудың іс жүзінде барлық гомозиготалы хромосомалар арасында болатындығын дәлелдеді. Ата-аналық дарақтардағы тіркестер тобына кіретін аллельдер, бөлінеді және гаметаларға түсетін жаңа үйлесімдер түзеді, бұл процесс генетикалық рекомбинация деп аталады. Мұндай гаметалардан аллельдердің жаңа үйлесімділігімен пайда болатын ұрпақтарды рекомбинанттар, немесе кроссоверлер деп атайды. Осылайша кроссиговер популяцияларда бақыланатын генетикалық өзгергіштіктің маңызды қайнар көзі болып табылады.
Кроссинговер процесін сипаттау үшін хиазмалар түзілуі кезіндегі, терінің сұр түсінің және ұзын қанаттардың (екі аллель де доминантты) және терінің қара түсінің және өскін қанаттардың (екі аллель де рецессивті) аллельдерін алып жүретін дрозофилданың гомологиялық хромосомалары жұптарының тәртібін қарастыруға болады. Гетерозиготалы сұр ұзынқанатты аталық және гомозиготалы қара өскін қанатты аналық арасындағы шағылысу F1-де сұр денелі және ұзын қанатты гетерозиготалы ұрпақтар берді (24.10 суреті).
F1 ұрпағынан алынған екі рецессивті ген бойынша гомозиготалы шыбындарды анализдейтін шағылыстыру нәтижесінде келесі нәтижелер алынды:
Ата-аналық фенотиптер сұр дене, ұзын қанаттар 965
қара дене, өскін қанаттар 944
Рекомбинантты фенотиптер қара дене, ұзын қанаттар 206
сұр дене, өскін қанаттар 185
Бұл қорытындылар көрсететіндей, дене түсін және қанаттар ұзындығын анықтайтын гендер тіркескен. (Егер де бұл гендер әр-түрлі хромосомаларда орналасса және сондықтан кездейсоқ түрде бөлінетін болса, онда F1-дің гетерозиготасын гомозиготамен шағылыстырғанда екі рецессивті белгі бойынша фенотиптер қатынасы 1:1:1:1 болатындығын еске саламыз.) Келтірілген шамадан, дене түсін және қанаттар ұзындығын анықтайтын гендер рекомбинациясының жиілігін есептеуге болады.
1909 жылы бельгиялық цитолог Янсенс мейоздың профаза І-індегі хиазмалардың түзілуін байқады (23.4 бөлімі).Бұл процесстің генетикалық шығу тегін Морган кроссинговерді (аллельдермен алмасу) хиазмалар түзілуі кезіндегі айырылу және гомологиялық хромосомалардың рекомбинациясы нәтижесінде болатындығы туралы ойын айта отырып түсіндірді. Мұнан былай микроскоптық зерттеулер деректерді рекомбинантты фенотиптер қатынасы туралы деректермен салыстыру, генетикалық материалмен алмасудың іс жүзінде барлық гомозиготалы хромосомалар арасында болатындығын дәлелдеді. Ата-аналық дарақтардағы тіркестер тобына кіретін аллельдер, бөлінеді және гаметаларға түсетін жаңа үйлесімдер түзеді, бұл процесс генетикалық рекомбинация деп аталады. Мұндай гаметалардан аллельдердің жаңа үйлесімділігімен пайда болатын ұрпақтарды рекомбинанттар, немесе кроссоверлер деп атайды. Осылайша кроссиговер популяцияларда бақыланатын генетикалық өзгергіштіктің маңызды қайнар көзі болып табылады.
Кроссинговер процесін сипаттау үшін хиазмалар түзілуі кезіндегі, терінің сұр түсінің және ұзын қанаттардың (екі аллель де доминантты) және терінің қара түсінің және өскін қанаттардың (екі аллель де рецессивті) аллельдерін алып жүретін дрозофилданың гомологиялық хромосомалары жұптарының тәртібін қарастыруға болады. Гетерозиготалы сұр ұзынқанатты аталық және гомозиготалы қара өскін қанатты аналық арасындағы шағылысу F1-де сұр денелі және ұзын қанатты гетерозиготалы ұрпақтар берді (24.10 суреті).
F1 ұрпағынан алынған екі рецессивті ген бойынша гомозиготалы шыбындарды анализдейтін шағылыстыру нәтижесінде келесі нәтижелер алынды:
Ата-аналық фенотиптер сұр дене, ұзын қанаттар 965
қара дене, өскін қанаттар 944
Рекомбинантты фенотиптер қара дене, ұзын қанаттар 206
сұр дене, өскін қанаттар 185
Бұл қорытындылар көрсететіндей, дене түсін және қанаттар ұзындығын анықтайтын гендер тіркескен. (Егер де бұл гендер әр-түрлі хромосомаларда орналасса және сондықтан кездейсоқ түрде бөлінетін болса, онда F1-дің гетерозиготасын гомозиготамен шағылыстырғанда екі рецессивті белгі бойынша фенотиптер қатынасы 1:1:1:1 болатындығын еске саламыз.) Келтірілген шамадан, дене түсін және қанаттар ұзындығын анықтайтын гендер рекомбинациясының жиілігін есептеуге болады.
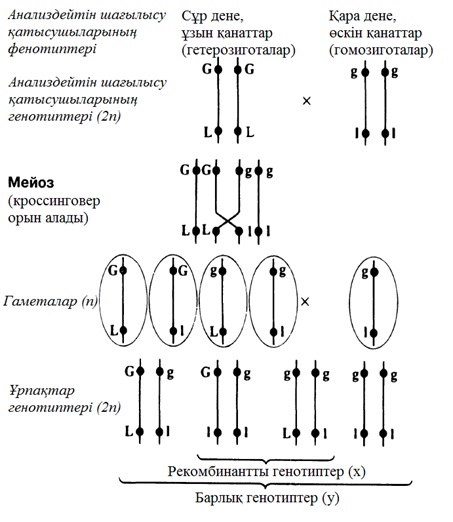
24.10 суреті. Кроссинговердің және рекомбинантты генотиптердің пайда болуының генетикалық түсіндірмесі. Рекомбинация (х) және дарақтардың жалпы саны (у) анықталатын дарақтар санын есептей отырып, рекомбинация жиілігін мына формула бойынша анықтауға болады:
Рекомбинациялар жиілігі (%) = х/у × 100
Рекомбинациялар жиілігі мына формуламен есептеледі

Біздің мысалда рекомбинация жиілігі мынаған тең
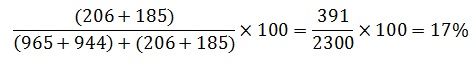
Бұл мағына гаметалар түзілуі кезінде болатын рекомбинациялар санына сәйкес келеді.Морганның шәкірттерінің бірі А. Х. Стертевант, рекомбинациялар жиілігінің гендердің хромосома бойымен сызықтық орналасуы туралы дәлелдейтіндігі жөнінде ой айтты. Стерванттың тағы да маңыздырақ болжамы, рекомбинациялар жиілігі гендердің хромосомалардағы салыстырмалы орналасуын көрсететіндігінде: тіркескен гендер бір-бірінен қаншалықты алыс орналасса, олардың арасында кроссинговер болуының мүмкіндігі көбірек, яғни кроссинговер жиілігі соншалықты жоғары (24.11 суреті).

24.11 суреті. Үш гендік локус – А, В және С – бір хромосомада орналасқан. Кроссинговер және бөліну жиілігі В және С гендері үшін, А және В гендеріне қарағанда жоғарырақ, өйткені кроссинговер жиілігі гендер арасындағы арақашықтыққа байланысты.
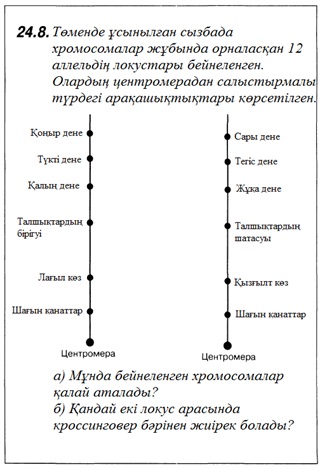
24.4. Генетикалық карталар
Рекомбинациялар жиілігі туралы мәліметтер ең алдымен олар генетиктерге гендердің хромосомалардағы салыстырмалы орналасу картасын құру мүмкіндігін беретіндігі үшін маңызды. Хромосомалардың карталары кроссинговер жиілігінің хромосомадағы болжанған арақашықтыққа тікелец көшіру жолымен құрылады. А және В гендері арасындағы 4%-ға тең кроссинговер жиілігі, бұл гендердің бір хромосомада 4 генетикалық бірлік (морганид) қашықтығында орналасқандығын білдіреді. Егер А және С гендері арасындағы кроссинговер жиілігі 9%-ға тең болса, онда оларды 9 морганидке тең қашықтық бөліп тұрады. Алайда, бұл мәліметтер А, В және С гендерінің орналасу реттілігі туралы еш нәрсе айтпайды (24.12 суреті).
Рекомбинациялар жиілігі туралы мәліметтер ең алдымен олар генетиктерге гендердің хромосомалардағы салыстырмалы орналасу картасын құру мүмкіндігін беретіндігі үшін маңызды. Хромосомалардың карталары кроссинговер жиілігінің хромосомадағы болжанған арақашықтыққа тікелец көшіру жолымен құрылады. А және В гендері арасындағы 4%-ға тең кроссинговер жиілігі, бұл гендердің бір хромосомада 4 генетикалық бірлік (морганид) қашықтығында орналасқандығын білдіреді. Егер А және С гендері арасындағы кроссинговер жиілігі 9%-ға тең болса, онда оларды 9 морганидке тең қашықтық бөліп тұрады. Алайда, бұл мәліметтер А, В және С гендерінің орналасу реттілігі туралы еш нәрсе айтпайды (24.12 суреті).
24.12 суреті. А, В және С гендік локустарының өзара орналасуын, мәтінде баяндалған ақпараттар негізінде анықтау.
Іс жүзінде әдетте кроссинговер жиілігін ең аз дегенде үш генге бір мезгілде анықтайды; бұл триангуляция деп аталатын тәсіл тек қана гендер арасындағы қашықты ғана емес, сонымен қатар олардың реттілігін анықтауға мүмкіндік береді. Мысалға P, Q, R және S, 4 геннің қатысуымен тәжірибелік шағылыстырулар қатары нәтижесінде орнатылған кроссинговерлер жиілігін қарастырайық:
P – Q = 24%
R – P = 14%
R – S = 8%
S – P = 6%
Гендердің реттілігін және олардың арасындағы қашықтықты орнату үшін, хромосоманы суреттейтін сызық сызады және төменде тізбектелген қимылдарды жасайды.
P – Q = 24%
R – P = 14%
R – S = 8%
S – P = 6%
Гендердің реттілігін және олардың арасындағы қашықтықты орнату үшін, хромосоманы суреттейтін сызық сызады және төменде тізбектелген қимылдарды жасайды.
24.13 суреті. P, Q, R және S гендерінің хромосомадағы орналасуын анықтау үшін триангуляция тәсілін қолдану
24.14. А суреті. Бірі А, В және С доминантты аллельдерін алып жүретін, ал басқасы а,b және c рецессивті аллельдерін алып жүретін гомологиялық хроматидтер жұбы. Кроссинговер екі нүктеде болады - *1 және *2. Б. Хромотидтер ажырауының нәтижесі: мұндағы аллельдер реттілігі басқа, дегенмен гендік локустардың реттілігі және олардың арасындағы арақашықтық бұрынғы қалпында қалады.
- Хромосоманың ортасына ең аз рекомбинация жиілігі бар гендерді орналастырады, яғни S – P = 6% (24.13, І сурет).
- Көлемі бойынша келесі рекомбинация жиілігін таңдайды, яғни R – S = 8%, және S-ке қатысты R-дің хромосомадағы екі мүмкін орнын көрсетеді (24.13,ІІ суреті).
- Дәл осылай келесі рекомбинация жиілігіне жасайды, яғни R – P = 14% (24.3, ІІІ суреті). Сонымен қатар R-дің P-ден оң жақта орналаса алмайтындығы айқындалады.
- Дәл осылай P – Q = 24%-ке де қайталайды (24.13, ІV суреті). Q орнын қосымша ақпаратсыз анықтауға болмайды. Егер, мысалы, Q – R үшін рекомбинация жиілігі 10% болса, онда бұл Q-генінің хромосоманың сол жағындағы ұшында орналасқандығын айқындайды.

24.5. Тіркес топтары жіне хромосомалар
Бұл тарауда баяндалған ақпараттар тұқымқуалау механизмі туралы бірте-бірте жинақталған білімді көрсетеді. Ғасырдың басында жүргізілген генетикалық зерттеулер, негізінен белгілердің тасымалдануындағы геннің рөлін айқындауға бағытталды. Жеміс шыбынында (Drosophila melanogaster) жүргізілген Морганның жұмыстары, фенотиптік белгілердің көбісі бұл шыбында төрт топқа бөлінгендігін және әр топқа кіретін белгілер бірге тұқымқуалайтындығын көрсетті. Тіркес топтарының санының хромосома жұптары санына сәйкес келетіндігі байқалды.
Басқа ағзаларды зерттеу ұқсас нәтижелерге әкелді. Алуан түрлі ағзаларды тәжірибелік шағылыстыруда, кейбір тіркес топтары басқаларынан көп екендігі анықталды (яғни, оларға көбірек ген кіреді). Бұл ағзалардың хромосомасын зерттеу, олардың әр-түрлі ұзындыққа ие екендігін көрсетті. Морган бұл бақылаклар арасындағы анық байланыстың барын дәлелдеді. Олар гендердің хромосомалардағы орналасуына алдағы дәлелдемелер қызметін атқарады.
24.5.1. Алып хромосомалар және гендер
1913 жылы Стертевант дрозофила хромосомаларындағы гендерді картаға түсіру бойынша өз жұмысын бастады, бірақ бұл хромосомалардағы айырылатын құрылымдарды гендермен байланыстыру мүмкіндігі пайда болғанға дейін 21 жыл бұрын болды. 1934 жылы дрозофиланың сілекей безіндегі жасушаларда басқа сомалық жасушаларға қарағанда хромосомалардың 100 есе ірі екендігі байқалды. Қандай да бір себептермен бұл хромосомалар бірнеше рет екі еселенеді, бірақ олардың қаз-қатар жататын бірнеше мыңы жиналмайынша бір-бірінен ажырамайды. Хромосомаларды бояп және оларды микроскоп астында зерттеу арқылы, олардың кезектесетін ашық және қою жолақтардан тұратындығын көруге болады (24.15 суреті). Әр хромосомаға өзінің ерекше жолақтар жиынтығы тән. Басында бұл жолақтарды гендер деп болжады немесе оған сенді, алайда жағдай олай емес болып шықты. Дрозофилада жасанды түрде микроскоп астында көрінетін көлденең жолақтардың сипатындағы белгілі бір өзгерістермен қатар жүретін әр-түрлі фенотиптік ауытқулар тудыруға болады. Бұл фенотиптік және хромосомалық ауытқулар өз кезегінде тәжірибелік шағылыстырудағы рекомбинациялар жиілігі туралы мәліметтер бойынша құрылған хромосомалық карталарда белгіленген гендік локустармен арақатынас орнатады. Хромосомалардағы жолақтар гендердің орнын көрсетеді деп айтуға болады, бірақ бұл гендердің өзі емес.
Бұл тарауда баяндалған ақпараттар тұқымқуалау механизмі туралы бірте-бірте жинақталған білімді көрсетеді. Ғасырдың басында жүргізілген генетикалық зерттеулер, негізінен белгілердің тасымалдануындағы геннің рөлін айқындауға бағытталды. Жеміс шыбынында (Drosophila melanogaster) жүргізілген Морганның жұмыстары, фенотиптік белгілердің көбісі бұл шыбында төрт топқа бөлінгендігін және әр топқа кіретін белгілер бірге тұқымқуалайтындығын көрсетті. Тіркес топтарының санының хромосома жұптары санына сәйкес келетіндігі байқалды.
Басқа ағзаларды зерттеу ұқсас нәтижелерге әкелді. Алуан түрлі ағзаларды тәжірибелік шағылыстыруда, кейбір тіркес топтары басқаларынан көп екендігі анықталды (яғни, оларға көбірек ген кіреді). Бұл ағзалардың хромосомасын зерттеу, олардың әр-түрлі ұзындыққа ие екендігін көрсетті. Морган бұл бақылаклар арасындағы анық байланыстың барын дәлелдеді. Олар гендердің хромосомалардағы орналасуына алдағы дәлелдемелер қызметін атқарады.
24.5.1. Алып хромосомалар және гендер
1913 жылы Стертевант дрозофила хромосомаларындағы гендерді картаға түсіру бойынша өз жұмысын бастады, бірақ бұл хромосомалардағы айырылатын құрылымдарды гендермен байланыстыру мүмкіндігі пайда болғанға дейін 21 жыл бұрын болды. 1934 жылы дрозофиланың сілекей безіндегі жасушаларда басқа сомалық жасушаларға қарағанда хромосомалардың 100 есе ірі екендігі байқалды. Қандай да бір себептермен бұл хромосомалар бірнеше рет екі еселенеді, бірақ олардың қаз-қатар жататын бірнеше мыңы жиналмайынша бір-бірінен ажырамайды. Хромосомаларды бояп және оларды микроскоп астында зерттеу арқылы, олардың кезектесетін ашық және қою жолақтардан тұратындығын көруге болады (24.15 суреті). Әр хромосомаға өзінің ерекше жолақтар жиынтығы тән. Басында бұл жолақтарды гендер деп болжады немесе оған сенді, алайда жағдай олай емес болып шықты. Дрозофилада жасанды түрде микроскоп астында көрінетін көлденең жолақтардың сипатындағы белгілі бір өзгерістермен қатар жүретін әр-түрлі фенотиптік ауытқулар тудыруға болады. Бұл фенотиптік және хромосомалық ауытқулар өз кезегінде тәжірибелік шағылыстырудағы рекомбинациялар жиілігі туралы мәліметтер бойынша құрылған хромосомалық карталарда белгіленген гендік локустармен арақатынас орнатады. Хромосомалардағы жолақтар гендердің орнын көрсетеді деп айтуға болады, бірақ бұл гендердің өзі емес.
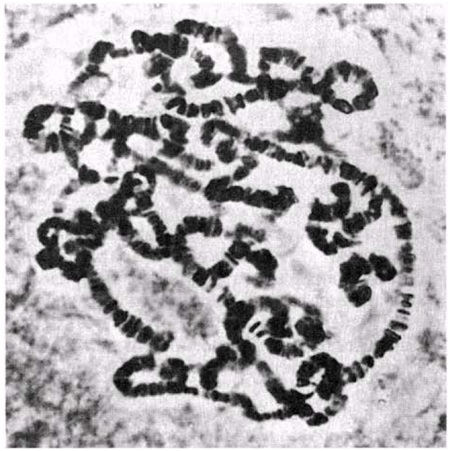
24.15 суреті. Drosophila melanogaster-дің сілекей бездерінің жасушаларынан алынған алып хромосомалар. Өздерінің центромералары арқылы байланысқан хромосомалардың төрт жұбы көрінеді.
24.6. Жынысты анықтау
Жоғарыда сипатталған ағзалардың фенотиптік белгілері және олардың хромосомаларының құрылымы арасындағы тәуелділік анықтамасының әсіресе нақты мысалы ретінде жынысты анықтау қызмет етеді. Дрозофилада екі жыныс арасындағы фенотиптік айырмашылықтар, хромосомалардағы айырмашылықтармен анық байланысты (24.16 суреті). Аталық және аналық жануарлар қатарының кариотипін зерттеу кезінде олардың арасында кейбір айырмашылықтар анықталды. Аталықтағы сияқты, аналық дарақтарда да барлық жасушаларда бірдей (гомологиялық) хромосомалар жұптары болады, бірақ хромосомалардың бір жұбы бойынша олар айырылады. Бұл жыныс хромосомалары (гетеросомалар). Барлық қалған хромосомаларды аутосомалар деп атайды. 24.16 суретінде көрінетіндей, дрозофилада төрт жұп хромосома бар. Үш жұп (ІІ, ІІІ және ІV) екі жыныста да бірдей, бірақ аналықта бірдей хромосомадан тұратын І-інші жұп, аталықтағы І-інші жұптан ерекшеленеді. Бұл жұптың хромосомалары X және Y-хромосомалар деп аталады; аналық генотипі XX, ал аталық генотип – XY. Жыныстық хромосомалар арасындағы мұндай айырмашылықтар көптеген жануарларғка және адамға тән (24.17 суреті). Құстарда және көбелектерде кері құбылыс байқалады: аналықтарда XY хромосомалар, ал аталықтарда XX хромосомалар бар. Кейбір жәндіктерде, мысалы турақанаттыларда Y-хромосома мүлдем болмайды, сондықтан да аталықтың жыныс жасушалары жалғыз X-хромосоманы алып жүреді (аталық генотипі XO).
Жоғарыда сипатталған ағзалардың фенотиптік белгілері және олардың хромосомаларының құрылымы арасындағы тәуелділік анықтамасының әсіресе нақты мысалы ретінде жынысты анықтау қызмет етеді. Дрозофилада екі жыныс арасындағы фенотиптік айырмашылықтар, хромосомалардағы айырмашылықтармен анық байланысты (24.16 суреті). Аталық және аналық жануарлар қатарының кариотипін зерттеу кезінде олардың арасында кейбір айырмашылықтар анықталды. Аталықтағы сияқты, аналық дарақтарда да барлық жасушаларда бірдей (гомологиялық) хромосомалар жұптары болады, бірақ хромосомалардың бір жұбы бойынша олар айырылады. Бұл жыныс хромосомалары (гетеросомалар). Барлық қалған хромосомаларды аутосомалар деп атайды. 24.16 суретінде көрінетіндей, дрозофилада төрт жұп хромосома бар. Үш жұп (ІІ, ІІІ және ІV) екі жыныста да бірдей, бірақ аналықта бірдей хромосомадан тұратын І-інші жұп, аталықтағы І-інші жұптан ерекшеленеді. Бұл жұптың хромосомалары X және Y-хромосомалар деп аталады; аналық генотипі XX, ал аталық генотип – XY. Жыныстық хромосомалар арасындағы мұндай айырмашылықтар көптеген жануарларғка және адамға тән (24.17 суреті). Құстарда және көбелектерде кері құбылыс байқалады: аналықтарда XY хромосомалар, ал аталықтарда XX хромосомалар бар. Кейбір жәндіктерде, мысалы турақанаттыларда Y-хромосома мүлдем болмайды, сондықтан да аталықтың жыныс жасушалары жалғыз X-хромосоманы алып жүреді (аталық генотипі XO).

24.16 суреті. Drosophila melanogaster-дің аталығы және аналығының хромосомалары. Хромосомалардың төрт жұбы көрсетілген. І жұп – жыныс хромосомалары.
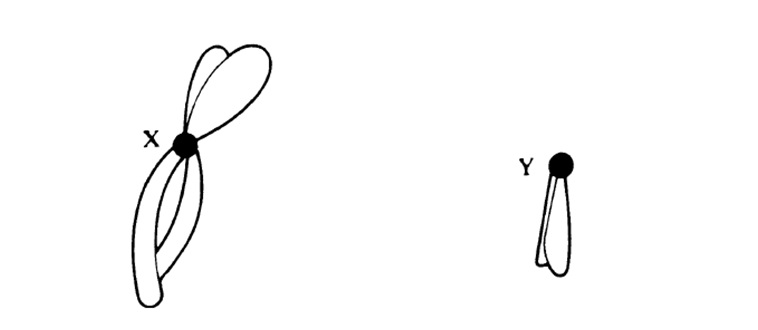
24.17 суреті. Мейоз метафазасындағы адамның жыныс хромосомалары.

24.18 суреті. Адамдағы жыныстар қатынасының генетикалық түсіндірмесі.
Гаметогенезде жыныстық хромосомалар бойынша қарапайым мендельдік ажырау байқалады. Мысалы, сүтқоректілерде әр жұмыртқа жасушасының құрамында бір X-хромосома бар; аталықтарда спермийдің жартысы X-хромосоманы, ал екінші жартысы бір Y-хромосоманы алып жүреді (24.18 суреті). Ұрықтың жынысы жұмыртқа жасушасын қай спермий ұрықтандыратындығына байланысты. XX-генотипінде құрамында тек Х-хромосомалар болатын бірдей гаметалар түзілетін болғандықтан дарақты гомогаметалы деп атайды, ал XY генотипінде гаметалардың жартысының құрамында X-, екінші жартысында – Y-хромосома болатындықтан гетерогаметалы деп атайды. Адамда тұлғаның генотиптік жынысын, беттің ішкі жағындағы эпителий жасушаларын микроскоп астында зерттей отырып анықтауға болады. Мұндай жасушаларда X-хромосомалардың біреуі белсенді емес, сұйықтығы ажыратылған жағдайда болады, және тығыз қаралау Барр денешігі деп аталатын денешік түріне ие. Барр денешіктерінің саны әрқашан бар Х-хромосомаларынан бірге кіші, яғни аталықта (XY) олар мүлде жоқ, ал аналықта (ХХ) ол тек біреу. Y-хромосоманың функциясы жануар түріне қарай ауытқитындығы белгілі. Адамда Y-хромосома алда жыныс мүшелерінің және аталық фенотиптің дамуына әсер ететін атабездің жетілуін қадағалайды (21.7.4 бөлім). Кейбір ағзаларда, алайда, Y-хромосома құрамында жынысты анықтайтын гендер болмайды. Оны онда өте аз ген болғандықтан, генетикалық инертті немесе бос деп атайды. Дрозофилада аталық белгілерді анықтайтын гендер аутосомаларда орналасатыны, және олардың фенотиптік әсері Х-хромосомалар жұбының болуымен жасырылатыны болжанады. Аталық белгілер тек қана бір Х-хромосоманың қатысуымен көрінеді. Бұл жыныспен шектелген тұқымқуалаушылықтың (жыныспен тіркескен тұқымқуалаушылыққа кері) үлгісі, мысалы әйел адамда сақалдың өсуін анықтайтын гендер басылады.
24.19. А және Б суреті. Морганмен жүргізілген қызылкөзді және ақкөзді дрозофилалар арасындағы реципрокты шағылысу. Ақ көздердің пайда болуының төмен жиілігіне назар аударыңыз. В. Қызыл көзді аталық және қызылкөзді (гетерозиготалы) F1 аналық арасындағы шағылысу. Г. Ақ көзді аталық және қызыл көзді (гетерозиготалы) F1 аналық арасындағы шағылысу. Ақ көз белгісі осы аллель бойынша гомозиготалы аналықтарда ғана байқалатындығына назар аударыңыз.
Морган және оның қызметкерлері дрозофиладағы көз түсінің тұқымқуалауы ата-аналық дарақтардың жынысына байланысты болатындығын байқаған. Көздің қызыл түсі аққа басымдылық көрсетеді. Қызыл көзді аталық және ақ қөзді аналықты шағылыстырғанда F1-де бірдей мөлшерде қызылкөз аналықтар және ақкөз аталықтар алынды (24.19, А суреті). Алайда ақкөзді аталықты қызылкөзді аналықпен шағылыстырғанда F1-де бірдей мөлшерде қызылкөзді аталықтар және ақкөді аналықтар алынды (24.19, Б суреті). Бұл F1 шыбындарды өзара шағылыстырғанда қызылкөзді аналықтар, қызылкөзді және ақкөзді аталықтар алынды, бірақ бірде-бір ақкөзді аналықтар алынған жоқ (24.19, В суреті). Аталықтарда рецессивті белгінің байқалу жиілігі аналықтарға қарағанда көп екендігі, ақкөзділікті анықтайтын рецессивті аллель Х-хромосомада орналасатындығы, ал Y-хромосоманың көз бояуының генінен айырылғандығы туралы ойға әкелді. Бұл гипотезаны тексеру үшін, Морган бастапқы ақкөзді аталықты, F1-дің қызыл көзді аналығымен шағылыстырды (24.19, Г суреті). Ұрпақтар қызылкөзді және ақкөзді аталықтар және аналықтардан құралды. Сондықтан да Морган көздің түсін анықтайтын генді тек Х-хромосома алып жүретіндігі, ал Y-хромосомада сәйкес локус жоқтығы туралы әділ тұжырым жасады. Бұл құбылыс жыныспен тіркескен тұқымқуалау атымен белгілі.


24.6.1. Жыныспен тіркескен тұқымқуалау
Жыныстық хромосомаларда орналасқан гендерді жыныспен тіркес деп атайды. Х-хромосомадағы гетерогаметалы дарақтарда, Y-хромосомасында гомолог жоқ аумақтар бар (24.20 суреті). Сондықтан аталық жынысты дарақтардағы осы аймақтық гендерімен анықталатын белгілері, олар рецессивті болғанның өзінде байқалады. Тіркестің бұл ерекше формасы жыныспен тіркескен белгілердің тұқымқуалауын түсіндіруге мүмкіндік береді, мысалы адамдағы түсті ажыратпау, ерте шаштың түсуі және гемофилия. Гемофилия – жыныспен тіркескен рецессивті белгі; гемофилия кезінде қанның ұюының күшеюінде маңызды рөл ойнайтын VIII факторының түзілуі бұзылады. VIII факторының синтезін анықтайтын ген, Х-хромосоманың гомологы жоқ аймағында орналасады, және екі аллельмен – қалыпты (доминантты) және мутантты (рецессивті) аллельмен ұсынылған. Келесі генотиптер мен фенотиптер мүмкін:
Генотип Фенотип
ХНХН Сау әйел
ХНХһ Сау әйел (тасымалдаушы)
ХНҮ Сау еркек
ХһҮ Гемофилиясы бар еркек
Жыныстық хромосомаларда орналасқан гендерді жыныспен тіркес деп атайды. Х-хромосомадағы гетерогаметалы дарақтарда, Y-хромосомасында гомолог жоқ аумақтар бар (24.20 суреті). Сондықтан аталық жынысты дарақтардағы осы аймақтық гендерімен анықталатын белгілері, олар рецессивті болғанның өзінде байқалады. Тіркестің бұл ерекше формасы жыныспен тіркескен белгілердің тұқымқуалауын түсіндіруге мүмкіндік береді, мысалы адамдағы түсті ажыратпау, ерте шаштың түсуі және гемофилия. Гемофилия – жыныспен тіркескен рецессивті белгі; гемофилия кезінде қанның ұюының күшеюінде маңызды рөл ойнайтын VIII факторының түзілуі бұзылады. VIII факторының синтезін анықтайтын ген, Х-хромосоманың гомологы жоқ аймағында орналасады, және екі аллельмен – қалыпты (доминантты) және мутантты (рецессивті) аллельмен ұсынылған. Келесі генотиптер мен фенотиптер мүмкін:
Генотип Фенотип
ХНХН Сау әйел
ХНХһ Сау әйел (тасымалдаушы)
ХНҮ Сау еркек
ХһҮ Гемофилиясы бар еркек
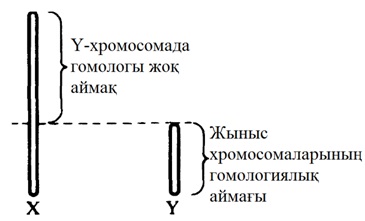
24.20 суреті. Жыныс хромосомаларының гомологиялық және гомологиялық емес аймақтары.
Жынысқа тіркескен кез-келген белгі бойынша гетерозиготалы аналық жыныс дарақтарын, сәйкес рецессивті геннің тасымалдаушысы деп атайды. Олар фенотиптік түрде қалыпты, бірақ олардың гаметаларының жартысы рецессивті генді алып жүреді. Әкесіндегі қалыпты қанға қарамастан, тасымалдаушы аналардың ұлдары 50% (1/2) мүмкіндікпен гемофилиядан зардап шегетін болады. Мұндай тасымалдаушы әйел мен қалыпты еркектің некесінен әр-түрлі фенотипті балалар туылуы мүмкін (24.21 суреті).

24.21 суреті. Жыныспен тіркес гемофилия аллелінің тұқымқуалау механизмі.
Гемофилияның тұқымқуалауының ең жақсы құжатталған мысалдарының бірі – ағылшын патшайымы Викторияның ұрпақтарының шежіресі. Гемофилия гені патшайым Викторияда немесе оның ата-анасының біреуіндегі мутация нәтижесінде пайда болған деп болжанады. 24.22 суретінде бұл геннің оның ұрпақтарына қалай берілгендігі көрсетілген.
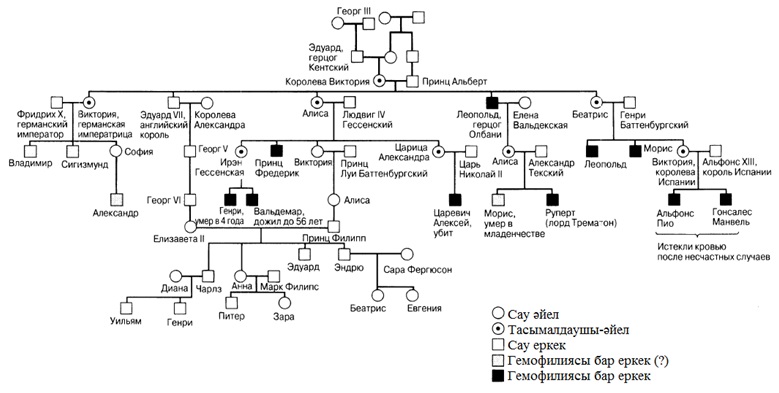
24.22 суреті. Виктория патшайымның ұрпақтарымен гемофилияның тұқымқуалауы.Сызбада тек қана гемофилияны тасымалдауға қатысқан немесе бұл аурумен зақымдалған ұрпақтар ұсынылған. Британдық корольдік шежіре, Виктория патшайымның ұрпақтарының бірде-біреуінде жеті ұрпақ бойына гемофилия байқалмағандығының неліктен екендігін көрсету үшін жалғастырылған.

24.7. Гендердің арасындағы өзара ара-қатынас
Бұл тарауда осы уақытқа дейін генетиканың салыстырмалы түрде қарапайым қырлары қарастырылды: басымдылық көрсету, моногибридті және дигибридті шағылысу, тіркес, жынысты анықтау және жыныспен тіркес. Алайда, гендер арасындағы басқа да өзара қатынастар белгілі, және дәл солар ағзаның фенотиптік белгілерінің көп бөлігін анықтауы мүмкін. Төменде бұл өзара қатынастардың кейбіреулері қарастырылатын болады.
24.7.1. Кодоминанттылық
Екі немесе одан да көп аллельдердің толығымен доминанттылық немесе рецессивтілік көрсетпейтін жағдайлар белгілі, себебі гетерозиготалы жағдайда олардың бір де біреуі екіншісіне басымдылық көрсетпейді. Бұл жағдай – кодоминанттылық немесе толық емес доминанттау – Мендельдің моногибридті шағылыстыру бойынша тәжірибелеріндегі жағдайдан айрықша болып табылады. Бақытына орай, Мендель кодоминанттылық тән емес белгілерді таңдады; кері жағдайда бұл оның ерте зерттеулерін қатты қиындатар еді.
Бұл тарауда осы уақытқа дейін генетиканың салыстырмалы түрде қарапайым қырлары қарастырылды: басымдылық көрсету, моногибридті және дигибридті шағылысу, тіркес, жынысты анықтау және жыныспен тіркес. Алайда, гендер арасындағы басқа да өзара қатынастар белгілі, және дәл солар ағзаның фенотиптік белгілерінің көп бөлігін анықтауы мүмкін. Төменде бұл өзара қатынастардың кейбіреулері қарастырылатын болады.
24.7.1. Кодоминанттылық
Екі немесе одан да көп аллельдердің толығымен доминанттылық немесе рецессивтілік көрсетпейтін жағдайлар белгілі, себебі гетерозиготалы жағдайда олардың бір де біреуі екіншісіне басымдылық көрсетпейді. Бұл жағдай – кодоминанттылық немесе толық емес доминанттау – Мендельдің моногибридті шағылыстыру бойынша тәжірибелеріндегі жағдайдан айрықша болып табылады. Бақытына орай, Мендель кодоминанттылық тән емес белгілерді таңдады; кері жағдайда бұл оның ерте зерттеулерін қатты қиындатар еді.

24.23 суреті. Андалуздық тұқымды тауықтарды алу (F1 будандары).
Кодоминанттылық өсімдіктерде де, жануарларда да бақыланады. Көптеген жағдайларда гетерозиготалар доминантты және рецессивті гомозиготалар фенотиптері арасындағы аралық фенотипке ие. Мысал ретінде бірыңғай қара және ақ жағал (ақ түсті фонда ұсақ көк дақтар бар, «шашыранды тауық» деп аталады) тауықтарды шағылыстыру нәтижесінде алынған андалузиялық тауықтар қызмет атқарады. Қара қауырсынды, меланиннің қара пигментінің синтезін анықтайтын аллель қамтамасыз етеді. «Шашыраңқы тауықтарда» бұл пигмент болмайды. Гетерозиготаларда меланин қауырсынның құлпырған көгілдір түсін жасай отырып тек қана шала синтезделеді.
Қанша дегенмен жартылай доминантты аллельдер үшін ортақ қабылданған белгілер болмағандықтан, шағылыстырулардың генетикалық нәтижелерін түсіндіру үшін аллельдерді белгілеуге арнайы белгілер енгізу қажет болды: қаралар – В, «шашыранды» − b, W, Bw немесе BBw . Гомозиготалы қара және «шашыранды» әтештер арасындағы шағылысу нәтижелері 24.23суретінде ұсынылған.
F1 ұрпақтарын өзара шағылыстырғанда F2-дегі фенотиптер қатынасы, моногибридті шағылысуға тән мендельдік 3:1 қатынасынан өзгеше болады. Бұл жағдайда, дарақтардың жартысында генотип F1 дарақтарындағыдай болатын 1:2:1 қатынасы шығады (24.24 суреті). 1:2:1 қатынасы кодоминанттаудағы шағылыстыру нәтижелеріне тән. Басқа мысалдар 24.4 кестесінде келтірілген.
Қанша дегенмен жартылай доминантты аллельдер үшін ортақ қабылданған белгілер болмағандықтан, шағылыстырулардың генетикалық нәтижелерін түсіндіру үшін аллельдерді белгілеуге арнайы белгілер енгізу қажет болды: қаралар – В, «шашыранды» − b, W, Bw немесе BBw . Гомозиготалы қара және «шашыранды» әтештер арасындағы шағылысу нәтижелері 24.23суретінде ұсынылған.
F1 ұрпақтарын өзара шағылыстырғанда F2-дегі фенотиптер қатынасы, моногибридті шағылысуға тән мендельдік 3:1 қатынасынан өзгеше болады. Бұл жағдайда, дарақтардың жартысында генотип F1 дарақтарындағыдай болатын 1:2:1 қатынасы шығады (24.24 суреті). 1:2:1 қатынасы кодоминанттаудағы шағылыстыру нәтижелеріне тән. Басқа мысалдар 24.4 кестесінде келтірілген.
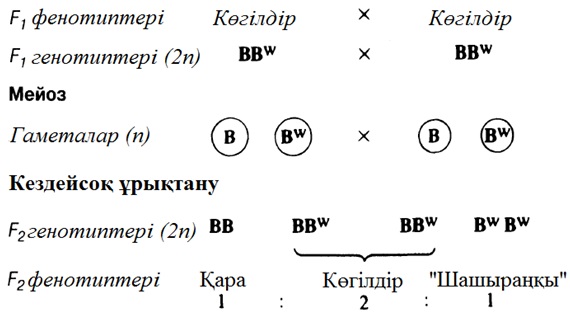
24.24 суреті. Андалуздық тұқым тауықтарын шағылыстырудың нәтижелері (F2 будандары).
24.4 кестесі. Кодоминанттылықтың мысалдары
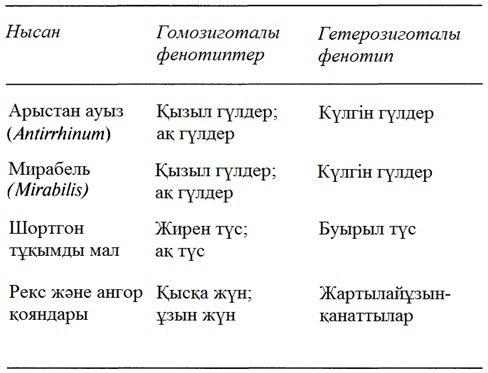
24.7.2. Көптік аллельдер
Осыған дейін қарастырылған барлық жағдайларда әр белгі, екі аллельдің біреуімен ұсыныла алатын бір генмен қадағаланады. Алайда кез-келген екеуі гомологиялық хромосомалардың бір локусында орналасатын үш және одан да көп аллельдермен қадағаланатын бірнеше әртүрлі формаларда бір белгі байқалатын көп мысалдар белгілі. Мұндай жағдайларда көптік аллельдер (немесе көптік аллеломорфтар) туралы айтады. Осылайша тышқандардың жүнінің түсі мен көзінің түсі және адамдарда қан тобы көптік аллельдермен қадағаланады.
Осыған дейін қарастырылған барлық жағдайларда әр белгі, екі аллельдің біреуімен ұсыныла алатын бір генмен қадағаланады. Алайда кез-келген екеуі гомологиялық хромосомалардың бір локусында орналасатын үш және одан да көп аллельдермен қадағаланатын бірнеше әртүрлі формаларда бір белгі байқалатын көп мысалдар белгілі. Мұндай жағдайларда көптік аллельдер (немесе көптік аллеломорфтар) туралы айтады. Осылайша тышқандардың жүнінің түсі мен көзінің түсі және адамдарда қан тобы көптік аллельдермен қадағаланады.

АВО қан тобының тұқымқуалауы
Қан топтары аутосомалық генмен қадағаланады. Бұл геннің локусын І әрпімен («изогемагглютиноген» сөзінен), ал оның үш аллельін – А, В және О әріптерімен белгілейді. А және В аллельдері бірдей деңгейде доминантты, ал О аллельі екуіне қатысты да рецессивті. 24.5 кестесінде ұсынылған генотиптер, қан топтарының фенотиптік білінуін анықтайды. Қанда бір доминантты аллель болса, антидене сияқты әрекет ететін агглютинин деп аталатын зат түзіледі. Мысалы ІAІO генотипінде эритроциттер мембранасында А агглютиногені түзіледі, ал плазма құрамында анти – В (қан А тобына сәйкес келеді) агглютиногені бар. Қан тобын анықтау тәсілдері туралы 14.9.7 бөлімінен қараңыз.
24.5 кестесі. Адамдағы қан топтарының генотиптері
Қан топтары аутосомалық генмен қадағаланады. Бұл геннің локусын І әрпімен («изогемагглютиноген» сөзінен), ал оның үш аллельін – А, В және О әріптерімен белгілейді. А және В аллельдері бірдей деңгейде доминантты, ал О аллельі екуіне қатысты да рецессивті. 24.5 кестесінде ұсынылған генотиптер, қан топтарының фенотиптік білінуін анықтайды. Қанда бір доминантты аллель болса, антидене сияқты әрекет ететін агглютинин деп аталатын зат түзіледі. Мысалы ІAІO генотипінде эритроциттер мембранасында А агглютиногені түзіледі, ал плазма құрамында анти – В (қан А тобына сәйкес келеді) агглютиногені бар. Қан тобын анықтау тәсілдері туралы 14.9.7 бөлімінен қараңыз.
24.5 кестесі. Адамдағы қан топтарының генотиптері
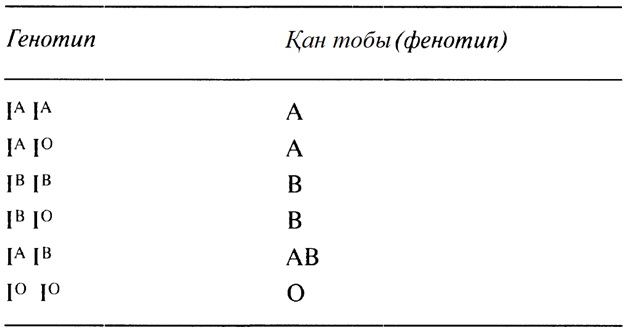

24.7.3. Летальды (өлімге душар етуші) гендер
Бір геннің бірнеше белгілерге, соның ішінде тіршілікке қабілеттілікке әсер ете алатын жағдайлар белгілі. Адамда және басқа сүтқоректілерде анықталған рецессивті ген туылғанда өлімге әкелетін, өкпенің ішкі жабысуының түзілуін тудырады. Басқа мысал ретінде, шеміршектің қалыптасуына әсер ететін және туа біткен кемтарлық тудыратын гендер қызмет етеді, олар ұрықтың немесе нәрестенің өліміне әкеледі.
Қауырсындардың «бұйралығын» тудыратын аллель бойынша гомозиготалы тауықтарда қауырсындардың толық дамымауы өзімен бірге бірнеше фенотиптік әсерлер алып жүреді. Мұндай тауықтарда жылу оқшаулау нашарлайды және олар жылу жоғалтудан қиналады. Бұл шығындар оларда құрылымдық және физиологиялық бейімделушіліктер жанында толтырылады, алайда олардың тиімділігі аз, сондықтан да бұл тауықтар арасында өлім-жітім көп.
Бір геннің бірнеше белгілерге, соның ішінде тіршілікке қабілеттілікке әсер ете алатын жағдайлар белгілі. Адамда және басқа сүтқоректілерде анықталған рецессивті ген туылғанда өлімге әкелетін, өкпенің ішкі жабысуының түзілуін тудырады. Басқа мысал ретінде, шеміршектің қалыптасуына әсер ететін және туа біткен кемтарлық тудыратын гендер қызмет етеді, олар ұрықтың немесе нәрестенің өліміне әкеледі.
Қауырсындардың «бұйралығын» тудыратын аллель бойынша гомозиготалы тауықтарда қауырсындардың толық дамымауы өзімен бірге бірнеше фенотиптік әсерлер алып жүреді. Мұндай тауықтарда жылу оқшаулау нашарлайды және олар жылу жоғалтудан қиналады. Бұл шығындар оларда құрылымдық және физиологиялық бейімделушіліктер жанында толтырылады, алайда олардың тиімділігі аз, сондықтан да бұл тауықтар арасында өлім-жітім көп.

24.25 суреті. Тышқандардағы жүннің түсінің тұқымқуалауының генетикалық түсіндірмесі.Шағылыстыру нәтижелері ҮҮ генотипінің летальды екендігін дәлелдейді.
Летальды геннің әсер етуінің айқын мысалы ретінде тышқандарда жүннің түсінің тұқымқуалауы қызмет атқарады. Агути сияқты жабайы тышқандардың жүні әдетте сұр, бірақ кейбір тышқандардың жүні сары болады. Сары тышқандар арасындағы шағылысуда сары тышқандармен қатар агути де, 2:1 қатынасындай алынады. Мұндай нәтижелердің жалғыз мүмкін түсіндірмесі мынада, жүннің сары түсі агутиге басымдылық көрсетеді және барлық сары тышқанда гетерозиготалы. Ауытқитын байланыс, гомозиготалы сары тышқандардың туылғанға дейін өлетіндігімен түсіндіріледі (24.25 суреті). Сары аталықтармен шағылыстырылған жүкті сары аналықтарды ашқан кезде, олардың жатырында өлі сары ұрықтар табылды. Ал сары тышқандар мен агутиді шағылыстырғанда жүкті аналықтардың жатырында өлі сары ұрықтар табылған жоқ, қанша дегенмен мұндай шағылысуда жүннің сары түсінің гені (ҮҮ) бойынша гомозиготалы ұрпақтар болуы мүмкін емес.
24.7.4. Гендік кешендер
Кейде бір гендік локуста толығымен бір фенотиптік белгіні қадағалайтын аллельдер жұбы орналасады, алайда мұндай жағдайлар өте сирек. Белгілердің көпшілігі гендік кешен түзетін бірнеше гендердің ара-қатынасымен анықталады. Мысалы, белгі әр-түрлі локустарда орналасатын өзара әрекеттесетін екі немесе одан да көп гендермен қадағалануы мүмкін. Осылай, әтештердегі айдар пішінінің тұқымқуалауы, екі локуста және оған қоса әр-түрлі хромосомаларда орналасатын гендермен қадағаланады; бұл гендердің өзара әрекеттесуі нәтижесінде төрт әр-түрлі фенотип туады, олар бұршақ тәрізді, раушан тәрізді, жаңғақ тәрізді және қарапайым айдар (24.26 суреті). Бұршақ тәрізді және раушан тәрізді айдарлардың дамуы доминантты аллельдің (сәйкесінше P немесе R) басқа доминантты аллельдің жоқтығы әсерінен болуымен анықталады. Жаңғақ тәрізді айдар аз мөлшерде бұршақ тәрізді және раушан тәрізді аллельдерден (яғни PR генотипінде) бір-бірден болған кезде, толық емес доминанттаудың модификацияланған түрінің нәтижесінде түзіледі. Қарапайым айдар екі рецессивті ген бойынша (яғни pprr) гомозиготаларда ғана дамиды. Бұл фенотиптер және генотиптер 24.6 кестесінде ұсынылған.
24.6 кестесі. Әр-түрлі пішінді айдарлары бар тауықтардың фенотиптері және мүмкін генотиптері
24.7.4. Гендік кешендер
Кейде бір гендік локуста толығымен бір фенотиптік белгіні қадағалайтын аллельдер жұбы орналасады, алайда мұндай жағдайлар өте сирек. Белгілердің көпшілігі гендік кешен түзетін бірнеше гендердің ара-қатынасымен анықталады. Мысалы, белгі әр-түрлі локустарда орналасатын өзара әрекеттесетін екі немесе одан да көп гендермен қадағалануы мүмкін. Осылай, әтештердегі айдар пішінінің тұқымқуалауы, екі локуста және оған қоса әр-түрлі хромосомаларда орналасатын гендермен қадағаланады; бұл гендердің өзара әрекеттесуі нәтижесінде төрт әр-түрлі фенотип туады, олар бұршақ тәрізді, раушан тәрізді, жаңғақ тәрізді және қарапайым айдар (24.26 суреті). Бұршақ тәрізді және раушан тәрізді айдарлардың дамуы доминантты аллельдің (сәйкесінше P немесе R) басқа доминантты аллельдің жоқтығы әсерінен болуымен анықталады. Жаңғақ тәрізді айдар аз мөлшерде бұршақ тәрізді және раушан тәрізді аллельдерден (яғни PR генотипінде) бір-бірден болған кезде, толық емес доминанттаудың модификацияланған түрінің нәтижесінде түзіледі. Қарапайым айдар екі рецессивті ген бойынша (яғни pprr) гомозиготаларда ғана дамиды. Бұл фенотиптер және генотиптер 24.6 кестесінде ұсынылған.
24.6 кестесі. Әр-түрлі пішінді айдарлары бар тауықтардың фенотиптері және мүмкін генотиптері

Бұршақтәрізді айдары бар таза тұқымды тауықтың және раушантәрізді айдары бар таза тұқымды әтештің арасындағы шағылымтыру нәтижесінде алынатын F2 генотиптері және F2-дегі фенотиптер ара қатынасы 24.27 суретінде көрсетілген.


24.26 суреті. Тауықтардағы айдарлар пішінінің өзгеруі. А. Қарапайым айдар. Б. Бұршақ тәрізді айдар. В. Раушан тәрізді айдар. Г. Жаңғақ тәрізді айдар.
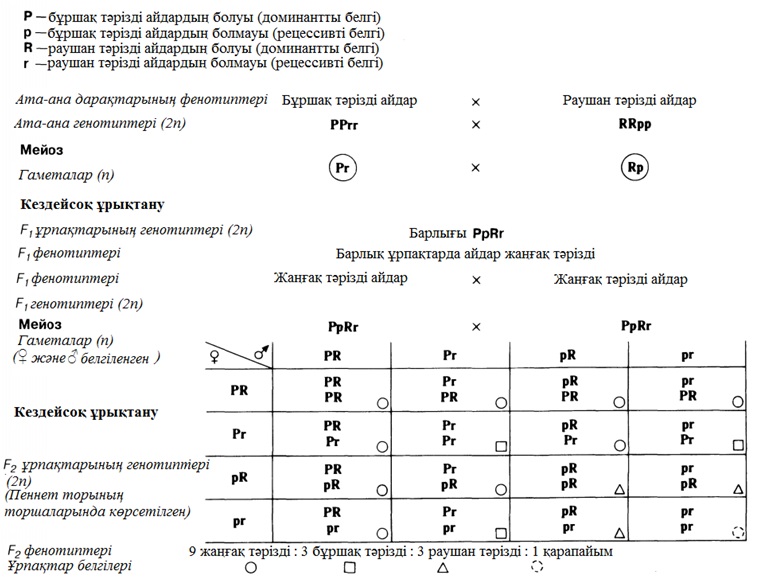
24.27 суреті. Тауықтардағы айдар пішінінің тұқымқуалауының генетикалық түсіндірмесі.
24.7.5. Эпистаз
Генді егер оның бар болуы басқа локуста орналасатын қандай-да бір геннің әсерін басатын болса, эпистатикалық (грек тілінен аударғанда epі – үстінде, жоғарыда, statis – аялдау, бөгеліс) гендерді кейде тежеуші гендер деп атайды, себебі олар гипостатикалық (грек тілінен аударғанда hypo – астында және statis) деп атайтын басқа гендерге әсер етеді.
Тышқандардағы жүннің түсі әр-түрлі локустарда орналасатын гендер жұбымен қадағаланады. Эпистатикалық ген бояудың болуын анықтайды және екі аллельге ие: боялған жүнді анықтайтын доминантты және альбинизмді (ақ түс) анықтайтын рецессивті. Гипостатикалық ген бояудың типін анықтайды және екі аллельге ие: агути (сұр түсті анықтайтын доминантты) және қара (рецессивті). Тышқандар өздерінің генотиптеріне қарай сұр және қара түске ие бола алады, бірақ бояудың байқалуы үшін, боялған жүннің аллельінің біруақыттағы болуы қажет. Альбинизмнің рецессивті аллельі бойынша гомозиготалы тышқандар, оларда агутидің және қара жүн аллельі болғанның өзінде альбинос болып қалады. Үш әр түрлі фенотип мүмкін: агути, қара жүн және альбинизм. Шағылыстыруларда бұл фенотиптерді, шағылысатын дарақтар генотипіне қарай әр-түрлі қатынаста алуға болады (24.28 суреті және 24.7 кестесі).
Генді егер оның бар болуы басқа локуста орналасатын қандай-да бір геннің әсерін басатын болса, эпистатикалық (грек тілінен аударғанда epі – үстінде, жоғарыда, statis – аялдау, бөгеліс) гендерді кейде тежеуші гендер деп атайды, себебі олар гипостатикалық (грек тілінен аударғанда hypo – астында және statis) деп атайтын басқа гендерге әсер етеді.
Тышқандардағы жүннің түсі әр-түрлі локустарда орналасатын гендер жұбымен қадағаланады. Эпистатикалық ген бояудың болуын анықтайды және екі аллельге ие: боялған жүнді анықтайтын доминантты және альбинизмді (ақ түс) анықтайтын рецессивті. Гипостатикалық ген бояудың типін анықтайды және екі аллельге ие: агути (сұр түсті анықтайтын доминантты) және қара (рецессивті). Тышқандар өздерінің генотиптеріне қарай сұр және қара түске ие бола алады, бірақ бояудың байқалуы үшін, боялған жүннің аллельінің біруақыттағы болуы қажет. Альбинизмнің рецессивті аллельі бойынша гомозиготалы тышқандар, оларда агутидің және қара жүн аллельі болғанның өзінде альбинос болып қалады. Үш әр түрлі фенотип мүмкін: агути, қара жүн және альбинизм. Шағылыстыруларда бұл фенотиптерді, шағылысатын дарақтар генотипіне қарай әр-түрлі қатынаста алуға болады (24.28 суреті және 24.7 кестесі).
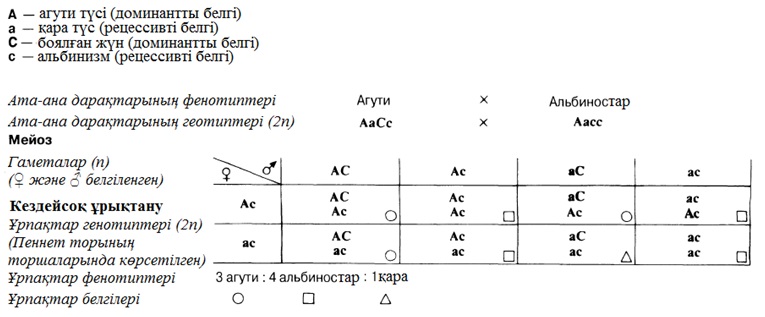
24.28 суреті. Тышқандардағы эпизтазада жүннің түсі бойынша фенотиптердің ерекше қатынасының генетикалық түсіндірмесі.
24.7 кестесі. Тышқандардағы жүн түсінің гендерінің эпистатикалық байланысуында ұрпақтарда туатын фенотиптердің кейбір қатынастары (сонымен қатар 24.28 суретін қараңыз)

24.7.6. Көпгенді тұқымқуалау
Ең айқын белгілердің көбісі әр-түрлі гендердің бірлескен әсерінің нәтижесі болып табылады. Бұл гендер көпгенді жүйе деп аталатын ерекше гендік кешен түзеді. Мұндай жүйенің әр жеке генінің үлесі, фенотиптерге қандай да бір елеулі әсер етуге өте аз, алайда олардың бірлескен әрекеті үздіксіз өзгергіштіктің генетикалық негізін құрайды.
24.8. Өзгергіштік
Өзгергіштік деп, бір популяцияға немесе түрге тиесілі ағзалар арасындағы сол немесе басқа белгі бойынша айырмашылықтар жиынтығын атайды. Берілген түр бойынша дарақтардың таңғажайып морфологиялық әртүрлілігі Дарвин және Уоллесті, олардың саяхаты кезінде қызықтырды. Тұқымқуалау бойынша мұндай заңды және дәлелденетін айырмашылықтардың тасымалдану сипаты Мендельдің зерттеулерінің негізі ретінде қызмет атқарды. Дарвин анықталған белгілердің іріктеу нәтижесінде дами алатындығын орнатқан (26.4.2 бөлімі) кезде, Мендель іріктеу жүргізілетін белгілердің ұрпақтан-ұрпаққа берілуін қамтамасыз ететін механизмді түсіндірді.
Мендель, даму кезінде фенотиптің морфологиялық, физиологиялық және биохимиялық ерекшеліктерінде байқалатын ағза генотипін, тұқымқуалайтын факторлардың қалай анықтайтынын сипаттады. Егер кез-келген белгінің фенотиптік байқалуы, ең соңында, бұл белгілерді қадағалайтын гендермен қамтамасыз етілсе, онда анықталған белгілердің даму деңгейіне орта үлкен әсер етуі мүмкін.
Ең айқын белгілердің көбісі әр-түрлі гендердің бірлескен әсерінің нәтижесі болып табылады. Бұл гендер көпгенді жүйе деп аталатын ерекше гендік кешен түзеді. Мұндай жүйенің әр жеке генінің үлесі, фенотиптерге қандай да бір елеулі әсер етуге өте аз, алайда олардың бірлескен әрекеті үздіксіз өзгергіштіктің генетикалық негізін құрайды.
24.8. Өзгергіштік
Өзгергіштік деп, бір популяцияға немесе түрге тиесілі ағзалар арасындағы сол немесе басқа белгі бойынша айырмашылықтар жиынтығын атайды. Берілген түр бойынша дарақтардың таңғажайып морфологиялық әртүрлілігі Дарвин және Уоллесті, олардың саяхаты кезінде қызықтырды. Тұқымқуалау бойынша мұндай заңды және дәлелденетін айырмашылықтардың тасымалдану сипаты Мендельдің зерттеулерінің негізі ретінде қызмет атқарды. Дарвин анықталған белгілердің іріктеу нәтижесінде дами алатындығын орнатқан (26.4.2 бөлімі) кезде, Мендель іріктеу жүргізілетін белгілердің ұрпақтан-ұрпаққа берілуін қамтамасыз ететін механизмді түсіндірді.
Мендель, даму кезінде фенотиптің морфологиялық, физиологиялық және биохимиялық ерекшеліктерінде байқалатын ағза генотипін, тұқымқуалайтын факторлардың қалай анықтайтынын сипаттады. Егер кез-келген белгінің фенотиптік байқалуы, ең соңында, бұл белгілерді қадағалайтын гендермен қамтамасыз етілсе, онда анықталған белгілердің даму деңгейіне орта үлкен әсер етуі мүмкін.
24.29 сурет. Дискретті (А) және үздіксіз (Б) өзгергіштік кезіндегі нұсқалар жиілігінің үлестірілуін көрсететін гистограммалар.
Кез-келген үлкен популяциядағы фенотиптік айырмашылықтарды зерттеу, өзгергіштіктің екі түрі болатындығын көрсетеді – дискретті және үздіксіз. Қандай да бір белгінің өзгергіштігін зерттеу үшін, мысалы адам бойын, бұл белгіні зерттелетін популяциядағы индивидуумдардың көп санында өлшеп көру керек. Өлшеулер нәтижелерін, популяциядағы бұл белгінің әр-түрлі нұсқаларының жиілігінің бөлінуін көрсететін гистограмма түрінде ұсынылады. 24.29 суретінде осындай зерттеулерде алынған қарапайым нәтижелер ұсынылған. Олар дискретті және үздіксіз өзгергіштік арасындағы айырмашылықты көрнекі білдіреді.
24.8.1. Дискретті өзгергіштік
Кейбір белгілер популяцияда нұсқалардың шектелген санымен ұсынылған. Мұндай жағдайларда дарақтар арасындағы айырмашылықтар анық көрсетілген, ал аралық формалар жоқ. Мұндай белгілерге , мысалы, мыналар жатады, адамның қан тобы, дрозофиланың қанаттарының ұзындығы, қайың мүркөбелегінің (Biston betularia) меланистік және ашық формалары, наурызгүлдің (Primula) аналық мойнының ұзындығы және жануарлардың және өсімдіктердің жынысы. Дискретті өзгергіштікке тән белгілер, әдетте екі немесе бірнеше аллельі бола алатын бір немесе екі басты гендермен қадағаланады. Сыртқы жағдайлар олардың фенотиптік экспрессиясына салыстырмалы түрде аз әсер етеді.
Қанша дегенмен, дискретті өзгергіштік кейбір анық көрінетін белгілермен шектелгендіктен, оны сонымен қатар сандық немесе үздіксіз өзгергіштікпен салыстырғанда сапалық өзгергіштік деп те атайды.
24.8.2. Үздіксіз өзгергіштік
Көптеген белгілер бойынша популяцияда бір шектен екіншісіне ауысудың толық қатары байқалады. Ең айқын мысалдар ретінде, масса, сызықтық өлшемдер, дененің толықтай немесе оның жеке бөліктерінің пішіні және түсі сияқты белгіле қызмет етеді. Үздіксіз өзгергіштікті көрсететін белгілер бойынша жиіліктің бөлінуі, қалыпты бөліну қисығына сәйкес келеді (2.3.3 қосымшасын қараңыз). Популяция мүшелерінің көбісі қисықтың орташа бөлігіне түседі, ал оның берілген белгінің екі шеткі маңызына сәйкес келетін соңында дарақтардың шамамен бірдей саны орналасады. Үздіксіз өзгергіштік тән белгілер, көптеген гендердің (полигендердің) және орта факторларының біріккен әрекетімен қамтамасыз етіледі. Бұл гендердің әрқайсысы фенотипке жеке түрде өте аз әсер етеді, бірақ олардың біріккен әсері өте елеулі.
24.8.3. Ортаның әсері
Кез-келген фенотиптік белгіні анықтайтын басты фактор – бұл генотип. Ағза генотипі ұрықтану кезінде анықталады, бірақ бұл генетикалық потенциялдың алдағы байқалуы, ағзаның дамуы кезінде оған әсер ететін сыртқы факторларға елеулі түрде байланысты. Осылай, мысалы, Мендельмен қолданылған ұзын сабақты асбұршақ сұрпы әдетте 180 см ұзындыққа жетті. Алайда бұл үшін жеткілікті жарықтандыру, сумен қамтамасыз ету және жақсы топырақ қажет болды. Оңтайлы жағдайлар болмағанда (шектеуші фактор болғанда) ұзын сабақтың гені өзінің әсерін толықтай түрде көрсете алған жоқ. Генотиптің және орта факторларының өзара әрекеттесу әсерін дат генетигі Иогансен көрсетті. Қысқа үрмебұршаққа жүргізілген тәжірибелер қатарында, ол бұл өздігінен тозаңданған өсімдіктердің әр ұрпағынан ең ауыр және ең жеңіл тұқымдарды таңдады және оларды келесі ұрпақты алу үшін өсірді. Бірнеше жылдар бойы бұл тәжірибелерді қайталай отырып, ол «ауыр» немесе «жеңіл» түрі шегінде тұқымдар орташа салмақ бойынша аз ажыратылған кезде әр-түрлі түр бойынша, яғни «ауыр» және «жеңіл» түрлер бойынша тұқымдардың орташа салмағындағы айырмашылықтар елеулі болғандығын анықтады. Бұл, белгінің фенотиптік байқалуына тұқымқуалаушылықпен қатар орта да әсер етеді деп санауға мүмкіндік береді. Бұл нәтижелер негізінде үздіксіз фенотиптік өзгергіштікті, «түрленетін генотипке әсер ететін өзгеретін орта факторларының жиынтық әсері» деп анықтауға болады. Бұл нәтижелер сонымен қатар берілген белгінің тұқымқуалау деңгейі бірінші кезекте генотиппен анықталатындығын көрсетеді. Даралық, темперамент және ақыл сияқты мұндай таза адамға тән қасиеттерге келетін болсақ, онда бар ақпараттарға қарағанда, олар тұқымқуалайтын және орта факторларына байланысты. Олар әр-түрлі индивидуумда әр-түрлі деңгейде бір-бірімен өзара әрекеттесе отырып, берілген белгінің ақырғы көрінісіне әсер етеді. Генетикалық және орта факторларындағы дәл осы айырмашылықтар адамдар арасындағы фенотиптік айырмашылықтарды құрайды. Бұл факторлардың қандай да бірі әрқашан басым болатындығы туралы нақты ақпараттар бізде жоқ, алайда орта ешқашан да генотиппен анықталған фенотипті шегінен тыс шығара алмайды (24.30 суреті).
24.8.1. Дискретті өзгергіштік
Кейбір белгілер популяцияда нұсқалардың шектелген санымен ұсынылған. Мұндай жағдайларда дарақтар арасындағы айырмашылықтар анық көрсетілген, ал аралық формалар жоқ. Мұндай белгілерге , мысалы, мыналар жатады, адамның қан тобы, дрозофиланың қанаттарының ұзындығы, қайың мүркөбелегінің (Biston betularia) меланистік және ашық формалары, наурызгүлдің (Primula) аналық мойнының ұзындығы және жануарлардың және өсімдіктердің жынысы. Дискретті өзгергіштікке тән белгілер, әдетте екі немесе бірнеше аллельі бола алатын бір немесе екі басты гендермен қадағаланады. Сыртқы жағдайлар олардың фенотиптік экспрессиясына салыстырмалы түрде аз әсер етеді.
Қанша дегенмен, дискретті өзгергіштік кейбір анық көрінетін белгілермен шектелгендіктен, оны сонымен қатар сандық немесе үздіксіз өзгергіштікпен салыстырғанда сапалық өзгергіштік деп те атайды.
24.8.2. Үздіксіз өзгергіштік
Көптеген белгілер бойынша популяцияда бір шектен екіншісіне ауысудың толық қатары байқалады. Ең айқын мысалдар ретінде, масса, сызықтық өлшемдер, дененің толықтай немесе оның жеке бөліктерінің пішіні және түсі сияқты белгіле қызмет етеді. Үздіксіз өзгергіштікті көрсететін белгілер бойынша жиіліктің бөлінуі, қалыпты бөліну қисығына сәйкес келеді (2.3.3 қосымшасын қараңыз). Популяция мүшелерінің көбісі қисықтың орташа бөлігіне түседі, ал оның берілген белгінің екі шеткі маңызына сәйкес келетін соңында дарақтардың шамамен бірдей саны орналасады. Үздіксіз өзгергіштік тән белгілер, көптеген гендердің (полигендердің) және орта факторларының біріккен әрекетімен қамтамасыз етіледі. Бұл гендердің әрқайсысы фенотипке жеке түрде өте аз әсер етеді, бірақ олардың біріккен әсері өте елеулі.
24.8.3. Ортаның әсері
Кез-келген фенотиптік белгіні анықтайтын басты фактор – бұл генотип. Ағза генотипі ұрықтану кезінде анықталады, бірақ бұл генетикалық потенциялдың алдағы байқалуы, ағзаның дамуы кезінде оған әсер ететін сыртқы факторларға елеулі түрде байланысты. Осылай, мысалы, Мендельмен қолданылған ұзын сабақты асбұршақ сұрпы әдетте 180 см ұзындыққа жетті. Алайда бұл үшін жеткілікті жарықтандыру, сумен қамтамасыз ету және жақсы топырақ қажет болды. Оңтайлы жағдайлар болмағанда (шектеуші фактор болғанда) ұзын сабақтың гені өзінің әсерін толықтай түрде көрсете алған жоқ. Генотиптің және орта факторларының өзара әрекеттесу әсерін дат генетигі Иогансен көрсетті. Қысқа үрмебұршаққа жүргізілген тәжірибелер қатарында, ол бұл өздігінен тозаңданған өсімдіктердің әр ұрпағынан ең ауыр және ең жеңіл тұқымдарды таңдады және оларды келесі ұрпақты алу үшін өсірді. Бірнеше жылдар бойы бұл тәжірибелерді қайталай отырып, ол «ауыр» немесе «жеңіл» түрі шегінде тұқымдар орташа салмақ бойынша аз ажыратылған кезде әр-түрлі түр бойынша, яғни «ауыр» және «жеңіл» түрлер бойынша тұқымдардың орташа салмағындағы айырмашылықтар елеулі болғандығын анықтады. Бұл, белгінің фенотиптік байқалуына тұқымқуалаушылықпен қатар орта да әсер етеді деп санауға мүмкіндік береді. Бұл нәтижелер негізінде үздіксіз фенотиптік өзгергіштікті, «түрленетін генотипке әсер ететін өзгеретін орта факторларының жиынтық әсері» деп анықтауға болады. Бұл нәтижелер сонымен қатар берілген белгінің тұқымқуалау деңгейі бірінші кезекте генотиппен анықталатындығын көрсетеді. Даралық, темперамент және ақыл сияқты мұндай таза адамға тән қасиеттерге келетін болсақ, онда бар ақпараттарға қарағанда, олар тұқымқуалайтын және орта факторларына байланысты. Олар әр-түрлі индивидуумда әр-түрлі деңгейде бір-бірімен өзара әрекеттесе отырып, берілген белгінің ақырғы көрінісіне әсер етеді. Генетикалық және орта факторларындағы дәл осы айырмашылықтар адамдар арасындағы фенотиптік айырмашылықтарды құрайды. Бұл факторлардың қандай да бірі әрқашан басым болатындығы туралы нақты ақпараттар бізде жоқ, алайда орта ешқашан да генотиппен анықталған фенотипті шегінен тыс шығара алмайды (24.30 суреті).

24.30 суреті. Адамдағы бойдың фенотиптік өзгеруі. Бұл балалардың барлығы бір жасты.
24.8.4. Өзгергіштік көздері
Дискретті және үздіксіз өзгергіштік пен орта арасындағы өзара қатынас, бірдей фенотипті екі ағзаның өмір сүруін мүмкін емес қылатындығын нақты елестету қажет. Митоз кезіндегі ДНҚ репликациясы механизмі жетілуге өте жақын, себебі жыныссыз көбейетін ағзалардағы генетикалық өзгергіштік мүмкіндіктері өте аз. Сондықтан мұндай ағзалардағы кез-келген көрінетін өзгергіштік, сыртқы орта әсерімен қамтамасыз етілерлік шығар. Жынысты жолмен көбейетін ағзаларға келсек, оларда генетикалық өзгергіштіктің қалыптасуы үшін зор мүмкіншіліктер бар. Генетикалық өзгергіштіктің іс жүзінде шектелмеген көздері ретінде кроссинговер және мейоз кезіндегі хромосомалардың тәуелсіз ажырауы, сонымен қатар ұрықтандыру кезіндегі гаметалардың қосылуы қызмет атқарады. Қысқаша бұл процесстер төменде сипатталған.
24.9. Мутациялар
Мутация деп берілген ағзаның ДНҚ санының немесе құрылымының өзгерісін айтады. Мутация, мутанттық жасушадан митоз немесе мейоз нәтижесінде шығатын жасушалармен мұралана алатын генотиптің өзгеруіне әкеледі. Мутация берілген популяциядағы қандай-да бір белгінің өзгерісін тудыруы мүмкін. Егер мутация гаметаларда пайда болса, онда ол тұқымқуалау арқылы беріледі, ал сомалық жасушалардағы мутациялар митоз нәтижесінде түзілген еншілес жасушалармен ғана тұқымқуалайды. Соңғысын сомалық мутациялар деп атайды.
Хромосомалардың санының немесе құрылымының өзгеруінде көрінетін мутациялар хромосомалық мутациялар немесе хромосомалық аберрациялар (қайта құрылулар) атымен белгілі. Хромосоманың кейбір аберрацияларында хромосомалар қатты өзгереді, бұл микроскоп астынан да көрінеді. Қазіргі кезде «мутация» терминін басты түрде, гендік немесе нүктелік мутация болған кезде бір локустағы ДНҚ өзгерістерін белгілеу үшін қолданады.
Мутацияны жаңа белгінің кездейсоқ пайда болуының себебі ретіндегі юолжам алғаш 1901 жылы Oenothera lamarckiana энотерасындағы тұқымқуалауды зерттеген голланд ботанигі Гуго де Фризбен ұсынылды. 9 жыл өткен соң Т. Морган дрозофиладағы мутацияларды зерттей бастады, және көп кешікпей бүкіл әлем генетиктерінің қатысуымен оның 500-ден астам мутациялары сәйкестендірілді.
24.9.1. Мутациялар жиілігі және олардың себептері
Мутациялар кездейсоқ түрде және кенеттен пайда болады, яғни кез-келген ген кез-келген сәтте мутациялана алады. Мутациялардың пайда болу жиілігі әр-түрлі ағзаларда түрліше.
ХХ ғасырдың жиырмасыншы жылдары Г. Дж. Мёллердің жұмыстарының нәтижесінде, мутациялар жиілігін ағзаға рентген сәулелерімен әсер ете отырып, олардың күтпеген деңгейімен салыстырғанда арттыруға болатындығы орнатылды. Алда мутациялар жиілігін елеулі түрде ультракүлгін, рентген және γ- сәулелері сияқты жоғарыэлектрлік электромагниттік сәулелендіру көмегімен арттыруға болатындығы анықталды. Жоғары энергия бөлшектері – γ және β- бөлшектер, нейтрондар және ғарыштық сәулелену де – мутагенді әсерге ие, яғни мутация тудыруға қабілетті. Мутагенді әсерге алуан түрлі химиялық заттар да ие, атап айтқанда иприт, кофеин, формальдегид, колхицин, темекінің кейбір компоненттері және барлық дәрілік препараттардың, қоректік консерванттардың пестицидтердің өсу саны.
24.9.2. Хромосомалық мутациялар
Хромосомалық мутациялар хромосомалар санының немесе олардың құрылымының өзгеруінің нәтижесі бола алады. Хромосомалық мутациялардың кейбір типтері бірнеше гендердің әрекетін өзгертеді және фенотипке гендік мутацияларға қарағанда әлдеқайда тереңірек әсер етеді. Хромосомалар санының өзгеруі әдетте мейоз кезіндегі қателіктер нәтижесінде болады, бірақ бұл митоздың бұзылуында да мүмкін. Бұл өзгерістер не анэуплоидияда – жеке хромосомаларды жоғалту немесе қосуда, не эуплоидияда (полиплоидияда) – хромосоманың толық гоплоиды жиынтығында байқалады.
Анэуплоидия
Анэуплоидия, еншілес жасушаларда қосымша хромосомалардың пайда болуында (n+1), (2n+1) және т.б. немесе қандай-да бір хромосомалардың жетіспеушілігінде (n−1),(2n−1) және т.б. байқалуы мүмкін. Анэуплоидия егер мейоздың анафаза І-інде бір немесе бірнеше жұптардың гомологиялық хромосомалары ажырамаса пайда болуы мүмкін. Мұндай жағдайда жұптың екі мүшесі де бір жасуша полюсіне бағытталады, және сонда анафаза ІІ-дегі гомологиялық хромосомалардың бөлінуі құрамында нормаға қарағанда бір немесе бірнешеге көп немесе аз хромосома бар гаметалардың түзілуіне әкелуі мүмкін. Бұл құбылыс ажырамау атымен белгілі. Жетіспейтін немесе артық хромосомасы бар гамета қалыпты гаплоидты гаметамен қосылғанда, хромосомалардың тақ саны бар зигота түзіледі: бұл зигота қандай-да бір екі гомологтың орнына олар үшеу немесе тек біреу болуы мүмкін.
Хромосомалар саны диплоидтыдан аз зигота, әдетте дамымайды, бірақ артық хромосомасы бар зиготалар, әдетте дамуға қабілетті. Егер бұл жануарларда болса, онда мұндай зиготалардан көпшілік жағдайларда айқын көрінетін аномалиялы (ауытқу) дарақтар дамиды. Адамда хромосомалардың ажырамауы нәтижесінде пайда болатын, ең жиі кездесетін хромосомалық мутациялардың арасында – трисомияның бір түрі бар, Даун синдромы деп аталады (2n=47). Бұл жағдай оны 1866 жылы сипаттаған дәрігердің тегімен аталған және бұл 21 – жұп хромосомаларының ажырамауынан пайда болады. Бұл мутация 25.7.6 бөлімінде сипатталған.
Дискретті және үздіксіз өзгергіштік пен орта арасындағы өзара қатынас, бірдей фенотипті екі ағзаның өмір сүруін мүмкін емес қылатындығын нақты елестету қажет. Митоз кезіндегі ДНҚ репликациясы механизмі жетілуге өте жақын, себебі жыныссыз көбейетін ағзалардағы генетикалық өзгергіштік мүмкіндіктері өте аз. Сондықтан мұндай ағзалардағы кез-келген көрінетін өзгергіштік, сыртқы орта әсерімен қамтамасыз етілерлік шығар. Жынысты жолмен көбейетін ағзаларға келсек, оларда генетикалық өзгергіштіктің қалыптасуы үшін зор мүмкіншіліктер бар. Генетикалық өзгергіштіктің іс жүзінде шектелмеген көздері ретінде кроссинговер және мейоз кезіндегі хромосомалардың тәуелсіз ажырауы, сонымен қатар ұрықтандыру кезіндегі гаметалардың қосылуы қызмет атқарады. Қысқаша бұл процесстер төменде сипатталған.
- Кроссинговер – мейоздың профаза І-інде болатын гомологиялық хромосомалардың хроматидтері арасындағы гендермен жегілген алмасу. Мұндай алмасу нәтижесінде тіркестердің жаңа топтары және аллельдердің жаңа комбинациялары құрылады (24.3 бөлімі).
- Хромосомалардың тәуелсіз ажырауы – мейоздың метафаза І-інде веретено жазықтығында гомологиялық хромосомалардың (биваленттердің) хроматидтерінің бағдары, жұптың әр мүшесі анафаза І де орын ауыстыратын бағытты анықтайды. Хроматидтердің бұл бағдары кездейсоқ сипатқа ие. Метафаза ІІ кезінде хроматидтер бағдары қайтадан кездейсоқ түрде болады және анафаза ІІ кезінде әр хромосоманың екі қарама-қарсы полюстердің қайсысына бағытталатынын анықтайды. Хромосомалардың кездейсоқ бағдары және кезекті тәуелсіз ажырауы (сегрегация), гаметалардағы хромосомалардың әр-түрлі комбинацияларының үлкен санын мүмкін етеді. Бұл санды есептеуге болады (24.2.1 бөлімі).
- Кездейсоқ ұрықтану – жынысты көбеюде аталық және аналық гаметалардың қосылуы толықтай кездейсоқ болуы (қандай жағдай болса да теориялық түрде) нәтижесінде туатын өзгергіштіктің үшінші көзі. Кез-келген аталық гамета кез-келген аналық гаметамен қосылуға потенциалды мүмкіндікке ие.
24.9. Мутациялар
Мутация деп берілген ағзаның ДНҚ санының немесе құрылымының өзгерісін айтады. Мутация, мутанттық жасушадан митоз немесе мейоз нәтижесінде шығатын жасушалармен мұралана алатын генотиптің өзгеруіне әкеледі. Мутация берілген популяциядағы қандай-да бір белгінің өзгерісін тудыруы мүмкін. Егер мутация гаметаларда пайда болса, онда ол тұқымқуалау арқылы беріледі, ал сомалық жасушалардағы мутациялар митоз нәтижесінде түзілген еншілес жасушалармен ғана тұқымқуалайды. Соңғысын сомалық мутациялар деп атайды.
Хромосомалардың санының немесе құрылымының өзгеруінде көрінетін мутациялар хромосомалық мутациялар немесе хромосомалық аберрациялар (қайта құрылулар) атымен белгілі. Хромосоманың кейбір аберрацияларында хромосомалар қатты өзгереді, бұл микроскоп астынан да көрінеді. Қазіргі кезде «мутация» терминін басты түрде, гендік немесе нүктелік мутация болған кезде бір локустағы ДНҚ өзгерістерін белгілеу үшін қолданады.
Мутацияны жаңа белгінің кездейсоқ пайда болуының себебі ретіндегі юолжам алғаш 1901 жылы Oenothera lamarckiana энотерасындағы тұқымқуалауды зерттеген голланд ботанигі Гуго де Фризбен ұсынылды. 9 жыл өткен соң Т. Морган дрозофиладағы мутацияларды зерттей бастады, және көп кешікпей бүкіл әлем генетиктерінің қатысуымен оның 500-ден астам мутациялары сәйкестендірілді.
24.9.1. Мутациялар жиілігі және олардың себептері
Мутациялар кездейсоқ түрде және кенеттен пайда болады, яғни кез-келген ген кез-келген сәтте мутациялана алады. Мутациялардың пайда болу жиілігі әр-түрлі ағзаларда түрліше.
ХХ ғасырдың жиырмасыншы жылдары Г. Дж. Мёллердің жұмыстарының нәтижесінде, мутациялар жиілігін ағзаға рентген сәулелерімен әсер ете отырып, олардың күтпеген деңгейімен салыстырғанда арттыруға болатындығы орнатылды. Алда мутациялар жиілігін елеулі түрде ультракүлгін, рентген және γ- сәулелері сияқты жоғарыэлектрлік электромагниттік сәулелендіру көмегімен арттыруға болатындығы анықталды. Жоғары энергия бөлшектері – γ және β- бөлшектер, нейтрондар және ғарыштық сәулелену де – мутагенді әсерге ие, яғни мутация тудыруға қабілетті. Мутагенді әсерге алуан түрлі химиялық заттар да ие, атап айтқанда иприт, кофеин, формальдегид, колхицин, темекінің кейбір компоненттері және барлық дәрілік препараттардың, қоректік консерванттардың пестицидтердің өсу саны.
24.9.2. Хромосомалық мутациялар
Хромосомалық мутациялар хромосомалар санының немесе олардың құрылымының өзгеруінің нәтижесі бола алады. Хромосомалық мутациялардың кейбір типтері бірнеше гендердің әрекетін өзгертеді және фенотипке гендік мутацияларға қарағанда әлдеқайда тереңірек әсер етеді. Хромосомалар санының өзгеруі әдетте мейоз кезіндегі қателіктер нәтижесінде болады, бірақ бұл митоздың бұзылуында да мүмкін. Бұл өзгерістер не анэуплоидияда – жеке хромосомаларды жоғалту немесе қосуда, не эуплоидияда (полиплоидияда) – хромосоманың толық гоплоиды жиынтығында байқалады.
Анэуплоидия
Анэуплоидия, еншілес жасушаларда қосымша хромосомалардың пайда болуында (n+1), (2n+1) және т.б. немесе қандай-да бір хромосомалардың жетіспеушілігінде (n−1),(2n−1) және т.б. байқалуы мүмкін. Анэуплоидия егер мейоздың анафаза І-інде бір немесе бірнеше жұптардың гомологиялық хромосомалары ажырамаса пайда болуы мүмкін. Мұндай жағдайда жұптың екі мүшесі де бір жасуша полюсіне бағытталады, және сонда анафаза ІІ-дегі гомологиялық хромосомалардың бөлінуі құрамында нормаға қарағанда бір немесе бірнешеге көп немесе аз хромосома бар гаметалардың түзілуіне әкелуі мүмкін. Бұл құбылыс ажырамау атымен белгілі. Жетіспейтін немесе артық хромосомасы бар гамета қалыпты гаплоидты гаметамен қосылғанда, хромосомалардың тақ саны бар зигота түзіледі: бұл зигота қандай-да бір екі гомологтың орнына олар үшеу немесе тек біреу болуы мүмкін.
Хромосомалар саны диплоидтыдан аз зигота, әдетте дамымайды, бірақ артық хромосомасы бар зиготалар, әдетте дамуға қабілетті. Егер бұл жануарларда болса, онда мұндай зиготалардан көпшілік жағдайларда айқын көрінетін аномалиялы (ауытқу) дарақтар дамиды. Адамда хромосомалардың ажырамауы нәтижесінде пайда болатын, ең жиі кездесетін хромосомалық мутациялардың арасында – трисомияның бір түрі бар, Даун синдромы деп аталады (2n=47). Бұл жағдай оны 1866 жылы сипаттаған дәрігердің тегімен аталған және бұл 21 – жұп хромосомаларының ажырамауынан пайда болады. Бұл мутация 25.7.6 бөлімінде сипатталған.
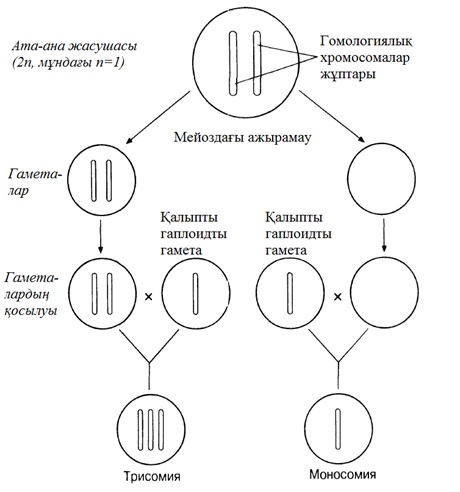
24.31 суреті. Гаметалар түзілуі кезіндегі хромосомалардың ажырамауы және аномалиялы гаметалардың қалыпты гаплоидты жасушалармен қосылуы. Бұл полисомияның әр-түрлі типтеріне әкеледі, оларда хромосомалар саны 2n+1 (трисомия), 2n+2 (тетрасомия) 2n+3 (пентасомия) және т.б.; немесе 2n-1 (моносомия)- ға тең болуы мүмкін, - қалыпты түрде аждырамаған гомологиялық хромосомалардың санына байланысты (сонымен қатар 25.34 суретін қараңыз).
Сонымен қатар аталық және аналық жыныс хромосомаларының ажырамауы да болуы мүмкін, бұл, екінші реттік жыныстық белгілерге және ұрықтылыққа, ал кейде ақыл-ой қабілетіне де әсер ететін анэуплоидияға әкеледі (25.7.7 және 25.7.8 бөлімдері).
Эуплоидия (полиплоидия)
Өздерінің гаплоидты санына еселі хромосомалардың артқан саны бар гаметаларды және сомалық жасушаларды полиплоидты деп атайды. Три − , тетра – және т.б. қосымшалар, хромосомалар санының, яғни плоидтылық деңгейінің қанша есе артқанын көрсетеді: 3n – триплоид, 4n – тетраплоид, 5n – пентаплоид және т.б. Өсімдіктерде полиплоидия жануарларға қарағанда жиірек кездеседі. Мысалы, жабықтұқымдылардың 300 000 белгілі түрлерінің шамамен жартысы – полиплоидты. Жануарлардағы полиплоидияның салыстырмалы түрде сиректігі, хромосомалар санының артуының, мейоздағы гаметалар түзілуіндегі қателіктер мүмкіндігін елеулі түрде көтеретіндігімен түсіндіріледі. Өсімдіктерге келер болсақ, онда олардың көпшілігі вегетативті көбеюге қабілетті, сондықтан олар полиплоидты жағдайда да қалпына келе алады. Полиплоидты ағзалар жиі жайлы белгілерге – үлкенірек көлемге, шыдамдылыққа, ауруларға төзімділікке ие. Олардың бұл қасиетін будандық қуат деп атайды (27.4.2 бөлімі). Мәдени өсімдіктердің көпшілігі – полиплоидтар, ірі тұқымдар түзеді, мүшелерді, гүлдерді немесе жапырақтарды қорға жинайды.
Полиплоидияның екі түрі болады – аутополиплоидия және аллополиплоидия.
АУТОПОЛИПЛОИДИЯ. Аутополиплоидия, берілген түрде хромосома жиынтықтары санының артуы нәтижесінде табиғи және жасанды жолмен пайда болуы мүмкін. Мысалы, егер хромосомалар репликациясы болса (интерфазада) және хроматидтер қалыпты ажыраса (анафазада), бірақ цитоплазма бөлінбесе, онда үлкен ядролы тетраплоидты (4n) жасуша түзіледі. Мұндай жасуша содан соң бөлінеді және тетраплоидты еншілес жасушалар береді. Бұл жасушалардағы цитоплазма мөлшері, тұрақты ядро – плазмалық қатынас сақталуы үшін өседі, және бұл өсімдіктің немесе оның қандай да бір бөлігінің жалпы мөлшерінің артуына әкеледі. Аутополиплоидияны лапыздың (Colchicum) түйнекбаданасынан бөлінетін колхицин алкалоиды көмегімен тудыруға болады. Колхицин шамамен 0,01% концентрацияда микротүтікшелерді бұза отырып веретено бөлінуінің түзілуін басады, сондықтан да хромосомалар анафаза кезінде ажырамайды. Колхицинді және оған жақын заттарды, темекі, томат және қант қызылшасы сияқты дақылдардың бағалы сұрыптарын жасау үшін қолданған. Аутополиплоидтардың құрамында хромосомалар жиынтығының жұп саны болса олар диплоидтар сияқты жемісті болуы мүмкін.
Жануарларда кейде полиплоидияның түріөзгерген формасы бақыланады. Онда жеке полиплоидты жасушалар және ұлпалар түзіледі. Бұл эндомитоз деп аталатын процесс хромосомалар жасушасының бөлінуімен жүрмейтін хромосомалар репликациясында болып табылады. Дрозофиланың сілекей безінің жасушаларындағы алып хромосомалар және адам бауырындағы тетраплоидты жасушалар эндомитоз нәтижесінде туады.
АЛЛОПОЛИПЛОИДИЯ. Аллополиплоидия стерильді (ұрықсыз) будандағы хромосомалар саны екіеселенген кезде пайда болады, мұның нәтижесінде ол фертильді (ұрықты) болады. Әр-түрлі түрлер арасындағы шағылысудан пайда болатын F1 дарақтары, әдетте стерильді, себебі олардың хромосомалары мейоз кезінде гомологиялық жұптар түзе алмайды. Бұл құбылысты будандық ұрықсыздық деп атайды. Алайды егер хромосомалар саны олардың бастапқы гаплоидты санына еселі болса, мысалы 2(n1+n2), 3(n1+n2) және т.б. бұл жерде n1 және n2 – ата-аналық түрлердегі хромосомалардың гаплоидты саны, онда дәл осындай полиплоидтармен шағылыстырғанда фертильді дарақтар беретін, бірақ ата-аналық түрдің бірімен шағылыстырғанда стерильді болатын жаңа түр пайда болады.
Аллополиплоидты түрлердің көбіне ата-аналық түрлердің диплоидтық санының жиынтығын беретін хромосомалардың диплоиды саны тән. Мысалы, Spartina maritime (stricta) (2n=60) –ні Spartina alterniflora (2n=62) –мен шағылыстыру нәтижесіңнде алынған, фертильді буданды аллополиплоид – Spartina Anglica (2n=122). Бұл екі түр арасындағы шағылысудан алынған F1 буданы стерильді және Spartina townsendil (2n=62) атауын алды. Аллополиплоидты өсімдіктердің көпшілігі екі ата-аналық түрден де өзінің белгілері бойынша ажыратылады; олардың арасында көбі адаммен өсірілетін тым бағалы дақылдардан. Мысалы, ұн алынатын бидайдың түрі Triticum aestivum (2n=42), 5000 жыл бойына шағылыстыру және іріктеу жолымен өсірілді. Бір дәнекті бидайды (2n=14) айқындалмаған диплоидпен (2n=14) шағылыстыру нәтижесінде бидайдың жаңа түрі – эммер (2n=28) алынды. Эммер бидайын тағы да бір жабайы өсетін түрмен (2n=14) шағылыстырды, нәтижесінде бастапқы бір дәнекті бидайдың гексаплоиды (6n) болып табылатын Triticum aestivum (2n=42) формасы туды. Түраралық будандастырудың басқа мысалы – шомырды қырыққабатпен шағылыстыру – 27.9 бөлімінде сипатталатын болады.
Жануарларда аллополиплоидия белгісіз дерлік, себебі оларда түраралық будандастыру сирек болады. Полиплоидия нәтижесінде генофонд жаңа гендер алмайды (27.4.2 бөлімі), бірақ гендердің жаңа комбинациялары пайда болады.
Хромосомалардың құрылымдық өзгерістері
Мейоздың профаза І кезіндегі кроссинговерде гомологиялық хромосомалар арасында генетикалық материалмен реципрокты алмасу болады. Бұл тіркестің ата-аналық топтарында аллельдердің реттілігінің өзгеруіне әкеледі, нәтижесінде рекомбинанттар пайда болады, бірақ қандай-да бір гендік локустардың шығынынсыз. Ұқсас әсерлер инверсия және транслокация сияқты хромосомалық қайта құрылуларда пайда болады. Басқа типтердің қайта құрылуларында – делегация және дупликацияларда – хромосомалардағы гендік локустардың саны өзгереді, және бұл фенотиптерге терең әсер етуі мүмкін. Инверсиямен, делегациямен, дупликациямен, ал кейбір жағдайларда транслокациямен байланысты хромосомалардағы құрылымдық өзгерістерді, мейоздың профаза І-індегі гомологиялық хромосомалар конъюгацияға кіріскен кезде микроскоп астында бақылауға болады. Гомологиялық хромосомалар синапс түзеді (конъюгацияланады; 23.3 бөлімі), ал қайта құрылулар тиген аймақтарда, гомологиялық хромосомалардың біреуі ілмек түзеді немесе ширатылады. Хромосомалардың қайсысы ілмек түзетіні және оның гендері қалай орналасатыны қайта құрылудың типіне байланысты.
Эуплоидия (полиплоидия)
Өздерінің гаплоидты санына еселі хромосомалардың артқан саны бар гаметаларды және сомалық жасушаларды полиплоидты деп атайды. Три − , тетра – және т.б. қосымшалар, хромосомалар санының, яғни плоидтылық деңгейінің қанша есе артқанын көрсетеді: 3n – триплоид, 4n – тетраплоид, 5n – пентаплоид және т.б. Өсімдіктерде полиплоидия жануарларға қарағанда жиірек кездеседі. Мысалы, жабықтұқымдылардың 300 000 белгілі түрлерінің шамамен жартысы – полиплоидты. Жануарлардағы полиплоидияның салыстырмалы түрде сиректігі, хромосомалар санының артуының, мейоздағы гаметалар түзілуіндегі қателіктер мүмкіндігін елеулі түрде көтеретіндігімен түсіндіріледі. Өсімдіктерге келер болсақ, онда олардың көпшілігі вегетативті көбеюге қабілетті, сондықтан олар полиплоидты жағдайда да қалпына келе алады. Полиплоидты ағзалар жиі жайлы белгілерге – үлкенірек көлемге, шыдамдылыққа, ауруларға төзімділікке ие. Олардың бұл қасиетін будандық қуат деп атайды (27.4.2 бөлімі). Мәдени өсімдіктердің көпшілігі – полиплоидтар, ірі тұқымдар түзеді, мүшелерді, гүлдерді немесе жапырақтарды қорға жинайды.
Полиплоидияның екі түрі болады – аутополиплоидия және аллополиплоидия.
АУТОПОЛИПЛОИДИЯ. Аутополиплоидия, берілген түрде хромосома жиынтықтары санының артуы нәтижесінде табиғи және жасанды жолмен пайда болуы мүмкін. Мысалы, егер хромосомалар репликациясы болса (интерфазада) және хроматидтер қалыпты ажыраса (анафазада), бірақ цитоплазма бөлінбесе, онда үлкен ядролы тетраплоидты (4n) жасуша түзіледі. Мұндай жасуша содан соң бөлінеді және тетраплоидты еншілес жасушалар береді. Бұл жасушалардағы цитоплазма мөлшері, тұрақты ядро – плазмалық қатынас сақталуы үшін өседі, және бұл өсімдіктің немесе оның қандай да бір бөлігінің жалпы мөлшерінің артуына әкеледі. Аутополиплоидияны лапыздың (Colchicum) түйнекбаданасынан бөлінетін колхицин алкалоиды көмегімен тудыруға болады. Колхицин шамамен 0,01% концентрацияда микротүтікшелерді бұза отырып веретено бөлінуінің түзілуін басады, сондықтан да хромосомалар анафаза кезінде ажырамайды. Колхицинді және оған жақын заттарды, темекі, томат және қант қызылшасы сияқты дақылдардың бағалы сұрыптарын жасау үшін қолданған. Аутополиплоидтардың құрамында хромосомалар жиынтығының жұп саны болса олар диплоидтар сияқты жемісті болуы мүмкін.
Жануарларда кейде полиплоидияның түріөзгерген формасы бақыланады. Онда жеке полиплоидты жасушалар және ұлпалар түзіледі. Бұл эндомитоз деп аталатын процесс хромосомалар жасушасының бөлінуімен жүрмейтін хромосомалар репликациясында болып табылады. Дрозофиланың сілекей безінің жасушаларындағы алып хромосомалар және адам бауырындағы тетраплоидты жасушалар эндомитоз нәтижесінде туады.
АЛЛОПОЛИПЛОИДИЯ. Аллополиплоидия стерильді (ұрықсыз) будандағы хромосомалар саны екіеселенген кезде пайда болады, мұның нәтижесінде ол фертильді (ұрықты) болады. Әр-түрлі түрлер арасындағы шағылысудан пайда болатын F1 дарақтары, әдетте стерильді, себебі олардың хромосомалары мейоз кезінде гомологиялық жұптар түзе алмайды. Бұл құбылысты будандық ұрықсыздық деп атайды. Алайды егер хромосомалар саны олардың бастапқы гаплоидты санына еселі болса, мысалы 2(n1+n2), 3(n1+n2) және т.б. бұл жерде n1 және n2 – ата-аналық түрлердегі хромосомалардың гаплоидты саны, онда дәл осындай полиплоидтармен шағылыстырғанда фертильді дарақтар беретін, бірақ ата-аналық түрдің бірімен шағылыстырғанда стерильді болатын жаңа түр пайда болады.
Аллополиплоидты түрлердің көбіне ата-аналық түрлердің диплоидтық санының жиынтығын беретін хромосомалардың диплоиды саны тән. Мысалы, Spartina maritime (stricta) (2n=60) –ні Spartina alterniflora (2n=62) –мен шағылыстыру нәтижесіңнде алынған, фертильді буданды аллополиплоид – Spartina Anglica (2n=122). Бұл екі түр арасындағы шағылысудан алынған F1 буданы стерильді және Spartina townsendil (2n=62) атауын алды. Аллополиплоидты өсімдіктердің көпшілігі екі ата-аналық түрден де өзінің белгілері бойынша ажыратылады; олардың арасында көбі адаммен өсірілетін тым бағалы дақылдардан. Мысалы, ұн алынатын бидайдың түрі Triticum aestivum (2n=42), 5000 жыл бойына шағылыстыру және іріктеу жолымен өсірілді. Бір дәнекті бидайды (2n=14) айқындалмаған диплоидпен (2n=14) шағылыстыру нәтижесінде бидайдың жаңа түрі – эммер (2n=28) алынды. Эммер бидайын тағы да бір жабайы өсетін түрмен (2n=14) шағылыстырды, нәтижесінде бастапқы бір дәнекті бидайдың гексаплоиды (6n) болып табылатын Triticum aestivum (2n=42) формасы туды. Түраралық будандастырудың басқа мысалы – шомырды қырыққабатпен шағылыстыру – 27.9 бөлімінде сипатталатын болады.
Жануарларда аллополиплоидия белгісіз дерлік, себебі оларда түраралық будандастыру сирек болады. Полиплоидия нәтижесінде генофонд жаңа гендер алмайды (27.4.2 бөлімі), бірақ гендердің жаңа комбинациялары пайда болады.
Хромосомалардың құрылымдық өзгерістері
Мейоздың профаза І кезіндегі кроссинговерде гомологиялық хромосомалар арасында генетикалық материалмен реципрокты алмасу болады. Бұл тіркестің ата-аналық топтарында аллельдердің реттілігінің өзгеруіне әкеледі, нәтижесінде рекомбинанттар пайда болады, бірақ қандай-да бір гендік локустардың шығынынсыз. Ұқсас әсерлер инверсия және транслокация сияқты хромосомалық қайта құрылуларда пайда болады. Басқа типтердің қайта құрылуларында – делегация және дупликацияларда – хромосомалардағы гендік локустардың саны өзгереді, және бұл фенотиптерге терең әсер етуі мүмкін. Инверсиямен, делегациямен, дупликациямен, ал кейбір жағдайларда транслокациямен байланысты хромосомалардағы құрылымдық өзгерістерді, мейоздың профаза І-індегі гомологиялық хромосомалар конъюгацияға кіріскен кезде микроскоп астында бақылауға болады. Гомологиялық хромосомалар синапс түзеді (конъюгацияланады; 23.3 бөлімі), ал қайта құрылулар тиген аймақтарда, гомологиялық хромосомалардың біреуі ілмек түзеді немесе ширатылады. Хромосомалардың қайсысы ілмек түзетіні және оның гендері қалай орналасатыны қайта құрылудың типіне байланысты.
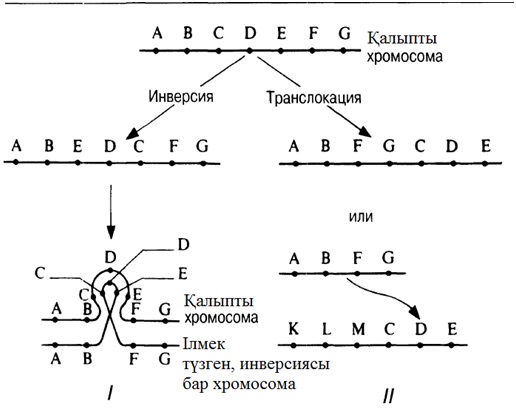
24.32 суреті. Инверсияның және транслокацияның сызбалық суреттері және олардың А – G гендерінің орналасуына әсері. І. Инверсиямен тудырылған мейоз профазасындағы ілмектің түзілуі. ІІ. C, D және E гендерін алып жүретін хромосома аймағы, үзілді және K,L және M гендерін алып жүретін хромосомаға қосылды.
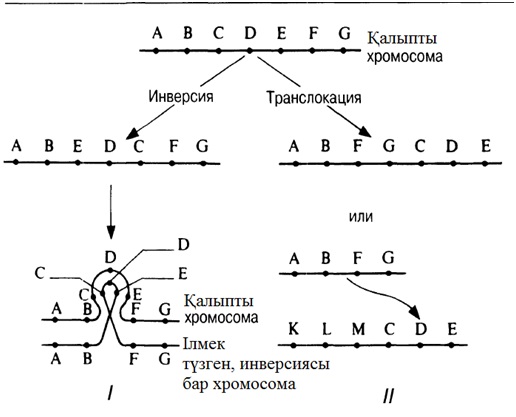
24.33 суреті. Делеция және дупликацияның сызбалық суреті және олардың А – G гендерінің орналасуына әсері. Екі жағдайда да ілмектер түзіледі.
Инверсия хромосоманың 180°-қа айналатын, содан соң қайта бұрынғы орнына келетін андай да бір аймағының кесілуі нәтижесінде пайда болады. Сонымен бірге генотиптің ешқандай да өзгерісі болмайды, бірақ фенотиптік өзгерістер болуы мүмкін (24.32 суреті). Бұл берілген хромосомадағы гендер реттілігінің мәні – геннің орналасу әсері туралы айғақтайды.
Транслокация кезінде хромосомалардың біреуінен аймақ бөлінеді, ол не сол хромосоманың басқа шетіне, не басқа, гомологиялық емес хромосоманың екі шетінің біріне қосылады (24.31 суреті). Және мұндай жағдайда фенотипте орын әсері байқалуы мүмкін. Гомологиялық емес хромосомалар арасындағы реципрокты транслокация, гомологиялық хромосомалардың екі жаңа жұбының түзілуіне әкелуі мүмкін. Даун синдромында хромосомалардың қалыпты диплоиды саны сақталған жағдайларда, бұның себебі ретінде қосымша хромосома 21-дің ірірек хромосомалардың біріне, әдетте 15-інші хромосомаға транслокациясы қызмет атқарады.
Хромосомалық мутацияның ең қарапайым формасы – делеция, немесе тапшылық, яғни хромосомамен қандай да бір аймақты жоғалту, бұның нәтижесінде хромосомада кейбір гендер жетіспейді (24.33 суреті). Делеция екі гомологиялық хромосоманың бірінде болуы мүмкін; мұндай жағдайда басқа қалыпты хромосомада орналасқан аллельдер олар рецессивті болса да экспрессияланады. Егер делеция гомологиялық хромосомалардағы бірдей локустарға тисе, бұл әдетте өліммен аяқталады.
Кейде хромосоманың қандай-да бір аймағы екі еселенеді, сондықтан да дупликация – бұл аймақта орналасқан гендер жиынтығының қайталануы туады. Қосымша жиынтық сол хромосомаға қосылған және оның шеттерінің бірінде орналасқан болуы мүмкін немесе басқа қандай-да бір хромосомаға қосылған болуы мүмкін.
24.9.3. Гендік мутациялар
Гендік мутация дегеніміз не?
Фенотиптің хромосомалық аберрациялармен (ауытқулармен) байланыссыз (микроскоптық зерттеулердің мәліметтерімен айқындалатын) кездейсоқ кенет өзгерістерін, жеке гендер құрылымындағы өзгерістермен ғана түсірдіруге болады. Гендік немесе нүктелік мутация (қанша дегенмен ол белгілі гендік локусқа жататындықтан) – хромосоманың белгілі аймағындағы ДНҚ молекуласының нуклеотидтік реттілігі өзгерісінің нәтижесі. Бір гендегі мұндай негіздердің реттілігі мРНҚ транскрипциясында қалпына келтіріледі және рибосомалардағы трасляция нәтижесінде түзілетін полипептидтік тізбектегі аминқышқылдар реттілігінің өзгеруіне әкелуі мүмкін.
Гендік мутациялардың түрлері
Гендік мутацияның қосылумен, түсумен немесе берілген гендегі негіздерді ауыстырумен байланысты алуан түрлі түрлері бар. Олар дупликация, кірістірме, делеция, инверсия немесе негіздерді ауыстыру формасында байқалады. Барлық жағдайларда олар нуклеотидті реттіліктің өзгеруіне, жиі өзгертілген полипептидтің түзілуіне де әкеледі. Мысалы, делеция санау жақтаушасының жылжуын тудырады, оның салдарлары 23.7 бөлімінде сипатталған.
Гендік мутациялардың салдарлары
Гаметалар қалыптасуы кезінде пайда болатын гендік мутациялар, ұрпақтардың барлық жасушаларына беріледі және түрдің алдағы тағдырына әсер етуі мүмкін. Ағзада болатын сомалық гендік мутациялар, митоз нәтижесінде мутанттық жасушалардан түзілетін жасушалармен ғана тұқымқуалайды. Олар өздері пайда болған ағзаға әсер етуі мүмкін, бірақ берілген дарақтың өлімінен генофондтан популяциялар жойылады. Сомалық мутациялар өте жиі пайда болатындығы және әдетте білінбейтін болып қалатындығы белгілі, бірақ кейбір жағдайларда сонымен бірге бой және бөліну бойынша жоғары жылдамдықты жасушалар түзіледі. Бұл жасушалар ісіктерге бастама беруі мүмкін – не басқа ұлпаларға әсер етпейтін қатерсіз ісіктер, не басқа ұлпаларда паразиттейтін қатерлі ісіктер (рак, меланомалар, саркомалар).
Гендік мутациялардың әсерлері шектен тыс алуан түрлі. Ұсақ гендік мутациялардың көп бөлігі фенотиптік байқалмайды, қанша дегенмен олар рецессивті, алайда генетикалық кодтағы бір ғана негіздің өзгеруі фенотипке терең әсер еткен жағдайлар қатары белгілі. Бір мысал ретінде орақ жасушалы анемия – адамда гемоглобин синтезіне жауапты гендердің бірінде негіздің ауысуымен туатын ауру қызмет атқарады. Бұл ауру және оның себептері туралы нақтырақ 25.7.2 бөлімінде сипатталған.
24.9.4. Мутацияның мәні
Хромосомалық және гендік мутациялар ағзаға алуан түрлі әсер етеді. Көптеген жағдайларда бұл мутациялар өлімге душар етеді және дамуды бұзады; адамда, мысалы, 20% жүктілік 12 аптаға дейінгі мерзімде өздігінен болған түсікпен аяқталады. Сонымен бірге, аборт жасаушылардың жартысында хромосомалық аномалиялар анықталады. Егер де кейбір хромосомалық мутациялар нәтижесінде анықталған гендер жақын орналасса, онда кейде олардың ортақ әсері қандай-да бір қолайлы белгінің пайда болуына әкеледі. Сонымен қатар, кейбір гендердің жақындасуы олардың кроссинговер нәтижесінде бөліну мүмкіндігін азайтады, ал қолайлы гендер жағдайында бұл артықшылықтар тудырады.
Гендік мутация, белгілі локуста бірнеше аллельдер болуына әкелуі мүмкін. Бұл берілген популяцияның гетерозиготалығын көбейтеді, оның генофондын алуан түрлірек етеді және популяция ішілік өзгергіштіктің күшеюіне әкеледі. Кроссинговердің, тәуелсіз ажыраудың, кездейсоқ ұрықтанудың және мутацияның нәтижесі ретіндегі гендердің тасымалы үздіксіз өзгергіштікті жоғарылатуы мүмкін, бірақ оның эволюциялық рөлі жиі өткінші болады, себебі мұндағы пайда болатын өзгерістер тез қалпына келуі мүмкін. Гендік мутацияларға келер болсақ, онда олардың кейбірі дискретті өзгергіштікті жоғарылатуы мүмкін, және бұл популяцияға тереңірек әсер етуі мүмкін. Гендік мутациялардың көпшілігі, көптеген ұрпақтар бойы іріктеуге сәтті шыдаған «қалыпты» аллельге қарағанда, рецессивті болып табылады.
Рецессивті бола отырып мутанттық аллельдер оларға кездесу мүмкін болмайынша, яғни гомозиготалы жағдайда болып, фенотипте байқалмайынша, көптеген ұрпақтар бойына популяцияда қалуы мүмкін. Уақыт өте келе фенотипте тез байқалатын доминантты мутанттық аллельдер де пайда болуы мүмкін (27.5 бөлім, Biston betularia).
Бұл тарауда тұжырымдалған ақпараттар популяция ішілік өзгергіштіктің шығу тегі және белгілердің тұқымқуалау механизмдері туралы түсінік береді, бірақ ол 2-тарауда сипатталған тірі ағзалардың таңғаларлық әртүрлілігінің қалайша пайда болғандығын түсіндірмейді. Бұл сұраққа жауап беру мүмкіндіктері келесі үш тараудың мазмұнын қамтиды.
Транслокация кезінде хромосомалардың біреуінен аймақ бөлінеді, ол не сол хромосоманың басқа шетіне, не басқа, гомологиялық емес хромосоманың екі шетінің біріне қосылады (24.31 суреті). Және мұндай жағдайда фенотипте орын әсері байқалуы мүмкін. Гомологиялық емес хромосомалар арасындағы реципрокты транслокация, гомологиялық хромосомалардың екі жаңа жұбының түзілуіне әкелуі мүмкін. Даун синдромында хромосомалардың қалыпты диплоиды саны сақталған жағдайларда, бұның себебі ретінде қосымша хромосома 21-дің ірірек хромосомалардың біріне, әдетте 15-інші хромосомаға транслокациясы қызмет атқарады.
Хромосомалық мутацияның ең қарапайым формасы – делеция, немесе тапшылық, яғни хромосомамен қандай да бір аймақты жоғалту, бұның нәтижесінде хромосомада кейбір гендер жетіспейді (24.33 суреті). Делеция екі гомологиялық хромосоманың бірінде болуы мүмкін; мұндай жағдайда басқа қалыпты хромосомада орналасқан аллельдер олар рецессивті болса да экспрессияланады. Егер делеция гомологиялық хромосомалардағы бірдей локустарға тисе, бұл әдетте өліммен аяқталады.
Кейде хромосоманың қандай-да бір аймағы екі еселенеді, сондықтан да дупликация – бұл аймақта орналасқан гендер жиынтығының қайталануы туады. Қосымша жиынтық сол хромосомаға қосылған және оның шеттерінің бірінде орналасқан болуы мүмкін немесе басқа қандай-да бір хромосомаға қосылған болуы мүмкін.
24.9.3. Гендік мутациялар
Гендік мутация дегеніміз не?
Фенотиптің хромосомалық аберрациялармен (ауытқулармен) байланыссыз (микроскоптық зерттеулердің мәліметтерімен айқындалатын) кездейсоқ кенет өзгерістерін, жеке гендер құрылымындағы өзгерістермен ғана түсірдіруге болады. Гендік немесе нүктелік мутация (қанша дегенмен ол белгілі гендік локусқа жататындықтан) – хромосоманың белгілі аймағындағы ДНҚ молекуласының нуклеотидтік реттілігі өзгерісінің нәтижесі. Бір гендегі мұндай негіздердің реттілігі мРНҚ транскрипциясында қалпына келтіріледі және рибосомалардағы трасляция нәтижесінде түзілетін полипептидтік тізбектегі аминқышқылдар реттілігінің өзгеруіне әкелуі мүмкін.
Гендік мутациялардың түрлері
Гендік мутацияның қосылумен, түсумен немесе берілген гендегі негіздерді ауыстырумен байланысты алуан түрлі түрлері бар. Олар дупликация, кірістірме, делеция, инверсия немесе негіздерді ауыстыру формасында байқалады. Барлық жағдайларда олар нуклеотидті реттіліктің өзгеруіне, жиі өзгертілген полипептидтің түзілуіне де әкеледі. Мысалы, делеция санау жақтаушасының жылжуын тудырады, оның салдарлары 23.7 бөлімінде сипатталған.
Гендік мутациялардың салдарлары
Гаметалар қалыптасуы кезінде пайда болатын гендік мутациялар, ұрпақтардың барлық жасушаларына беріледі және түрдің алдағы тағдырына әсер етуі мүмкін. Ағзада болатын сомалық гендік мутациялар, митоз нәтижесінде мутанттық жасушалардан түзілетін жасушалармен ғана тұқымқуалайды. Олар өздері пайда болған ағзаға әсер етуі мүмкін, бірақ берілген дарақтың өлімінен генофондтан популяциялар жойылады. Сомалық мутациялар өте жиі пайда болатындығы және әдетте білінбейтін болып қалатындығы белгілі, бірақ кейбір жағдайларда сонымен бірге бой және бөліну бойынша жоғары жылдамдықты жасушалар түзіледі. Бұл жасушалар ісіктерге бастама беруі мүмкін – не басқа ұлпаларға әсер етпейтін қатерсіз ісіктер, не басқа ұлпаларда паразиттейтін қатерлі ісіктер (рак, меланомалар, саркомалар).
Гендік мутациялардың әсерлері шектен тыс алуан түрлі. Ұсақ гендік мутациялардың көп бөлігі фенотиптік байқалмайды, қанша дегенмен олар рецессивті, алайда генетикалық кодтағы бір ғана негіздің өзгеруі фенотипке терең әсер еткен жағдайлар қатары белгілі. Бір мысал ретінде орақ жасушалы анемия – адамда гемоглобин синтезіне жауапты гендердің бірінде негіздің ауысуымен туатын ауру қызмет атқарады. Бұл ауру және оның себептері туралы нақтырақ 25.7.2 бөлімінде сипатталған.
24.9.4. Мутацияның мәні
Хромосомалық және гендік мутациялар ағзаға алуан түрлі әсер етеді. Көптеген жағдайларда бұл мутациялар өлімге душар етеді және дамуды бұзады; адамда, мысалы, 20% жүктілік 12 аптаға дейінгі мерзімде өздігінен болған түсікпен аяқталады. Сонымен бірге, аборт жасаушылардың жартысында хромосомалық аномалиялар анықталады. Егер де кейбір хромосомалық мутациялар нәтижесінде анықталған гендер жақын орналасса, онда кейде олардың ортақ әсері қандай-да бір қолайлы белгінің пайда болуына әкеледі. Сонымен қатар, кейбір гендердің жақындасуы олардың кроссинговер нәтижесінде бөліну мүмкіндігін азайтады, ал қолайлы гендер жағдайында бұл артықшылықтар тудырады.
Гендік мутация, белгілі локуста бірнеше аллельдер болуына әкелуі мүмкін. Бұл берілген популяцияның гетерозиготалығын көбейтеді, оның генофондын алуан түрлірек етеді және популяция ішілік өзгергіштіктің күшеюіне әкеледі. Кроссинговердің, тәуелсіз ажыраудың, кездейсоқ ұрықтанудың және мутацияның нәтижесі ретіндегі гендердің тасымалы үздіксіз өзгергіштікті жоғарылатуы мүмкін, бірақ оның эволюциялық рөлі жиі өткінші болады, себебі мұндағы пайда болатын өзгерістер тез қалпына келуі мүмкін. Гендік мутацияларға келер болсақ, онда олардың кейбірі дискретті өзгергіштікті жоғарылатуы мүмкін, және бұл популяцияға тереңірек әсер етуі мүмкін. Гендік мутациялардың көпшілігі, көптеген ұрпақтар бойы іріктеуге сәтті шыдаған «қалыпты» аллельге қарағанда, рецессивті болып табылады.
Рецессивті бола отырып мутанттық аллельдер оларға кездесу мүмкін болмайынша, яғни гомозиготалы жағдайда болып, фенотипте байқалмайынша, көптеген ұрпақтар бойына популяцияда қалуы мүмкін. Уақыт өте келе фенотипте тез байқалатын доминантты мутанттық аллельдер де пайда болуы мүмкін (27.5 бөлім, Biston betularia).
Бұл тарауда тұжырымдалған ақпараттар популяция ішілік өзгергіштіктің шығу тегі және белгілердің тұқымқуалау механизмдері туралы түсінік береді, бірақ ол 2-тарауда сипатталған тірі ағзалардың таңғаларлық әртүрлілігінің қалайша пайда болғандығын түсіндірмейді. Бұл сұраққа жауап беру мүмкіндіктері келесі үш тараудың мазмұнын қамтиды.
24-тарау
ҚОЛДАНБАЛЫ ГЕНЕТИКА
ХХ жүзжылдықтың екінші жартысында биология өзінің «Алтын ғасырына» келді. 1953 жылы ДНҚ құрылымының ашылуынан жақында пайда болған адамның генетикалық кодын ашу мүмкіндіктеріне дейінгі өткен уақытта, генетика және молекулалық биологияның түйісінде жаңа құдіретті ғылым саласы – биотехнология қалыптасты. Оның мағынасы, ең болмаса АҚШ-та академиялық зерттеулерге бөлінетін қаражаттың жартысы дерлік биотехнологияға шығындалатындығын көрсетеді. Биотехнология өнеркәсіпте, медицинада, ауыл шаруашылығында және басқа көптеген салаларда қолданыс табады. Оның жетістіктері адам игілігіне бағытталуы мүмкін, бірақ адамзатқа сансыз бәлелер әкелуі мүмкін. Дәл осы мәселемен, ХХ ғасырдың бірінші жартысында, атом құрылымының ашылуымен ядролық энергияны бейбіт мақсатта да, талқандау мақсатында да қолдану мүмкіндігі пайда болған кезде, физиктер қақтығысты. Джеймс Уотсонмен бірге ДНҚ құрылымының мағынасын ашқан үшін Нобель сыйлығын алған Фрэнсис Крик және Морис Уилкинс физиктер болған, және биологиямен айналыспастан бұрын, қару жасаумен айналысқан. Жинақталған тәжірибе бұл ғалымдарды өздерінің зерттеулерінің этикалық қырларына өте мұқият қарауға мәжбүр етті. Міне сондықтан да, 70-жылдардың соңында гендік инженерияның әдептілігінде және қауіпсіздігінде күдік пайда болған кезде, Крик және Уилкинс өздерінің жұмыстарын тоқтатты. Сіздер – болашақ биологсыздар және сіздер де бір кездері молекулалық биология саласындағы біздің біліміміздің тереңдеуіне қарай қалай да туатын, талқылау үстінде қабылданатын жаңа даулы мәселелерге жауапты боласыздар. Біз жеке көзқарасымызды қалыптастыра отырып, қаншалықты көп ақпаратты меңгерсек, және бұл мәселелерді қаншалықты көп адам талқылауға дайын болса, соншалықты бізбен қабылданған шешімдердің дұрыс болу мүмкіндігі көбірек.
Гендік инженерия
Бұл тараудың бірінші бөлімінде гендік инженерия туралы сөз қозғалатын болады. Бұл қолданбалы генетика және биотехнология арсеналында бар ең күшті тәсіл. Оның көмегімен өсімдіктер және жануарлар хромосомаларында кодталған генетикалық нұсқауларды зерттеуге және өзгертуге болады. Адамдар өзіне пайда ретінде басқа ағзаларды өзгерту және (болашақта) тұқымқуалайтын ауруларды емдеу мүмкіндігін алды (гендік терапия). Гендік инженерияның дамуындағы басты кезеңдер 25.1 кестесінде ұсынылған.
25.1. Бактериялардың гендік инженериясы
Гендік инженерияның негізгі тәсілдері 70-жылдардың басында жетілдірілді. Бұл тәсілдердің мәні – ағзаға жаңа ген енгізу. Мұндай ген жаңадан синтезделуі де, басқа ағзадан көшірілуі де мүмкін. Егер бактерия геномына қандай-да бір нәруызды кодтайтын генді салса, бактерия жасушасы бұл нәруызды шығаратын жанды фабрикаға айналдырады. Мысал ретінде бактерия жасушасына, адам инсулинін, адам бойының гормонын және бұқа соматотропинін (БСТ) кодтайтын гендерді көшіруді қарастырамыз (25.2.1-25.2.3 бөлімдері).
25.1 кестесі. Гендік инженерияның дамуындағы басты белгілер. (The encyclopaedia of molecular biology, J. Kendrew, 1994, Blackwell Science бойынша)
Гендік инженерия
Бұл тараудың бірінші бөлімінде гендік инженерия туралы сөз қозғалатын болады. Бұл қолданбалы генетика және биотехнология арсеналында бар ең күшті тәсіл. Оның көмегімен өсімдіктер және жануарлар хромосомаларында кодталған генетикалық нұсқауларды зерттеуге және өзгертуге болады. Адамдар өзіне пайда ретінде басқа ағзаларды өзгерту және (болашақта) тұқымқуалайтын ауруларды емдеу мүмкіндігін алды (гендік терапия). Гендік инженерияның дамуындағы басты кезеңдер 25.1 кестесінде ұсынылған.
25.1. Бактериялардың гендік инженериясы
Гендік инженерияның негізгі тәсілдері 70-жылдардың басында жетілдірілді. Бұл тәсілдердің мәні – ағзаға жаңа ген енгізу. Мұндай ген жаңадан синтезделуі де, басқа ағзадан көшірілуі де мүмкін. Егер бактерия геномына қандай-да бір нәруызды кодтайтын генді салса, бактерия жасушасы бұл нәруызды шығаратын жанды фабрикаға айналдырады. Мысал ретінде бактерия жасушасына, адам инсулинін, адам бойының гормонын және бұқа соматотропинін (БСТ) кодтайтын гендерді көшіруді қарастырамыз (25.2.1-25.2.3 бөлімдері).
25.1 кестесі. Гендік инженерияның дамуындағы басты белгілер. (The encyclopaedia of molecular biology, J. Kendrew, 1994, Blackwell Science бойынша)

25.1.1. Шолу
Қазіргі кезде кез-келген геннің көшірмесін алу – мүлдем қиын емес мәселе. Кейбір жағдайларда бұл үшін бір ғана бастапқы көшірме қажет. Бірдей көшірмелердің көбін алуды клондау деп атайды. Клондаушы вектор ретінде (яғни клондау қажет ДНҚ тасымалдаушысы) дәстүрлі түрде плазмидаларды немесе бактериофагтарды қолданады. Плазмидалар – бұл кейбір бактерияларда анықталған ДНҚ-ның кішкентай сақина тәрізді бөлігі. Олар негізгі (хромосомалық) ДНҚ-дан бөлектенген және одан тәуелсіз репликациялана алады (2.3.1 бөлімі). Бактериофагтар (немесе қысқалық үшін фагтар) – бұлар өз ДНҚ-сын осы ДНҚ репликацияланатын бактерия жасушасына енгізе алатын вирустар (2.19 суреті). Клондау қажет ДНҚ бөліктерін, не плазмидалық, не фагтік ДНҚ-мен біріктіреді. Әр-түрлі ағзалардың ДНҚ бөліктерінен тұратын «түзіліс», рекомбинантты ДНҚ деп аталады. Егер мұндай ДНҚ-ны бактерия жасушасына енгізсе, онда әр бөліну циклі сайын рекомбинантты ДНҚ көшірмесінің саны да көбейеді. Бактериялық геномға енгізілген бөгде ген өнеркәсіптік мөлшерде пайдалы нәруыз алуға қолданыла алады, мысалы қалыпты жағдайда бактериялық жасушамен өндірілмейтін адам инсулині.
Клондау процесі сызба түрінде 25.1 суретінде көрсетіледі және толығымен төменде сипатталады. Жаңа гендерді өсімдік және жануар эмбриондарына, трансгенді деп аталатын ағзаларды (бұл генді ұрпағына бере алатын ағзаларды) шығару мақсатымен енгізу қиынырақ мәселе болып табылады. Бұл тақырып 25.3 – 25.5 бөлімдерінде талқыланатын болады.
Бактериялардың гендік инженериясын бес кезеңге бөлуге болады.
Кезең 1. Ағзаның барлық қалған гендерінің ішінен қажет геннің көшірмесін бөліп алу.
Кезең 2. Қажет генді векторға орналастыру.
Кезең 3. Реципиентті жасушаға қажет генді енгізу үшін векторды қолдану.
Кезең 4. Бөгде ДНҚ (донорлық ДНҚ) кірген жасушаларды іріктеу.
Кезең 5. Генді клондау.
Жоғарыда ұсынылған кезеңдердің реттілігі – ең қарапайым. Алайда кейбір жағдайларда әрекеттер реттілігі басқаша болуы мүмкін; мысалы, векторға гендердің бүтін тобы (немесе кітапханасы) орналасып және клондалған болуы және содан кейін ғана қажетті ген бөлініп алынуы мүмкін.
25.1.2. Кезең 1. Қажетті геннің көшірмесін алу
Бұл процесстің ең қиын бөлігі. Мысалы, адам геномында (геном – бұл жасушадағы барлық ДНҚ) шамамен 3 млрд. Нуклеотид және ≈ 100 000 (соңғы мәліметтер бойынша ≈ 30 000) ген саналады. Қарапайым ген ұзындығында бірнеше мың жұп нуклеотидке ие, сондықтан да нақты генді табудың өзі онша оңай емес. Геннің көшірмелерін алу үшін үш тәсіл қолданылады:
кері транскриптаза көмегімен оның мРНҚ-сының матрицасынан ген көшірмесін алады;
жасанды ген синтездейді;
ДНҚ-ны рестрикционды ферменттермен (рестриктазалармен) кесуді және құрамында қажетті ген бар бөлікті іздеуді қамтитын «дробовик» («shotgun») тәсілін қолданады.
Гендік инженерия әдістемесін біз алдымен тікелей мақсатқа жетелейтін, яғни қажетті генді бөліп алуға мүмкіндік беретін алғашқы екі тәсілді қолдану мысалында нақты түсіндіреміз. Содан соң біз үшінші тәсілді қарастырамыз.
Кері транскриптазаның қолданылуы
Диплоидты жасушада әр геннің тек екі көшірмесі (бірі – анадан алынған хромосомада, екіншісі – әкесінен алынған хромосомада) болғанына қарамастан, белсенді қолданылатын ген әдетте мРНҚ-ның мыңдаған комплементарлы молекулаларын түзеді (23.8 бөлімі). Бәрінен жиірек берілген геннің қандай жасушаларда белсенді екендігі белгілі. Мысалы, инсулин кодтайтын ген ұйқы безінің β- жасушаларында белсенді. Ретровирустар құрамында РНҚ молекуласында ДНҚ-ның комплементарлы көшірмесін синтездей алатын фермент бар. РНҚ матрицасында ДНҚ алу – бұл қалыпты транскрипцияға, РНҚ-ның ДНҚ матрицасында синтезделгеніне қарама-қарсы процесс, сондықтан ферментті кері транскриптаза деп атаған. Ол гендік инженерияның бағалы құралы болды. (Ретровирустардың өздері бұл ферментті, өзінің РНҚ-дан тұратын геномын, жаңа жасушаларды жұқтыруға қабілетті ДНҚ-ға айналдыру үшін қолданады; 2.4.5 бөлімі.) Егер бізді қызықтырған геннің қай жасушаларда белсенді екені белгілі болса, бұл жасушалардан мРНҚ бөліп алу қиын емес. Бұл мәселе орындалған кезде, мРНҚ кері транскриптаза көмегімен қажетті геннің көшірмесін ДНҚ-ға айналдырады (25.1 суреті). Мұндай тәсілмен алынған ДНҚ-ны, оның біртізбекшелі немесе екітізбекшелі екендігіне қарамастан комплементарлы ДНҚ, немесе кДНҚ деп аталады.
Ген синтезі
Гендегі негіздер реттілігін не тікелей, не бұл генмен кодталатын нәруыздың аминқышқыл реттілігіне негізделе отырып анықтауға болады. Содан соң генді нуклеотидтерден (әр негіздің бір нуклеотид бөлігі екендігін еске түсіреміз), оларды бір-бірімен дұрыс тәртіппен байланыстыра отырып қарастыруға болады. Қазір мұндай тәсілмен тек қысқа гендерді құрастыру мүмкін, алайда тәсілдердің жетілдірілуімен кез-келген геннің синтезі ескішілдікке негізделген процедура болады. Бұл тәсіл проинсулиннің және соматостатиннің гендерін алу үшін қолданылды. Соматостатин (сонымен қатар бой гормонының ингибиторы ретінде белгілі) – бұл бар болғаны 14 аминқышқылдан тұратын нәруыздық гормон.
Қазіргі кезде кез-келген геннің көшірмесін алу – мүлдем қиын емес мәселе. Кейбір жағдайларда бұл үшін бір ғана бастапқы көшірме қажет. Бірдей көшірмелердің көбін алуды клондау деп атайды. Клондаушы вектор ретінде (яғни клондау қажет ДНҚ тасымалдаушысы) дәстүрлі түрде плазмидаларды немесе бактериофагтарды қолданады. Плазмидалар – бұл кейбір бактерияларда анықталған ДНҚ-ның кішкентай сақина тәрізді бөлігі. Олар негізгі (хромосомалық) ДНҚ-дан бөлектенген және одан тәуелсіз репликациялана алады (2.3.1 бөлімі). Бактериофагтар (немесе қысқалық үшін фагтар) – бұлар өз ДНҚ-сын осы ДНҚ репликацияланатын бактерия жасушасына енгізе алатын вирустар (2.19 суреті). Клондау қажет ДНҚ бөліктерін, не плазмидалық, не фагтік ДНҚ-мен біріктіреді. Әр-түрлі ағзалардың ДНҚ бөліктерінен тұратын «түзіліс», рекомбинантты ДНҚ деп аталады. Егер мұндай ДНҚ-ны бактерия жасушасына енгізсе, онда әр бөліну циклі сайын рекомбинантты ДНҚ көшірмесінің саны да көбейеді. Бактериялық геномға енгізілген бөгде ген өнеркәсіптік мөлшерде пайдалы нәруыз алуға қолданыла алады, мысалы қалыпты жағдайда бактериялық жасушамен өндірілмейтін адам инсулині.
Клондау процесі сызба түрінде 25.1 суретінде көрсетіледі және толығымен төменде сипатталады. Жаңа гендерді өсімдік және жануар эмбриондарына, трансгенді деп аталатын ағзаларды (бұл генді ұрпағына бере алатын ағзаларды) шығару мақсатымен енгізу қиынырақ мәселе болып табылады. Бұл тақырып 25.3 – 25.5 бөлімдерінде талқыланатын болады.
Бактериялардың гендік инженериясын бес кезеңге бөлуге болады.
Кезең 1. Ағзаның барлық қалған гендерінің ішінен қажет геннің көшірмесін бөліп алу.
Кезең 2. Қажет генді векторға орналастыру.
Кезең 3. Реципиентті жасушаға қажет генді енгізу үшін векторды қолдану.
Кезең 4. Бөгде ДНҚ (донорлық ДНҚ) кірген жасушаларды іріктеу.
Кезең 5. Генді клондау.
Жоғарыда ұсынылған кезеңдердің реттілігі – ең қарапайым. Алайда кейбір жағдайларда әрекеттер реттілігі басқаша болуы мүмкін; мысалы, векторға гендердің бүтін тобы (немесе кітапханасы) орналасып және клондалған болуы және содан кейін ғана қажетті ген бөлініп алынуы мүмкін.
25.1.2. Кезең 1. Қажетті геннің көшірмесін алу
Бұл процесстің ең қиын бөлігі. Мысалы, адам геномында (геном – бұл жасушадағы барлық ДНҚ) шамамен 3 млрд. Нуклеотид және ≈ 100 000 (соңғы мәліметтер бойынша ≈ 30 000) ген саналады. Қарапайым ген ұзындығында бірнеше мың жұп нуклеотидке ие, сондықтан да нақты генді табудың өзі онша оңай емес. Геннің көшірмелерін алу үшін үш тәсіл қолданылады:
кері транскриптаза көмегімен оның мРНҚ-сының матрицасынан ген көшірмесін алады;
жасанды ген синтездейді;
ДНҚ-ны рестрикционды ферменттермен (рестриктазалармен) кесуді және құрамында қажетті ген бар бөлікті іздеуді қамтитын «дробовик» («shotgun») тәсілін қолданады.
Гендік инженерия әдістемесін біз алдымен тікелей мақсатқа жетелейтін, яғни қажетті генді бөліп алуға мүмкіндік беретін алғашқы екі тәсілді қолдану мысалында нақты түсіндіреміз. Содан соң біз үшінші тәсілді қарастырамыз.
Кері транскриптазаның қолданылуы
Диплоидты жасушада әр геннің тек екі көшірмесі (бірі – анадан алынған хромосомада, екіншісі – әкесінен алынған хромосомада) болғанына қарамастан, белсенді қолданылатын ген әдетте мРНҚ-ның мыңдаған комплементарлы молекулаларын түзеді (23.8 бөлімі). Бәрінен жиірек берілген геннің қандай жасушаларда белсенді екендігі белгілі. Мысалы, инсулин кодтайтын ген ұйқы безінің β- жасушаларында белсенді. Ретровирустар құрамында РНҚ молекуласында ДНҚ-ның комплементарлы көшірмесін синтездей алатын фермент бар. РНҚ матрицасында ДНҚ алу – бұл қалыпты транскрипцияға, РНҚ-ның ДНҚ матрицасында синтезделгеніне қарама-қарсы процесс, сондықтан ферментті кері транскриптаза деп атаған. Ол гендік инженерияның бағалы құралы болды. (Ретровирустардың өздері бұл ферментті, өзінің РНҚ-дан тұратын геномын, жаңа жасушаларды жұқтыруға қабілетті ДНҚ-ға айналдыру үшін қолданады; 2.4.5 бөлімі.) Егер бізді қызықтырған геннің қай жасушаларда белсенді екені белгілі болса, бұл жасушалардан мРНҚ бөліп алу қиын емес. Бұл мәселе орындалған кезде, мРНҚ кері транскриптаза көмегімен қажетті геннің көшірмесін ДНҚ-ға айналдырады (25.1 суреті). Мұндай тәсілмен алынған ДНҚ-ны, оның біртізбекшелі немесе екітізбекшелі екендігіне қарамастан комплементарлы ДНҚ, немесе кДНҚ деп аталады.
Ген синтезі
Гендегі негіздер реттілігін не тікелей, не бұл генмен кодталатын нәруыздың аминқышқыл реттілігіне негізделе отырып анықтауға болады. Содан соң генді нуклеотидтерден (әр негіздің бір нуклеотид бөлігі екендігін еске түсіреміз), оларды бір-бірімен дұрыс тәртіппен байланыстыра отырып қарастыруға болады. Қазір мұндай тәсілмен тек қысқа гендерді құрастыру мүмкін, алайда тәсілдердің жетілдірілуімен кез-келген геннің синтезі ескішілдікке негізделген процедура болады. Бұл тәсіл проинсулиннің және соматостатиннің гендерін алу үшін қолданылды. Соматостатин (сонымен қатар бой гормонының ингибиторы ретінде белгілі) – бұл бар болғаны 14 аминқышқылдан тұратын нәруыздық гормон.
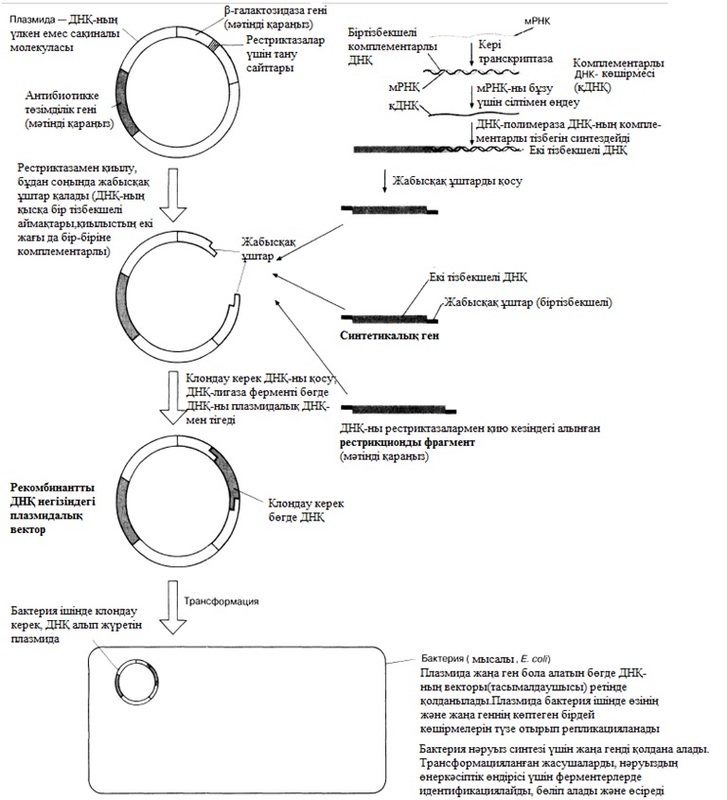
25.1 суреті. Гендік инженерия. Гендерді клондау үшін құрастырылған процедуралар сызбасы. Нақтырақ мәтінде түсіндірілген.
«Дробовик» тәсілі – рестрикционды ферменттердің қолданылуы
Бұл гендерді бөліп алудың алғаш тәсілі – 1960-жылдардың соңы – 1970-жылдардың басында құрастырылды. Оның пайда болуы рестрикционды (ағылшыншадан аударғанда restricting - шектейтін) эндонуклеаза немесе рестриктаза деп аталатын ферменттердің ашылуы арқасында мүмкін болды. Бактерияларда анықталған бұл ферменттер, ДНҚ-ны кесуге қабілетті, мысалы олар бактериялық жасушаға басып кіретін кез-келген вирустық ДНҚ-ны кеседі, осылайша жасуша ішінде вирустардың көбеюін шектейді (рестрикциялайды). Бактериялардың әр түрі әр-түрлі рестрикционды эндонуклеазаны продукциялайды. Олардың әрқайсысы нуклеин қышқылын («нуклеаза» сөзі осыдан) қатаң анықталған нүктелерде кеседі («эндо» дегеніміз, фермент молекуланы соңынан шабуылдамай, ішінен бөлетіндігін білдіреді). Фермент негіздердің қандай-да бір реттілігін таниды және дәл солмен ғана әрекеттеседі. Кесілу нүктелері рестрикциялар сайты деп аталады. Қазіргі кезде 230 әртүрлі реттіліктер қатынасындағы 2000-нан астам рестриктазалар бөліп алынды. Өзінің өзіндік ДНҚ-сын бактерия, метильді топтың рестрикция сайттарында анықталған негіздерге қосылу жолымен қорғайды.
Әр ферменттің атауы, оны бөліп алған бактерияның атауынан шығады (25.2 суреті). Рестрикцияланатын реттілік-нысана жиі ұзындығында алты негізге ие екендігіне және палиндромды болып табылатындығына, яғни екі бағытта да бірдей оқылатындығына назар аударыңыз. 25.2 суретін қарастыра отырып, ДНҚ-ның екі комплементарлы тізбектері қарама-қарсы бағытта оқылатындығын есте ұстаңыз. Кейбір рестриктазалар сатылы кесінділер жасайды. Олардың әсерінің нәтижесінде шығыңқы біртізбекшелі ұшы бар ДНҚ бөліктері түзіледі (мысалы, EcoRJ, 25.2 суреті). Мұндай ұштар «жабысқақ» деп аталады; оларды ДНҚ бөліктерін қайта біріктіру үшін қолданады. Олар сол рестриктазамен кесілген ДНҚ-ның басқа молекулаларының комплементарлы жабысқақ ұшы бар сутектік байланыстардың түзілуі есебінен жабысқандай болады (25.1 суреті). Мысалы, кеңінен қолданылатын рестриктаза EcoRJ жабысқақ ұш – ТТАА түзеді. Өтпейтін ұш жасайтын рестриктазалар бар. Оларға, мысалы, HindJJ жатады (25.2 суреті). Кез-келген ағзаның ДНҚ-сын рестриктазаларының көмегімен бөлшектеуге болады. Түзілген рестрикциялық бөлшектердің ұзындығы қолданылатын рестрикционды эндонуклеазаның түріне және берілген ферментпен танылатын негіздердің реттілігінің қайда орналасқандығына байланысты (25.3 суреті).
ДНҚ-дағы әр нуклеотидтің теріс зарядталған фосфатты топты алып жүретіндігі белгілі, сондықтан әр-түрлі ұзындықты ДНҚ бөліктері бірдей зарядталмаған. Бұл айырмашылықтарды ДНҚ бөліктерін электрлік жазықтықта гельэлектрофорез тәсілімен алу үшін қолдануға болады (25.4 суреті). Тәсілдің атына қарай, ол агарозды (өте үлкен бөліктерге) немесе полиакриламидті (кіші өлшемді бөлшектерге) гельді қолдануды ұйғарады. Қанша дегенмен ДНҚ түссіз болғандықтан, гельдегі осы немесе өзге бөліктің электрофорезден кейінгі орнын не бояу, не радиоактивті таңбаланған ДНҚ қолдану және гельге фотосуреттік қабықшаны қоятын радиоавтография жүргізу көмегімен анықтайды. Радиоактивті сәулелену қабықшаны, ДНҚ орналасқан жерде жарықтандырады.
Донорлық ағзаның ДНҚ-сын рестриктазалармен өңдеген кезде, бөліктердің біреуінің құрамында кездейсоқ түрде қажет геннің толық көшірмесінен артық ешнәрсе болмайтындығымен көзделеді. Бұл гендерді бөліп алудың ерекше емес тәсілін дробовик тәсілі деп атайды. Оны зерттеудегі қиынырақ мәселе – қажет генді алып жүретін бөлікті табу, бұл мәселе 25.1.3 бөлімінде талқыланады.
Бұл гендерді бөліп алудың алғаш тәсілі – 1960-жылдардың соңы – 1970-жылдардың басында құрастырылды. Оның пайда болуы рестрикционды (ағылшыншадан аударғанда restricting - шектейтін) эндонуклеаза немесе рестриктаза деп аталатын ферменттердің ашылуы арқасында мүмкін болды. Бактерияларда анықталған бұл ферменттер, ДНҚ-ны кесуге қабілетті, мысалы олар бактериялық жасушаға басып кіретін кез-келген вирустық ДНҚ-ны кеседі, осылайша жасуша ішінде вирустардың көбеюін шектейді (рестрикциялайды). Бактериялардың әр түрі әр-түрлі рестрикционды эндонуклеазаны продукциялайды. Олардың әрқайсысы нуклеин қышқылын («нуклеаза» сөзі осыдан) қатаң анықталған нүктелерде кеседі («эндо» дегеніміз, фермент молекуланы соңынан шабуылдамай, ішінен бөлетіндігін білдіреді). Фермент негіздердің қандай-да бір реттілігін таниды және дәл солмен ғана әрекеттеседі. Кесілу нүктелері рестрикциялар сайты деп аталады. Қазіргі кезде 230 әртүрлі реттіліктер қатынасындағы 2000-нан астам рестриктазалар бөліп алынды. Өзінің өзіндік ДНҚ-сын бактерия, метильді топтың рестрикция сайттарында анықталған негіздерге қосылу жолымен қорғайды.
Әр ферменттің атауы, оны бөліп алған бактерияның атауынан шығады (25.2 суреті). Рестрикцияланатын реттілік-нысана жиі ұзындығында алты негізге ие екендігіне және палиндромды болып табылатындығына, яғни екі бағытта да бірдей оқылатындығына назар аударыңыз. 25.2 суретін қарастыра отырып, ДНҚ-ның екі комплементарлы тізбектері қарама-қарсы бағытта оқылатындығын есте ұстаңыз. Кейбір рестриктазалар сатылы кесінділер жасайды. Олардың әсерінің нәтижесінде шығыңқы біртізбекшелі ұшы бар ДНҚ бөліктері түзіледі (мысалы, EcoRJ, 25.2 суреті). Мұндай ұштар «жабысқақ» деп аталады; оларды ДНҚ бөліктерін қайта біріктіру үшін қолданады. Олар сол рестриктазамен кесілген ДНҚ-ның басқа молекулаларының комплементарлы жабысқақ ұшы бар сутектік байланыстардың түзілуі есебінен жабысқандай болады (25.1 суреті). Мысалы, кеңінен қолданылатын рестриктаза EcoRJ жабысқақ ұш – ТТАА түзеді. Өтпейтін ұш жасайтын рестриктазалар бар. Оларға, мысалы, HindJJ жатады (25.2 суреті). Кез-келген ағзаның ДНҚ-сын рестриктазаларының көмегімен бөлшектеуге болады. Түзілген рестрикциялық бөлшектердің ұзындығы қолданылатын рестрикционды эндонуклеазаның түріне және берілген ферментпен танылатын негіздердің реттілігінің қайда орналасқандығына байланысты (25.3 суреті).
ДНҚ-дағы әр нуклеотидтің теріс зарядталған фосфатты топты алып жүретіндігі белгілі, сондықтан әр-түрлі ұзындықты ДНҚ бөліктері бірдей зарядталмаған. Бұл айырмашылықтарды ДНҚ бөліктерін электрлік жазықтықта гельэлектрофорез тәсілімен алу үшін қолдануға болады (25.4 суреті). Тәсілдің атына қарай, ол агарозды (өте үлкен бөліктерге) немесе полиакриламидті (кіші өлшемді бөлшектерге) гельді қолдануды ұйғарады. Қанша дегенмен ДНҚ түссіз болғандықтан, гельдегі осы немесе өзге бөліктің электрофорезден кейінгі орнын не бояу, не радиоактивті таңбаланған ДНҚ қолдану және гельге фотосуреттік қабықшаны қоятын радиоавтография жүргізу көмегімен анықтайды. Радиоактивті сәулелену қабықшаны, ДНҚ орналасқан жерде жарықтандырады.
Донорлық ағзаның ДНҚ-сын рестриктазалармен өңдеген кезде, бөліктердің біреуінің құрамында кездейсоқ түрде қажет геннің толық көшірмесінен артық ешнәрсе болмайтындығымен көзделеді. Бұл гендерді бөліп алудың ерекше емес тәсілін дробовик тәсілі деп атайды. Оны зерттеудегі қиынырақ мәселе – қажет генді алып жүретін бөлікті табу, бұл мәселе 25.1.3 бөлімінде талқыланады.

25.2 суреті. Кейбір әдетте қолданылатын рестриктазалар. EcoRI, HindIII және BamHI, «жабысқақ ұштар» қалдыра отырып, ДНҚ-да ирек тәрізді (сатылы) қиықтар жасайды. Түзілген жабысқақ ұш, мысалы EcoRI, басқа түзілген жабысқақ ұш EcoRI-мен біріге алады. HindII және HpaI дөңес ұштар қалдырады. EcoRI және HpaI үшін нақтырақ сызбалар ұсынылған.
Үзілмелі гендер
Кері транскриптазаны қолдану немесе геннің химиялық синтезі дробовик тәсілі бойынша артықшылықтарға ие, қанша дегенмен бұдан алынатын ген «үзілмелі» емес. Үзілмелі гендер құрамында нәруыздың аминқышқылдық реттілігін кодтамайтын, интрондар деп аталатын ДНҚ-ның бір немесе бірнеше аймақтары бар. Интрондар дың қызметі әлі күнге дейін белгісіз. Тек қана құрамында интрондар бар эукариоттық генді бактерияға орналастырса, онда бактерия пайдасыз нәруыз синтездейтіндігі белгілі, қанша дегенмен бактерияларда мРНҚ-дан интрондарды жоюға арналған ферменттер жоқ. Интрондардың мРНҚ-дан қалай жоғалатындығы 25.5 суретінде көрсетілген.
Кері транскриптазаны қолдану немесе геннің химиялық синтезі дробовик тәсілі бойынша артықшылықтарға ие, қанша дегенмен бұдан алынатын ген «үзілмелі» емес. Үзілмелі гендер құрамында нәруыздың аминқышқылдық реттілігін кодтамайтын, интрондар деп аталатын ДНҚ-ның бір немесе бірнеше аймақтары бар. Интрондар дың қызметі әлі күнге дейін белгісіз. Тек қана құрамында интрондар бар эукариоттық генді бактерияға орналастырса, онда бактерия пайдасыз нәруыз синтездейтіндігі белгілі, қанша дегенмен бактерияларда мРНҚ-дан интрондарды жоюға арналған ферменттер жоқ. Интрондардың мРНҚ-дан қалай жоғалатындығы 25.5 суретінде көрсетілген.
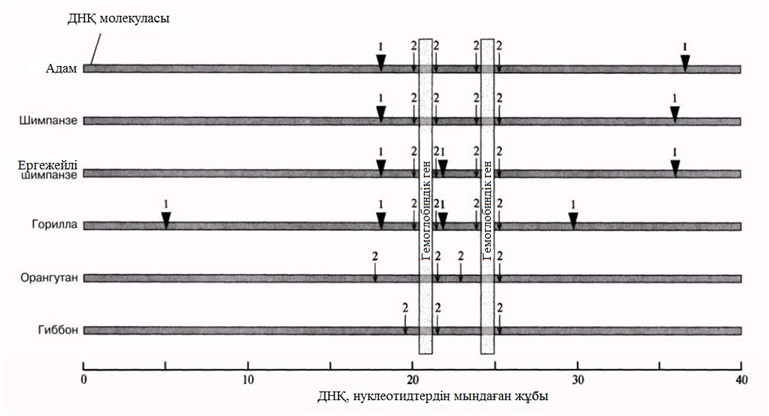
25.3 суреті. Рестриктазаларды ДНҚ-ны қиюға және әр-түрлі ұзындықтағы рестрикционды фрагменттерді алуға қолдану. Бұл суретте құрамында әр-түрлі маймылдардың екі гемоглобиндік гені бар ДНҚ аймағын екі рестриктазамен өңдеген. Фермент 1 ▼белгісімен, фермент 2 ↓ белгісімен белгіленген. Фермент 1 адам ДНҚ-сын екі фрагментке бөледі, ал фермент 2 - бес фрагментке. Бұл фрагменттердің, сонымен қатар екі ферментпен бірге өңдеудегі алынған фрагменттердің ұзындықтары, қиындылардың орнын (рестрикция сайттарын) бір-біріне қатысты орналастыра алады. Бұл мәліметтер негізінде, сызбада суреттелген «рестрикционды карталар» құрады. Картаны құруға қаншалықты көп рестриктазалар қолданылса, ол соншалықты нақтырақ болады. Түрлер арасындағы туыстық қаншалықты жақын болса, олардың ДНҚ-лары және олардағы рестрикция сайттарының орналасуы соншалықты ұқсас екендігіне назар аударыңыз. [Сызба 7 – 4 суреті негізінде құрастырылған, 294 бет, Molecular biology of the cell, 3rd ed., B. Alberts et al. (1994) Garland.]
25.4 суреті. Әр-түрлі ұзындықтағы ДНҚ фрагменттерін қию үшін қолданылатын гель-электрофорез. Фрагменттер ДНҚ-ны бір немесе бірнеше рестриктазалармен қиғанда түзіледі. Ең үлкен фрагменттер барлығынан жай қозғалады, өйткені оларға гельдегі саңылаулар арқылы өту қиынырақ. Фотосуретте олар гельдегі бастапқы қуыстардан жақында орналасқан.
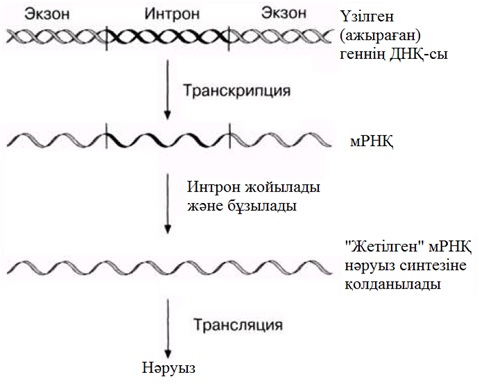
25.5суреті. Құрамында интрон бар геннің транскрипциясы және трансляциясы. Интронды қоршайтын аймақтар экзондар деп аталады. Гендер құрамында көп интрон болуы мүмкін. Нәруыздар тек қана экзондармен кодталады.
25.1.3. Кезең 2. Гендерді векторға орналастыру
Жоғарыда түсіндірілгендегі векторлар ретінде бәрінен де жиірек плазмидалық ДНҚ-ны немесе фагтік ДНҚ-ны қолданады. Алдымен плазмалық ДНҚ-ға орналастыру сызбасын қарастырамыз. Негізінде фагтік ДНҚ кезінде де дәл сол процедура қайталанады.
Плазмидалық ДНҚ
Бактериялардағы плазмалық ДНҚ-ның сақина тәрізді молекулалары, негізгі хромосомалық ДНҚ-ға қарағанда азырақ, сондықтан оларды бір-бірінен оңай ажыратуға болады. Бактериялық жасушалар бұзады және центрифугалайды. Сонымен бірге хромосомалық ДНҚ тұнбада болады, ал плазмидалық ДНҚ тұнбаның жоғарысында сұйық фазада қалады. Рестриктазалармен өңдеуден бұрын плазмидалық ДНҚ-ны тазалайды (25.1 суреті).
Егер донорлық ДНҚ-ны бөліп алу үшін (мысалы, «дробовик» тәсілімен) рестриктазаны қолданған болса, дәл сол рестриктазамен плазмидалық ДНҚ өңделуі тиіс. Донорлық ДНҚ-ның, арасында қажетті генді бірлік те бар рестрикционды бөліктерді кейіннен плазмидалық ДНҚ-мен араластырады. Араласқан кезде олар жабысқақ ұштарымен байланысады. Мысалы ААТТ жабысқақ ұшы комплементарлы жабысқақ ұш – ТТАА-мен байланысатын болады. Басында байланысу сутектік байланыстар есебінен болады, бірақ ДНҚ-лигаза деп аталатын ферментті қосқаннан кейін, фосфодиэфирлі байланыстар түзіледі.
Егер донорлық ДНҚ ретінде кДНҚ немесе синтетикалық ген қолданылса (жоғарыдағы гендерді бөліп алудың алғашқы екі тәсілін қараңыз), онда ДНҚ-ны векторға орналастыру үшін 25.6 суретінде ұсынылған сызба бойынша әрекет ету керек. Егер «дробовик» тәсілін қолданғанда өтпейтін ұш түзетін фермент пайдаланылған болса дәл осы процедура қажет.
Фагтік вектор
Фагтардың векторлар ретіндегі артықшылығы плазмидаларды тасымалдай алатындармен салыстырғанда, ДНҚ-ның көп бөліктерін клонда мүмкіндігі болып табылады. Басқалардан жиірек бұл мақсат үшін λ фагін қолданады (2.20 суреті). Фагтік ДНҚ-ның бөлшегін клондау қажет ДНҚ алмастырады. Бұл бөлшек қожайынның бактериялық жасушасындағы фагтік ДНҚ репликациясы үшін қажет емес. Сызба түрінде бұл процедура, 25.7 суретінде ұсынылған Онда жиі «дробовик» тәсілімен алынған донорлық ДНҚ-ны қолданады.
Жоғарыда түсіндірілгендегі векторлар ретінде бәрінен де жиірек плазмидалық ДНҚ-ны немесе фагтік ДНҚ-ны қолданады. Алдымен плазмалық ДНҚ-ға орналастыру сызбасын қарастырамыз. Негізінде фагтік ДНҚ кезінде де дәл сол процедура қайталанады.
Плазмидалық ДНҚ
Бактериялардағы плазмалық ДНҚ-ның сақина тәрізді молекулалары, негізгі хромосомалық ДНҚ-ға қарағанда азырақ, сондықтан оларды бір-бірінен оңай ажыратуға болады. Бактериялық жасушалар бұзады және центрифугалайды. Сонымен бірге хромосомалық ДНҚ тұнбада болады, ал плазмидалық ДНҚ тұнбаның жоғарысында сұйық фазада қалады. Рестриктазалармен өңдеуден бұрын плазмидалық ДНҚ-ны тазалайды (25.1 суреті).
Егер донорлық ДНҚ-ны бөліп алу үшін (мысалы, «дробовик» тәсілімен) рестриктазаны қолданған болса, дәл сол рестриктазамен плазмидалық ДНҚ өңделуі тиіс. Донорлық ДНҚ-ның, арасында қажетті генді бірлік те бар рестрикционды бөліктерді кейіннен плазмидалық ДНҚ-мен араластырады. Араласқан кезде олар жабысқақ ұштарымен байланысады. Мысалы ААТТ жабысқақ ұшы комплементарлы жабысқақ ұш – ТТАА-мен байланысатын болады. Басында байланысу сутектік байланыстар есебінен болады, бірақ ДНҚ-лигаза деп аталатын ферментті қосқаннан кейін, фосфодиэфирлі байланыстар түзіледі.
Егер донорлық ДНҚ ретінде кДНҚ немесе синтетикалық ген қолданылса (жоғарыдағы гендерді бөліп алудың алғашқы екі тәсілін қараңыз), онда ДНҚ-ны векторға орналастыру үшін 25.6 суретінде ұсынылған сызба бойынша әрекет ету керек. Егер «дробовик» тәсілін қолданғанда өтпейтін ұш түзетін фермент пайдаланылған болса дәл осы процедура қажет.
Фагтік вектор
Фагтардың векторлар ретіндегі артықшылығы плазмидаларды тасымалдай алатындармен салыстырғанда, ДНҚ-ның көп бөліктерін клонда мүмкіндігі болып табылады. Басқалардан жиірек бұл мақсат үшін λ фагін қолданады (2.20 суреті). Фагтік ДНҚ-ның бөлшегін клондау қажет ДНҚ алмастырады. Бұл бөлшек қожайынның бактериялық жасушасындағы фагтік ДНҚ репликациясы үшін қажет емес. Сызба түрінде бұл процедура, 25.7 суретінде ұсынылған Онда жиі «дробовик» тәсілімен алынған донорлық ДНҚ-ны қолданады.
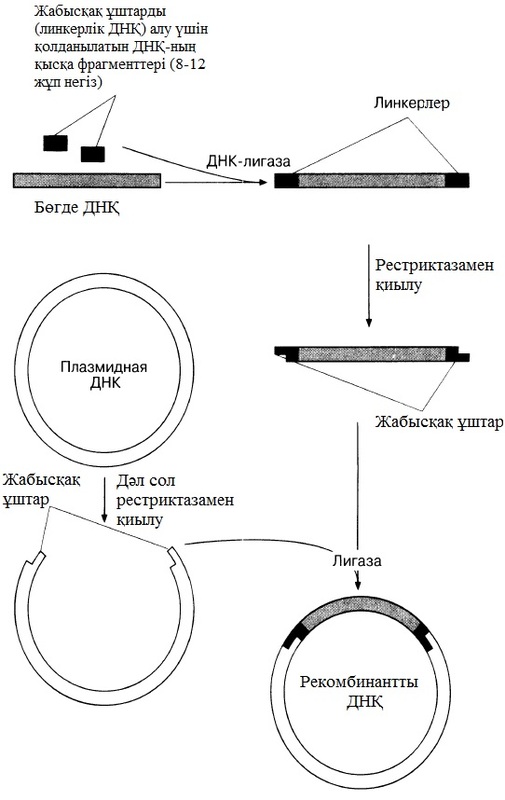
25.6 суреті. ДНҚ-ны векторға орналастырмастан бұрын, ДНҚ-ның рекомбинантты молекуласын жасау мақсатымен, жабысқақ ұштарды дөңес ұштарға ие ДНҚ фрагментіне қосу.
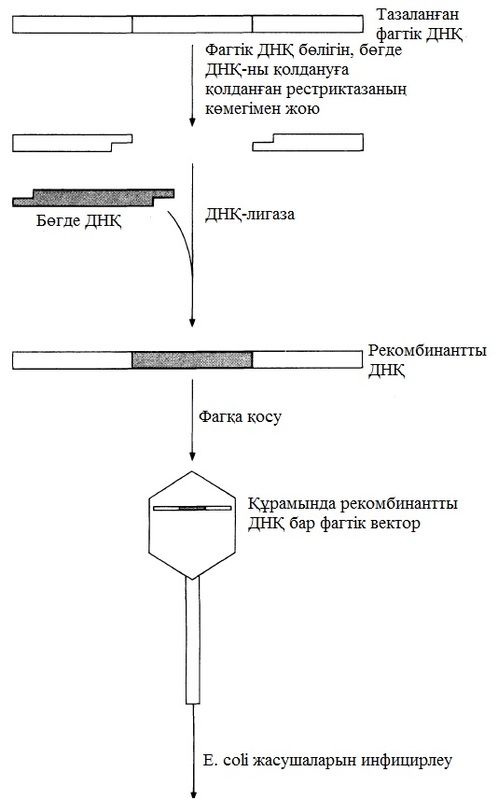
25.7суреті. ДНҚ-ны фагтік векторға қосу.
25.1.4. Кезең 3. Векторды қожайын жасушасына енгізу
Бұл кезеңде фагтік немесе плазмидалық векторды олар көбейе (өзін және оның құрамындағы бөгде донорлық ДНҚ-ны клондау) алатын бактериялық жасушаға енгізеді. Ереже бойынша, бұл мақсаттар үшін адам ішегінің қарапайым мекендеушісі – Escherichia coli-ді қолданады. Бұл мақсаттар үшін ішек таяқшасы бұл бактерияның генетикасы жақсы зерттелгендіктен, сонымен қатар ол тез өсетін (екіеселену уақыты шамамен 30 мин құрайды) болғандықтан таңдайды. Гендік инженерия үшін тек қана ерекше зертханалық жағдайларда тірі қалатын арнай мутант E.coli алынды. Сондықтан да ол егер оның құрамындағы бөгде гендермен бірге қоршаған ортаға кездейсоқ түсетін болса адамды жұқтыруға қабілетсіз.
Плазмидалық векторды қолданған мкезде, плазмида препаратын құрамында E.coli өсіріндісі бар пробиркаға қосады. Сол жерге кальций иондарын (әдетте кальций хлориді түрінде) енгізеді және жасушалар қысқа жылулық шокқа душар болады. Нәтижесінде E.coli-дің жасуша мембранасында, плазмидалар жасуша ішіне ене алатын саңылаулар тез түзіледі. Жаңа ДНҚ-ны бактерия жасушасына енгізу процесі трансформация деп аталады.
Фагтік векторларды жасушаларға, агаризондалған ортасы бар табақшаларда өсетін бактериялық газонды жұқтыру жолымен енгізеді (12.8 бөлімі).
25.1.5. Кезең 4. ДНҚ клондау
Құрамында ДНҚ-ның рекомбинантты молекуласы бар бір ғана жалғыз фагтік бөлшек, бір күннен азырақ уақытта өзінің және осы молекуланың 1012-нен көбірек бірдей көшірмелерін түзе алады. Құрамында плазмидалар бар E.coli жасушаларын, әдетте Петри табақшаларындағы қоректік агарға себеді. Мұнда жасушалар уақыт өте келе көрінетін отарлар түзе отырып, әр 30 мин сайын бөлінеді. Сонымен бірге фагтік векторларды қолданғандағыдай, қажетті ДНҚ-ның дәл сондай саны пайда болады, және оның үстіне бактерия бөлінген кезде оларда плазмиданың жүздеген көшірмелері синтезделеді. Бұл екі тәсілдер көмегімен қысқа уақыт ішінде миллиардтаған клондар алынады. Алдағы клондауды жүргізбестен бұрын, түрлендірілген бактериялардың іріктеуін жүргізу қажет.\
Түрлендірілген бактерияларды іріктеу
Вектор ретінде плазмидалар қолданылған жағдайды қарастырайық. Плазмидалық ДНҚ-ны бактериялы өсіріндімен араластырған кезде екі мәселе туады. Біріншіден барлық бактериялар түрлендірілмейді (плазмидалар алады). Екіншіден, барлық плазмидалар бөгде донорлық ДНҚ-ны алып жүрмейді. Бұл мәселелер, екі ерекше құрылымға ие плазмидаларды епті түрде айналып өтеді (25.8 суреті).
Бұл кезеңде фагтік немесе плазмидалық векторды олар көбейе (өзін және оның құрамындағы бөгде донорлық ДНҚ-ны клондау) алатын бактериялық жасушаға енгізеді. Ереже бойынша, бұл мақсаттар үшін адам ішегінің қарапайым мекендеушісі – Escherichia coli-ді қолданады. Бұл мақсаттар үшін ішек таяқшасы бұл бактерияның генетикасы жақсы зерттелгендіктен, сонымен қатар ол тез өсетін (екіеселену уақыты шамамен 30 мин құрайды) болғандықтан таңдайды. Гендік инженерия үшін тек қана ерекше зертханалық жағдайларда тірі қалатын арнай мутант E.coli алынды. Сондықтан да ол егер оның құрамындағы бөгде гендермен бірге қоршаған ортаға кездейсоқ түсетін болса адамды жұқтыруға қабілетсіз.
Плазмидалық векторды қолданған мкезде, плазмида препаратын құрамында E.coli өсіріндісі бар пробиркаға қосады. Сол жерге кальций иондарын (әдетте кальций хлориді түрінде) енгізеді және жасушалар қысқа жылулық шокқа душар болады. Нәтижесінде E.coli-дің жасуша мембранасында, плазмидалар жасуша ішіне ене алатын саңылаулар тез түзіледі. Жаңа ДНҚ-ны бактерия жасушасына енгізу процесі трансформация деп аталады.
Фагтік векторларды жасушаларға, агаризондалған ортасы бар табақшаларда өсетін бактериялық газонды жұқтыру жолымен енгізеді (12.8 бөлімі).
25.1.5. Кезең 4. ДНҚ клондау
Құрамында ДНҚ-ның рекомбинантты молекуласы бар бір ғана жалғыз фагтік бөлшек, бір күннен азырақ уақытта өзінің және осы молекуланың 1012-нен көбірек бірдей көшірмелерін түзе алады. Құрамында плазмидалар бар E.coli жасушаларын, әдетте Петри табақшаларындағы қоректік агарға себеді. Мұнда жасушалар уақыт өте келе көрінетін отарлар түзе отырып, әр 30 мин сайын бөлінеді. Сонымен бірге фагтік векторларды қолданғандағыдай, қажетті ДНҚ-ның дәл сондай саны пайда болады, және оның үстіне бактерия бөлінген кезде оларда плазмиданың жүздеген көшірмелері синтезделеді. Бұл екі тәсілдер көмегімен қысқа уақыт ішінде миллиардтаған клондар алынады. Алдағы клондауды жүргізбестен бұрын, түрлендірілген бактериялардың іріктеуін жүргізу қажет.\
Түрлендірілген бактерияларды іріктеу
Вектор ретінде плазмидалар қолданылған жағдайды қарастырайық. Плазмидалық ДНҚ-ны бактериялы өсіріндімен араластырған кезде екі мәселе туады. Біріншіден барлық бактериялар түрлендірілмейді (плазмидалар алады). Екіншіден, барлық плазмидалар бөгде донорлық ДНҚ-ны алып жүрмейді. Бұл мәселелер, екі ерекше құрылымға ие плазмидаларды епті түрде айналып өтеді (25.8 суреті).
- Плазмидалар белгілі антибиотикке тұрақтылық генін алып жүреді. Егер бактерияларды өсіретін қоректік ортаға бұл антибиотикті қосса, онда тек қана түрленген жасушалар (құрамында плазмидалар бар) көбейе және отар түзе алады.
- Плазмидалар β-галактозидаза ферментін кодтайтын генді алып жүреді, әрі бұл ген құрамында қосымша ген қызметіне әсер етпейтін бірнеше рестрикция сайттары болады. Β-Галактозидаза – бұл лактозаны галактозаға және глюкозаға ыдырататын фермент. Берілген ферменттің түссіз зат Х-gal-ге әрекетінен көгілдір түсті өнім түзіледі. Егер бөгде ДНҚ геннің ішіндегі рестрикция сайтына орналастырылса, онда ген жұмыс жасамайды. Сондықтан, егер антибиотик бар жерде өскен бактерияларды, кейіннен Х-gal бар ортада өсірсе, онда құрамында донорлық ДНҚ жоқ отарлар көгілдір түске боялған болады. Түссіз колониялар түзетін бактериялар құрамында донорлық ДНҚ болады, және оларды кейінгі клондау үшін таңдауға болады.
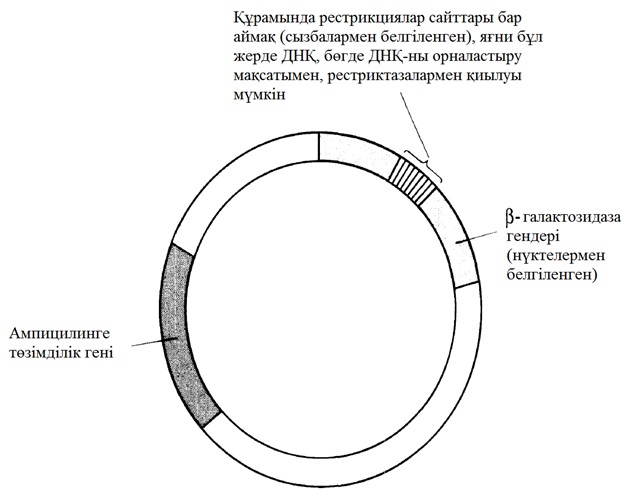
25.8 суреті. Құрамында ампициллин антибиотигіне төзімділік гені бар плазмидалық вектор. Мұндай геннің бар болуының арқасында, біз ампициллин бар ортада плазмиданы алып жүретін жасушаларды іріктей аламыз. Құрамныда плазмида жоқ жасушалар, мұндай ортада өледі.
25.1.6. Құрамында қажет ген бар бактерияларды іріктеу
Егер клондаудың бірінші кезеңінде «дробовик» тәсілі қолданылса, онда донорлық ДНҚ-ны қосып алатын бактерияларға қажетті гені бар ДНҚ бөлігін міндетті түрде алып жүруі мүлдем қажет емес. Донорлық ДНҚ рестрикционды бөліктердің өте көп санының қоспасын құрайды (адам ДНҚ-сы кезінде – миллионға дейін). Бұл бөліктердің барлығы клондалатын жағдайдың өзінде, қажетті ген немесе ДНҚ бөлігі құрамында олардың біреуі немесе бірнешеуі болуы мүмкін. Мұндай жолмен алынған клондар қоспасы кітапхана деп аталады. Кітапхананы сонымен қатар, бірінші кезеңде кері транскриптазаны және мРНҚ қоспасын қолданған кезде де алуға болады. Кейде бұны, қажетті мРНҚ-ны таза күйінде бөліп алу мүмкін болмағанда, жасауға тура келеді. Осылайша, жалғыз емес ген (бір типтің мРНҚ-сымен синтезделген немесе есептелген) клондалған жағдайларда, төртінші кезеңнен кейін кітапханалар түріндегі бактериялық өсірінділерді алады.
Егер клондаудың бірінші кезеңінде «дробовик» тәсілі қолданылса, онда донорлық ДНҚ-ны қосып алатын бактерияларға қажетті гені бар ДНҚ бөлігін міндетті түрде алып жүруі мүлдем қажет емес. Донорлық ДНҚ рестрикционды бөліктердің өте көп санының қоспасын құрайды (адам ДНҚ-сы кезінде – миллионға дейін). Бұл бөліктердің барлығы клондалатын жағдайдың өзінде, қажетті ген немесе ДНҚ бөлігі құрамында олардың біреуі немесе бірнешеуі болуы мүмкін. Мұндай жолмен алынған клондар қоспасы кітапхана деп аталады. Кітапхананы сонымен қатар, бірінші кезеңде кері транскриптазаны және мРНҚ қоспасын қолданған кезде де алуға болады. Кейде бұны, қажетті мРНҚ-ны таза күйінде бөліп алу мүмкін болмағанда, жасауға тура келеді. Осылайша, жалғыз емес ген (бір типтің мРНҚ-сымен синтезделген немесе есептелген) клондалған жағдайларда, төртінші кезеңнен кейін кітапханалар түріндегі бактериялық өсірінділерді алады.